
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Отряд Fusulinida (А. Д. Миклухо-Маклай, Д. М. Раузер-Черноусова, С. Е. Розовская)
Раковина спирально-плоскостная, инволютная, реже эволютная, двусторонне симметричная, от сжатой по оси навивания или шарообразной до веретенообразной или цилиндрической, иногда развертывающаяся в последнем обороте; по многочисленным оборотам раковина подразделена на большое число камер септами, иногда и септулами - выростами стенок. Ось навивания ранних оборотов нередко повернута к оси последующих. Стенка раковины секреционная известковая, микрозернистая, из одного или нескольких слоев, с порами в виде параллельных трубочек или сложных альвеол. Дополнительные скелетные образования в виде хомат, псевдохомат, парахомат, осевых уплотнений. Устья в основании септ, единичное срединное или в виде ряда отверстий; кроме того, могут быть дополнительные отверстия в септах (куникули и септальные поры) и в спиральных септулах. Карбон - пермь, Надсемейства: Fusulinidea и Verbeekinidea.
Первая фузулина была описана в 1829. г. Фишером в России. Основоположником изучения фузулинид следует считать В. И. Мёллера (1878).
Наиболее характерными признаками отр. Fusulinida можно считать особенности септального аппарата, т. е. складчатость септы и строение септул, структуру стенок раковины и характер дополнительных отложений.
Септы составляют со стенкой раковины обычно почти прямой угол (рис. 203); септальные борозды более или менее прямолинейны или слегка волнисты и септы, являясь непосредственным продолжением стенки, обычно сохраняют общую с ней структуру, хотя бы в своей верхней части; септы чаще плоские, реже выпуклые вперед по навиванию. Складчатость септ может отсутствовать (прямые септы), или может быть широкой и неглубокой (волнистые септы), или узкой и глубокой. В последнем случае она обнаруживается в осевом сечении в виде арок по основанию оборота, образованных при разрезе складок септ (рис. 204, 205). В случае правильной и глубокой складчатости происходит подразделение нижней части камеры на камерки путем срастания соседних складок, направленных друг к другу противоположными сторонами (рис. 204). Иногда имеются френотеки - тонкие пластинки, разделяющие у некоторых родов камеры поперек и прикрепляющиеся различным образом к септам, а также к основанию или потолку камер. У некоторых представителей надсем. Verbeekinidea, кроме септ, имеются еще септулы, образованные выростами стенки без ее загиба. Септулы могут располагаться между септами продольно от полюса к полюсу и в перпендикулярном направлении; первые называются аксиальными, вторые - спиральными. Последние бывают первого и второго порядка (рис. 206). Аксиальные септулы и спиральные первого порядка в своей нижней части нередко уплотнены, иногда булавовидно расширены с тонкой верхней частью. Спиральные септулы первого порядка длинные, соприкасаются с парахоматами (рис. 207).
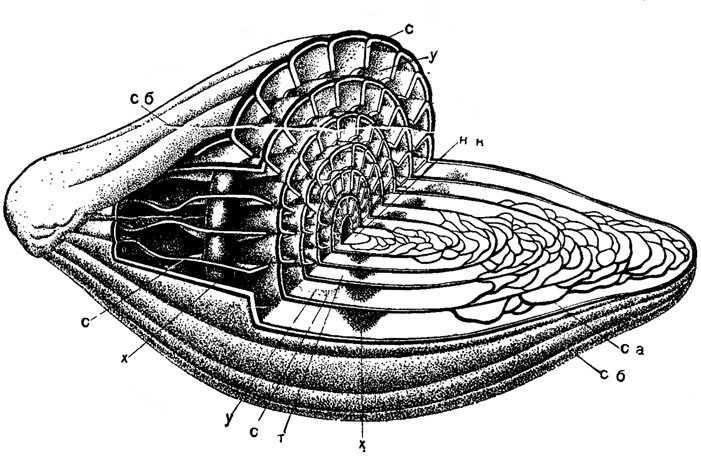
Рис. 203. Схема строения Fusulinella: нк - начальная камера, са - септальные арки, с - септы, сб - септальные борозды, х - хоматы, у - устье, т - туннель
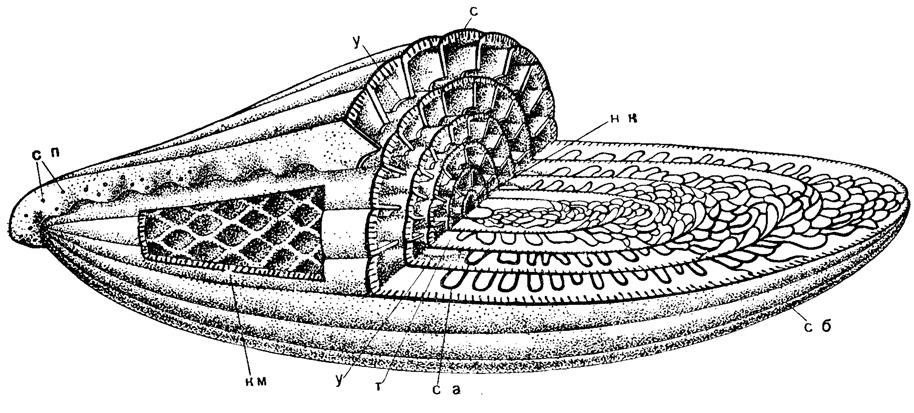
Рис. 204. Схема строения Pseudofusulina: нк - начальная камера, с - септы, сб - септальные борозды, са - септальные арки, км - камерки, у - устье, т - туннель, сп - септальные поры
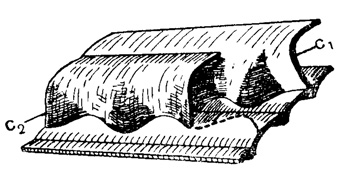
Рис. 205. Схема складчатых перегородок Fusulinidae: с1 - септа, с2 - последующая септа - частично удалена, а след ее соприкосновения со стенкой предыдущего оборота показан пунктиром (Sigal, 1952)
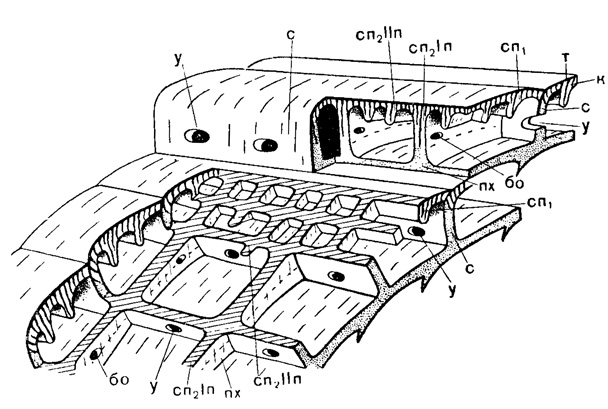
Рис. 206. Схема внутреннего строения Neoschwagerina: т - тектум, к - кернотека, с - септы, сп - септулы: сп1 - аксиальные, сп2 - спиральные; I п - 1 порядка, II п - II порядка, у - устье, бо - боковые отверстия, пх - парахоматы (Sigal, 1952)
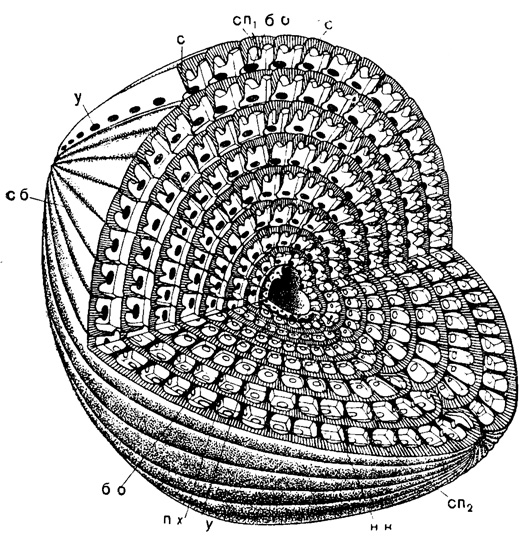
Рис. 207. Схема строения Neoschwagerina: нк - начальная камера, с - септы, сб - септальные борозды; септулы: сп1 - септулы аксиальные, сп2 - септулы спиральные, пх - парахоматы, у - устье, бо - боковые отверстия (по Дуткевичу, 1932)
Стенка раковины Fusulinida достигает очень большой сложности строения и служит признаком высокого таксономического значения (рис. 208). У наиболее примитивных форм (эоштаффелл) стенка однослойная, состоит из первичного слоя - протеки, которая оконтуривается сверху тонкой пленкой более темного цвета - тектумом. В процессе развития стенки выделяется вторичный слой - текторий (профузулинелловый тип). Уже на этой стадии развития в стенке нередко появляются тонкие прямые поры (Соловьева, 1955). Позднее стенка становится четырехслойной (фузулинелловый тип), состоящей из тектума, диафанотеки (более прозрачный слой) и двух текториев (наружного и внутреннего). Поры нередко пронизывают все слои. У некоторых родов (протритициты, гемифузулины, путреллы) дифференциация на слои в наружных оборотах становится менее заметной и поры более грубыми. Этот тип стенки является переходной ступенью от четырехслойной стенки фузулинеллового типа к двухслойной швагеринового, состоящей из тектума и кериотеки. Тонкие поры наблюдаются в стенке с кериотекой только в верхней части, а в нижней, путем слияния или разветвления пор, образуются более широкие полости (альвеолы), разделяющиеся трабекулами, утолщающимися книзу. У вербеекинид наблюдается пористая протека и иногда более плотный слой, у неошвагеринид строение сходное с кериотекальным, но трабекулы сливаются в нижней части и удлиняются, образуя септулы, а у наиболее специализированных представителей стенка становится очень тонкой и плотной.
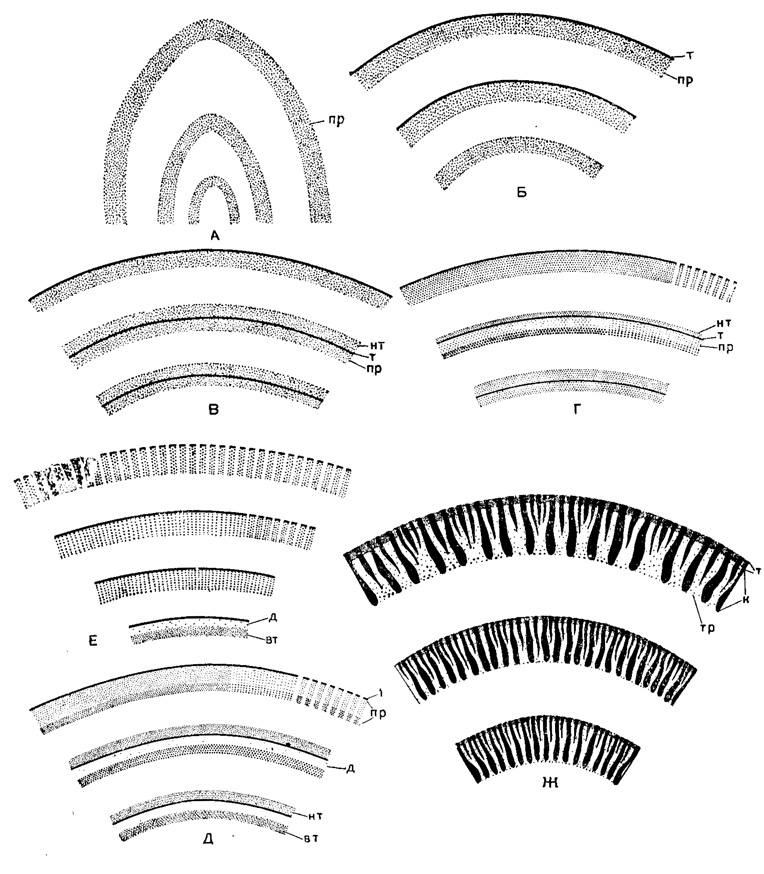
Рис. 208. Строение стенки Fusulinidea: А - Eostaffella, Б - Schubertella, В - Profusulinellaf Г - каширские Profusulinella и Аliutovella, Д - Fusulinella, Е - Hemifusulina, Ж - Schwagerina; пр - протека, т - тектум, нт - наружный текториум, вт - внутренний текториум, д - диафанотека, к - кериотека, тр - трабекулы
Дополнительные скелетные образования, отлагающиеся поверх стенки раковины Fusulinida разнообразны. В пределах надсем. Fusulinidea это хоматы, псевдохоматы и осевые уплотнения, а у надсем. Verbeekinidea - парахоматы. Хоматы представляют собой два спиральных валика, проходящих в срединной области раковины по основанию оборотов и ограничивающих с двух сторон устье последовательных септ. У Verbeekinidea таких валиков много вдоль многочисленных устьев (рис. 209).
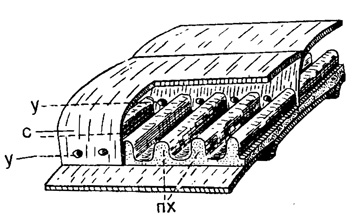
Рис. 209. Схема внутреннего строения Misellina: с - септа, пх - парахоматы, у - устье (Sigal, 1952)
Спиральный канал между устьями, открытый сверху и ограниченный с боков хоматами, парахоматами или складками септ, называется туннелем. Из надсем. Fusulinidea только у полидиексодин туннелей много. У сильно складчатых форм подсем. Polydiexodinae сообщение камер между собой осуществляется еще дополнительно через куникулы, т. е. каналы, проходящие через резорбированную нижнюю часть септ в местах соприкосновения вершин соседних складок, направленных друг к другу (рис. 210). У Verbeekinidea в спиральных септулах первого порядка наблюдаются овальные отверстия, так называемые боковые отверстия.
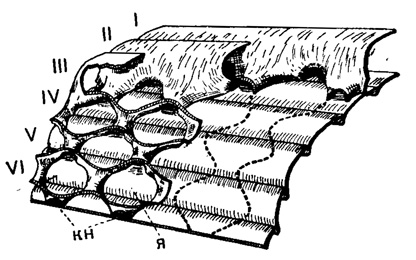
Рис. 210. Схема внутреннего строения Parafusulina: в левой части чертежа камеры III-VI показаны частично сломанными; видно как происходит слияние противоположных складок с образованием куникулей (кн) и ячеек (я); в правой части чертежа перегородки вовсе удалены, линии их соприкосновения с поверхностью предыдущего оборота показаны пунктиром (Sigal, 1952)
Сем. Fusulinidae, установленное Мёллером в 1878 г., объединяло три рода: Fusulina Fischer, Schwagerina Moeller и Hemifusulina Moeller. В конце XIX и начале XX в. систематика фузулинид изменилась незначительно. Бурное изучение их в тридцатых и сороковых годах привело к разукрупнению сем. Fusulinidae на ряд подсемейств. В этот период классификация пересматривалась многими авторами (С. Dunbar, 1933, см. Cushman, 1933; Galloway, 1933; Дуткевич, 1934; Gubler, 1935; Раузер-Черноусова, 1937;Dunbar and Skinner, 1931, 1937; Dunbar and Henbest, 1930; Путря, 1940 и др.). Однако все они ограничивались пересмотром системы в пределах подсемейств или родовых групп.
Данбар и Хенбест (Dunbar and Henbest, 1942) признали существование четырех подсемейств, Томпсон (Thompson, 1948) предложил классификацию, которая делит Fusulinidae на шесть подсемейств. В том же году Глесснер (Glaessner, 1948) предложил надсем. Endothyridae, охватывающее фузулинид и эндотирид, а Данбар (Dunbar, 1948, см. Cushman, 1948, рр. 142-170) сем. Fusulinidae разделил на два семейства: Fusulinidae и Neoschwagerinidae. Позднее А. Д. Миклухо-Маклай (1949,1953), выделив дополнительно два подсемейства, включил в Fusulinidae семь подсемейств; С. Е. Розовская (19501) ввела еще одно подсем. Fusulinellinae, установленное еще в 1910 г. X. Штаффом и Д. Ведекиндом (Staff und Wedekind, 1910). Сири (Ciry, 1952; Sigal, 1952) классифицирует фузулинид как надсемейство, приводя в нем четыре подсемейства.
Ревизия всех существующих классификаций фузулинид позволила нам прийти к выводу о недостаточности формально-морфологического метода, положенного в основу большинства ранее существующих схем систематики, и на основе онто-филогенетического метода предложить систему, приближающуюся к естественной, в следующем виде: отряд Fusulinida с двумя надсемействами: Fusulinidea и Verbeekinidea. Первое объединяет четыре семейства (Ozawainellidae, Fusulinidae, Schubertellidae, Schwagerinidae) с девятью подсемействами, второе - два семейства (Verbeekinidae и Neoschwagerinidae).
Fusulinida появляются с ранневизейского времени. Лишь с начала ср. карбона начинается более быстрое развитие сем. Ozawainellidae с отделением от него сем. Fusulinidae и Schubertellidae. Следующий этап развития Fusulinida можно начинать с верхнекаменноугольного времени, когда появляются представители сем. Schwagerinidae, достигающего расцвета в швагериновое время. В пермское время в области Тетиса происходит усиленное родообразование во всех ветвях филогенетического древа и появляется новое надсем. Verbeekinidea.
Географическое распространение Fusulinida отличается большой широтой, особенно в нижнем и среднем карбоне. Только с конца верхнего карбона наблюдается некоторое обособление родового и видового состава бассейнов разных стран. В перми в области Тетиса развит весьма своеобразный высокоспециализированный тип фузулинид, которые целиком вымирают в конце перми.
Фузулиниды, по всей вероятности, произошли на заре визейского века от эндотир группы Endothyra staffelloides, отличающихся относительной симметричностью навивания, сравнительно большим числом оборотов и камер и присутствием хомат. В визейское время обособились два крупных ствола: Pseudoendothyra с толстой диафанотекой и Eostaffella со слабо дифференцированной стенкой (рис. 211). Псевдоэндотиры только в пермское время дают начало новым родам подсем. Staffellinае (Staffella, Narikinella, Sphaerulina и др.) с видами крупных размеров и с большим числом оборотов. По-видимому среди последней группы родов находятся корни Verbeekinidea: наиболее примитивные Eoverbeekina очень сходны со Sphaerulina.
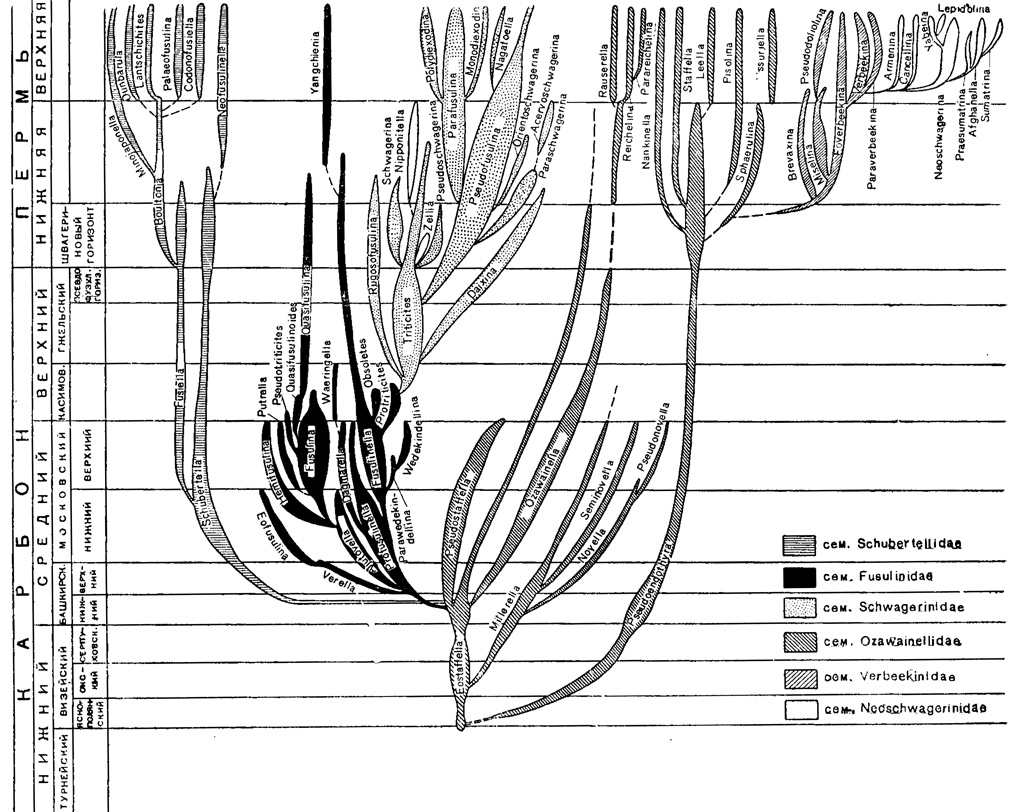
Рис. 211. Филогенетическая схема отряда Fusulinida
От Eostaffella, исходного рода подсем. Ozawainellinae, в середине визейского века берут начало Milleretla, а с башкирского века - Pseudostaffella и Ozawainella. От озаваинелл в позднепермское время произошли развернутые Reichelina и Parareichelina и неправильно навитые Rauserella.
От эоштаффелл в раннебашкирское время отделяется сем. Schubertellidae, характеризующееся также медленной эволюцией и слабой дифференциацией стенки. Фузиеллы дают начало роду Boultonia подсем. Boultoniinae со своеобразной складчатостью вследствие особой тонкости септ, присущей всей ветви этого интересного подсемейства. Характерно появление в конечных ветвях этого древнего семейства ряда родов с развернутой раковиной.
Уже в раннебашкирское время от псевдоштаффелл отделяются Profusulinella (подсем. Fusulinellinae), дающие в позднебашкирское время начало двум родам: Verella (подсем. Eofusulininae) и Aljutovella. У последней уже появляется складчатость септ, дальнейшее развитие которой приводит к выделению подсем. Fusulininae. Тесно соединен с фузулинеллами (по характеру онтогенеза и отсутствию резкого морфологического перерыва) род Protriticites, появляющийся с середины мячковского времени и в касимовское время дающий переходы к роду Тriticites сем. Schwagerinidae. Для подсем. Schwagerininae в начале позднего карбона исходным является Род Triticites, повсеместно распространенный и вымирающий в ранней перми. В швагериновом горизонте и перми господствующее положение занимает под Pseudofusulina. Доказанным можно считать происхождение от тритицитов ругозофузулин с морщинистой стенкой; несколько позднее, с псевдофузулинового времени, путем редукции хомат даиксин, образуются Pseudofusulina с правильной складчатостью и в швагериновое время - шарообразные формы Pseudoschwagerina, Zellia и Schwagerina. Эти три рода наиболее полно развиты в швагериновое время, но лишь псевдошвагерины распространены не только в Евразии, но и в Америке. От псевдофузулин в швагериновое время отделяются известные повсюду вздутые Paraschwagerina, с тесно свернутыми оборотами на юношеской стадии развития.
В пермское время отделяется другая весьма важная ветвь длинных цилиндрических форм с куникулами - Parafusulina (подсем. Polydiexodininae), распространенных почти повсеместно. По-видимому, потомками этого рода являются позднепермские роды: восточноазиатский Моnodiexodina, с очень низкой сладчатостью септ, и Polydiexodina, с многочисленными туннелями.
В пределах надсем. Verbeekinidea четко выделяются два семейства: Verbeekinidae и Neoschwagerinidae. От Eoverbeekina намечаются три филогенетические ветви: путем резких изменений формы раковины в направлении сжатия по оси навивания образуется Brevaxina, и вытягивания по оси - Misellina; появление у последней альвеолоподобной структуры стенки указывает на направление эволюции к конечному члену второй ветви - Pseudodoliotina. К третьей ветви относятся Verbeekina и Paraverbeekina. От последней, путем появления спиральных септул 1-го порядка в наружных оборотах, происходит А темпа, дающая постепенные переходы к Саnсеllinа, с хорошо развитыми спиральными септулами 1-го порядка, и затем к Neoschwagerina. Последние путем более сильного развития аксиальных и спиральных септул 2-го порядка переходят в Yabeina и затем Lepidolina, с тонкой и почти однородной стенкой. Вторая ветвь, представленная родами Praesumatrina, Afghanella и Sumatrina, характеризуется специализацией септул, изменением структуры стенки и ее утонением.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'