
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Надсемейство Nonionidea Schultze, 1854
[nom. transl. Subbotina, 1959 (ex Nonionidae Schultze, 1854)]
Начальная часть раковины трохоидная, поздняя - различно устроенная. Стенка пористая зернистая. Много прикрепленных форм. Семь семейств: Anomalinidae, Nonionidae, Planorbulinidae, Rupertiidae, Victoriellidae, Homotremidae, Cymbaloporidae.
Раковина прикрепленная или свободная, на ранней стадии спирально-коническая, на поздней - скрыто-трохоидная, уплощенно-выпуклая или равнодвояковыпуклая, иногда с двухрядным, однорядным или полукольцевым нарастанием камер последних оборотов (рис. 558). Устье в виде щели или арки в основании септальной поверхности с продолжением на одну из сторон (боковая часть устья) или на септальную поверхность; иногда протягивается до пупка, редко в септальные швы, у прикрепленных очень различно по форме и положению (рис. 559). Стенка однослойная и многослойная (рис. 560) с хитиноидным прослоем у камер начальных оборотов, равномерно или неравномерно пористая, часто со скульптурой (рис. 561). Юра - ныне. Два подсемейства: Anomalininae и Cibicidinae.
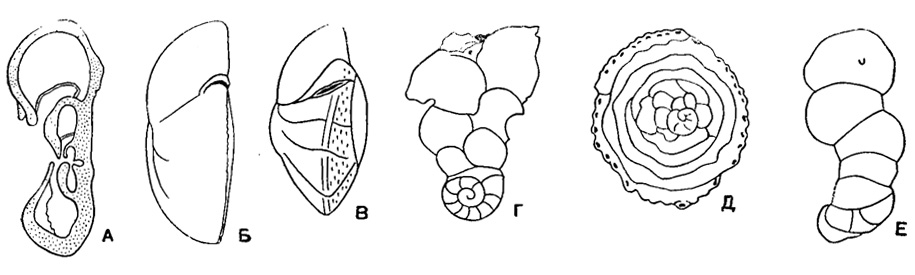
Рис. 558. Характер навивания спирали и форма раковины у представителей сем. Anomalinidae. A - спирально-коническое строение на ранней стадии и скрыто-трохоидное во взрослой стадии; Б, В - свободно живущие формы: Б - уплощенно-выпуклая раковина, В - равнодвояковыпуклая раковина; Г, Е - прикрепленные формы: Г - раковина с последними камерами, расположенными в два ряда, Д - раковина с полукольцевыми последними камерами дисковидная, Б - раковина с неправильно однорядным расположением последних камер
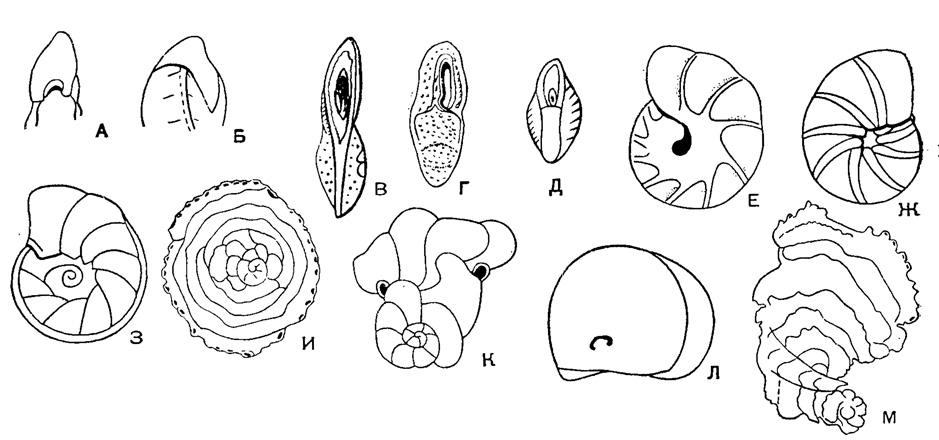
Рис. 559. Различная форма и положение устья у представителей сем. Anomalinidae. А-Б - устье на периферическом крае: А - арковидное, Б - щелевидное; В-Д - устье удлиненное, на септальной поверхности; Е-З - устье щелевидное: Е - на брюшной стороне доходит до пупка, Ж - заходит в септальные швы, З - протягивается наспинную сторону; И - устья в виде крупных пор на периферии; К - устья округлые с короткой шейкой и губой; Л - устье терминальное с зубом; М - устья в виде сосочков на периферии последней камеры
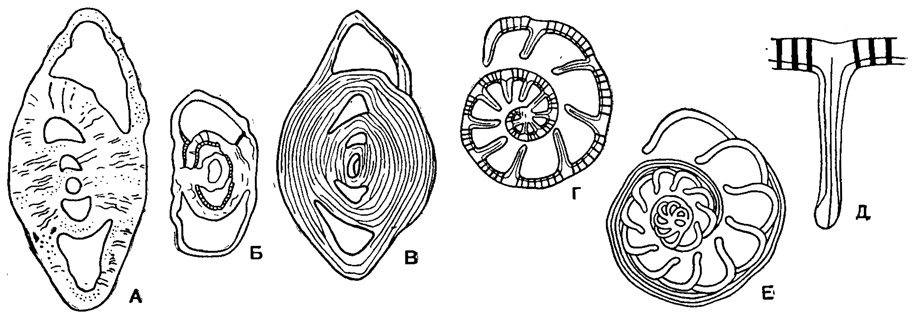
Рис. 560. Строение стенки у представителей сем. Anomalinidae. А-В, осевое сечение (по Б. М. Келлеру): А - однослойная, Б - двуслойная, В - многослойная; Г-Е, поперечное сечение (по Andreae): Г - трехслойная стенка у спиральной и септальных перегородок, Д - то же в увеличенном виде, Е - однослойные септальные перегородки и постепенное утолщение спиральной перегородки
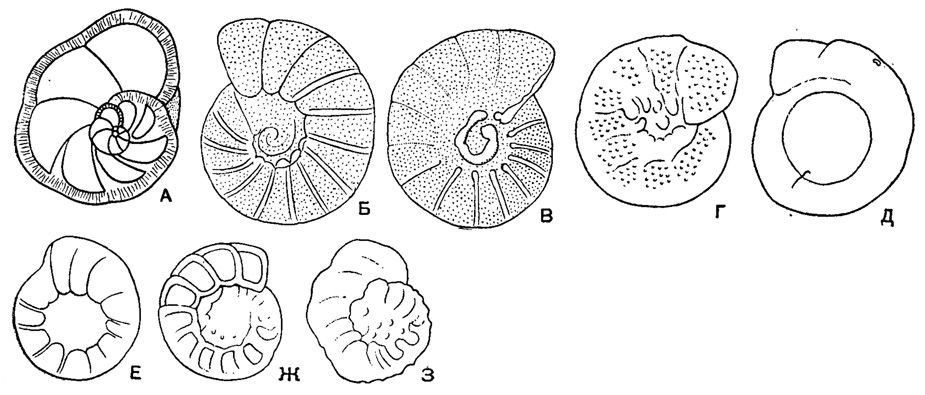
Рис. 561. Пористость и скульптура стенки у представителей сем. Anomalinidae. А - поровые каналы, пронизывающие стенку (схема сечения); Б, В - равномерная пористость у рода Anomalina (Brotzenella); Г, Д - неравномерная пористость у рода Anomalina (Pseudovalvulineria); Б - прозрачная шишка на спинной стороне у А. (Pseudovalvulineria) и стекловатые швы (Б, В) на обеих сторонах - раковины; Д, Е - непрозрачные шишки на спинной стороне А. (Pseudovalvulineria) и на брюшной стороне у Cibicides (Cibicidoides); Ж, З - скульптированные септальные швы на спинной стороне у С. (Cibicidoides) и А. (Pseudovalvulineria); Ж - двуконтурные уплощенные, выпуклые швы на спинной стороне у С. (Cibicidoides); З - бугристые, выпуклые и шиповидные швы на спинной стороне Anomalina (Pseudovalvulineria)
Спирально-коническое строение начальной части раковины и хитиноидный прослой в начальных оборотах доказывает генетическую связь аномалинид с роталиидами. Возможно, они возникли из роталоидного силурийского рода Aristerospira Ehrenberg, 1858. В нижнем и особенно в в. мелу обильно развиты роды Cibicides и Anomalina (подроды Gavelinella, Pseudovalvulineria, CibicidoideSy Anomalinoides, Gemellides), появляются Stichocibicides и Karreria. Наибольшего расцвета все они достигли в кайнозое, где еще появились роды Ganella и Palmerinella и особенно широко развились представители рода Cibicides. Основная линия развития прослеживается на свободных и полуподвижных формах, но наибольшего разнообразия достигают прикрепленные, сильно специализированные. Среди них выделяются формы с уплощенно-кольцевыми камерами - Cyclocibicides, Annulocibicides, с двурядно выпрямленными рядами камер - Dyocibicides, с однорядно нарастающими камерами - Stichocibicides и с сильно уплощенной эволютной раковиной - подроды Laticarinina, Planulina.
Раковина свободная, уплощенно-выпуклая, реже вдавленная; периферический край округлый или слабо заостренный (рис. 562). Боковая часть устья на брюшной стороне, реже устье только на септальной поверхности камеры, очень редко продолжается в септальные швы (рис. 559). Стенка равномерно пористая, реже более крупные поры на брюшной стороне (рис. 561). Четыре рода.
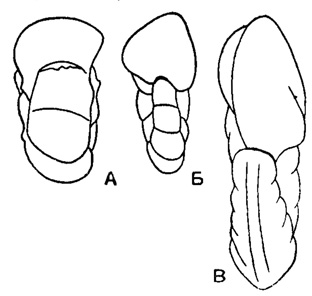
Рис. 562. Характер периферического края у родов подсемейства Anomalininae. А - широко-округлый; Б - закругленный; расширяющийся к концу оборота; В - узкий, заостряющийся
Anomalina Orbigny, 1826. Тип рода - А. punctulata Orbigny, 1826; современный, Франция. Раковина уплощенно-выпуклая; спинная сторона более эволютная, чем брюшная, с шишкой в центральной части; на брюшной стороне отчетливый пупок, узкий или очень широкий, часто закрыт пластинками или пупочной шишкой (рис. 563). Периферический край закругленный, широкий. Устье заходит в пупок, реже не доходит до него, иногда заходит в септальные швы. Стенка чаще равномерно пористая (рис. 561). Виды многочисленные. Юра - ныне. Широко распространенный род. Четыре подрода: Anomalina Orbigny, 1826 (рис. 564-565); Gavelinella Brotzen, 1942 (рис. 566); Pseudovalvulineria Brotzen, 1942 (рис. 567); Brotzenella Vassilenko, 1958 (рис. 568).
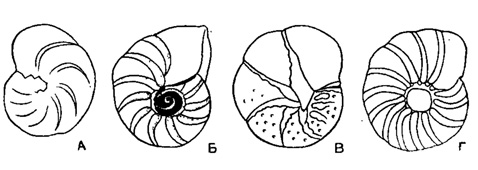
Рис. 563. Характер пупочной области для раковин рода Anomalina. А - пупок узкий, открытый (подрод Anomalina); Б - пупок широкий, окружен тонкой каймой (подрод Gavetinetta); В - пупок узкий, закрыт широкими пластинками, заходящими в септальные швы (подрод Pseudovalvulineria); Г - пупок закрыт обособленной стекловатой шишкой (подрод Bro'zenella)

Рис. 564-572. Сем. Anomalinidae. 564 А-В. Anomalina (Anomalina) affinis (Hantken); внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 50, в. эоцен, п-ов Мангышлак (колл. Н. К. Быковой). 565 А-В. Anomalina (Anomalina) granosa (Hantken); внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 50, в. эоцен, п-ов Мангышлак (колл. В. П. Василенко). 566 А-В. Anomalina (Gavelinella) pertusa (Marsson); внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 50, в. Маастрихт, Украина, р. Десна (колл. В. П. Василенко). 567 А-В. Anomalina (Pseudovalvulineria) clementiana (Orbigny); внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 50, в. сантон, Прикаспийская впадина, пос. Джаксыбай (Дайн, 1934). 568 А-В. Anomalina (Brotzenella) monterelensis Marie; внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 50, в. кампан, Гурьевская обл., г. Актулагай (В. П. Василенко, 1954). 569-570. Ganella neumannae Aurouse et Boulanger. 569 А-Б, взрослый экземпляр, внешний вид: А - сбоку, Б - с периферического края; 570 А, Б - молодой экземпляр, внешний вид: А - сбоку, Б - с периферического края, × 53, ипрский ярус, Гана, Нижние Пиренеи (Aurouse et Boulanger, 1954). 571 А-В. Boldia lobata (Terquem); внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 40, эоцен, Франция (Terquem, 1882). 572 А-В. Palmerinella palmerae Bermudez; внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 50, современный, побережье о-ва Кубы (Bermudez, 1934)
Ganella Aurouze et Boulanger, 1954. Тип рода - G. neumannae Aurouze et Boulanger, 1954; ипрский ярус, Гана (Н. Пиренеи). Раковина спирально-плоскостная в начальной стадии, полуэволютная с обеих сторон; во взрослой стадии слабо трохоидная. Устье в виде удлиненной щели, окаймленное губой, расположено на уплощенной септальной поверхности; только у молодых особей оно доходит до ее основания (рис. 569-570). По характеру строения начальной части род включен в это семейство только условно. Один вид в ипрском ярусе Н. Пиренеев.
Вне СССР; Boldia van Bellen, 1946 (рис. 571), Palmerinella Bermudez, 1934 (рис. 572).
Раковина прикрепленная или свободная, асимметричная, выпуклая или уплощенная. На поздней стадии у специализированных родов с двухрядным, однорядным или полукольцевым расположением камер. У свободных форм контур округлый, у прикрепленных - различных очертаний. Боковая часть устья на спинной стороне; у прикрепленных устье терминальное, в виде крупных пор или трубочек, в виде круглых или овальных отверстий, с зубом или с губой. Обычно стенка крупнопористая на спинной стороне, редко равномерно пористая, иногда многослойная. Девять родов (Рис. 558 и 559).
Cibicides Montfort, 1808 (Buningia Finlay, 1939). Тип рода - Cibicides refulgetis Montfort, 1808; современный, Адриатическое море. Раковина от плоской до сильно выпуклой; спинная сторона плоская, вдавленная или выпуклая, более эволютная, чем брюшная; пупок узкий, иногда с шишкой или различается начальный оборот (рис. 573). Периферический край узкий, широкий, килеватый, уплощенный, реже округлый (рис. 574). Боковая часть устья на спинной, реже и на брюшной стороне. Стенка обычно неравнопористая. Виды многочисленные. Карбон?, триас?, юра?, мел - ныне. Широко распространенный род. Шесть подродов: Cibicides Montfort, 1808 (рис. 575); Cibicidoides Brotzen, 1942 (рис. 576); Anomalinoides Brotzen, 1942 (рис. 577); Gemellides Vassilenko, 1954 (рис. 578); Planulina Orbigny, 1826 (рис. 579); Laticarinina Galloway et Wissler, 1927 (рис. 580-581).
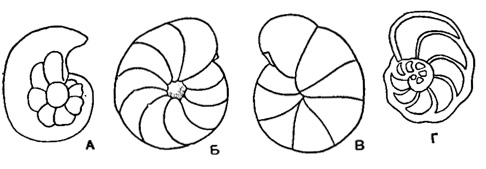
Рис. 573. Характер пупочной области у рода Cibicides. А, Б - пупок отчетливый, закрыт стекловатой шишкой, камеры сходятся к ней плотным кольцом (подроды Laticarinina, Cibicidoides); В - пупок закрытый, камеры сходятся плотно, шишки нет (подрод Anomalinoides); Г - в пупочной области различается начальный оборот (подрод Planulina)
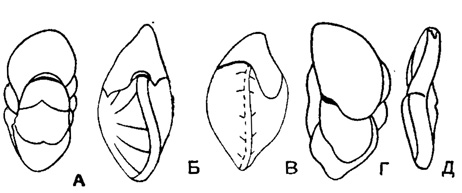
Рис. 574. Периферический край и форма септальной поверхности у рода Cibicides. А - край широкий, округлый, септальная поверхность вздутая, округлая (подрод Anomalinoides); Б, В - край широкий, заостренный, септальная поверхность треугольная, стрельчатая (подроды Cibicidoides и Gemellides); Г - край уплощенно-выпуклый, узкий, септальная поверхность овально-треугольная (подрод Cibicides); D - край узкий, уплощенный, септальная поверхность узкая, плоская (подрод Planulina)

Рис. 575-584. Сем. Anomalinidae. 575 А-В. Cibicides (Cibicides) refulgens Montfort; внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 50, современный, Баренцево море (колл. Н. А. Волошиновой). 576 А-В. Cibicides (Cibicidoides) eriksdalensis Brotzen; внешний вид: A - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 50, н. сантон, Украина, р. Десна (В. П. Василенко, 1954). 577 А-В. Cibicides (Anomatinoides) pinguis (Jennings) subap. neckajae Vassilenko; внешний вид: A - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 50, в. сенон, З. Сибирь, ст. Называевская (В. П. Василенко, 1954). 578 А-В. Cibicides (Gemellides) orcinus Vassilenko; внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 50, н. Маастрихт, п-ов Мангышлак (В. П. Василенко, 1954). 579 А-В. Cibicides (Planulina) arimiriensis (Orbigny); внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 50, плиоцен, Албания (колл. В. П. Василенко). 580-581. Cibicides (Laticarinina) pauperaia (Parker et Jones); 580 - более эволютный экземпляр; внешний вид сбоку; 581 А-Б, типичный экземпляр; внешний вид: А - сбоку, Б - с периферического края, × 30, современный, сев. часть Атлантического океана (Parker et Jones, 1865). 582-583. Stichocibicides cubensis Cushman et Bermudez; 582 A-Б, внешний вид: A - со свободной (брюшной) стороны, Б - со стороны устья, × 25, 583 А-Б, внешний вид: А - со стороны прикрепления (спинной), Б - со стороны устья, × 33, эоцен, о-в Куба (Cushman and Bermudez, 1936). 584 А-В. Karreria fallax Rzehak; внешний вид: А - со стороны, прилегающей к субстрату (спинной), В - со свободной (брюшной) стороны, В - с устьевого окончания, × 50, датский ярус, п-ов Мангышлак (В. П. Василенко, 1954)
Stickocibicides Cushmanet Bermudez, 1936. Тип рода - S. cubensis Cushman et Bermudez, 1936; в. эоцен, Куба. Раковина прикрепленная, в ранней стадии как цибицидес, в поздней - камеры нарастают однорядно; спинная сторона уплощенная, эволютная, брюшная - выпуклая, инволютная; периферический край узкий, килеватый. Устье округлое или эллиптическое, терминальное, вблизи периферического края, без губы. Стенка мелкопористая, гладкая (рис. 582- 583). Единичные виды. Альб США; в эоцене Виргинии, о-вов Кубы и Гаити.
Karreria Rzеhak, 1891. Тип рода - К. fallax Rzehak, 1895; палеоген, Австрия. Отличается от предыдущего рода широким и округлым периферическим краем, более широкими и длинными камерами, устьем с зубом у камер однорядной части (рис. 584). Несколько видов. Сенон - палеоцен СССР, З. Европы, Мексики.
Dyocibicides Cushman et Valentine, 1930. Тип рода - D. biserialis Cushman et Valentine, 1930; современный, у берегов Калифорнии. Раковина прикрепленная, спинная сторона уплощенная, в ранней части как у Cibicides, в поздней стадии камеры нарастают двухрядно, быстро увеличиваясь в размерах. Устье в виде удлиненной щели на внешнем крае последней камеры с губой. Стенка крупнопористая (рис. 585). Несколько видов. Эоцен Колумбии; миоцен Ю. Германии.

Рис. 585-594. Сем. Anomalinidae. 585 А-Б. Dyocibicides biserialis Cushman et Valentine; внешний вид; A - со стороны прикрепления (спинной), Б - со свободной (брюшной) стороны, × 30, современный, Калифорния (Cushman et Valentine, 1930). 586-587. Vagocibicides maoria Finlay; внешний вид: 586 - взрослый экземпляр со свободной (брюшной) стороны, 587 - молодой экземпляр с периферического края со стороны устья, × 20, третичный, Новая Зеландия (Finlay, 1939). 588-589. Rectociblcides miocenicus Cushman et Ponton; внешний вид: 588 - типичный экземпляр, 589 - экземпляр с трубковидными устьями, × 16, н. миоцен, Флорида (Cushman and Ponton, 1932). 590-592 Cibicidella variabilis (Orbigny); 590 - внешний вид, современный, Канарские о-ва (Orbigny, 1839); 591 А-Б, внешний вид: А - со стороны прикрепления (спинной), Б - со свободной (брюшной) стороны, современный (Soldani, 1789); 592 - внешний вид, современный (Cushman, 1927); 593 А-Б. Annulocibicides projectens Cushman et Ponton; внешний вид: A - со стороны прикрепления (спинной), в - со свободной (брюшной) стороны, × 25, н. миоцен, Флорида (Cushman and Ponton, 1932). 594 А-Б. Cyclocibicides vermiculatus (Orbigny); внешний вид: A - со стороны прикрепления (спинной), Б - со свободной (брюшной) стороны, × 30, современный, Средиземное море (Brady, 1884)
Vagocibicides Finlay, 1939. Тип рода - V. maoria Finlay, 1939; третичные, Новая Зеландия. В начальной части раковина как у Cibicides, в дальнейшем несколько камер расположено двухрядно, а затем однорядно. Устье у камер двухрядной стадии в центре устьевой поверхности, со слабо приподнятым краем, позднее округлое, в несколько пониженной части устьевой поверхности. Стенка мелко и равномерно пористая (рис. 586-587). Единичные виды в третичных отложениях Новой Зеландии и в плиоцене Японии.
Вне СССР: Rectocibicides Cushman et Ponton, 1932 (рис. 588-589); Cibicidella Cushman, 1927 (рис. 590-592); Annulocibicides Cushman et Ponton, 1932 (рис. 593); Cyclocibicides Cushman, 1927 (рис. 594).
Роды Anomalinella Cushman, 1927 и Webbina Orbigny, 1839 в семейство не включены, так как их систематическое положение и генетические взаимоотношения с другими родами семейства пока не ясны.
[nom. transl. Reuss, 1860 (ex Nonioninae Schultze, 1854)]
Раковина свободная, большей частью на поздних стадиях развития спирально-плоскостная, двусторонне-симметричная, редко асимметричная. Состоит из небольшого числа оборотов (11/2-3). Устье большей частью медианное, или щелевидное, или в виде одного ряда округлых отверстий, редко ситовидное или септальное. У некоторых родов имеется система каналов; в последнем случае в пупочной области развита скульптура из прозрачного скелетного вещества в виде шишки или зернистости, а иногда имеются и выходные отверстия каналов в пупочной области и на септальных швах. Стенка обычно стекловатая - зернистая. Н. мел - ныне. Три подсемейства: Nonioninae, Nonionellinae и Меlonisinae. Тесно связано с сем. Anomalinidae, от представителей которого происходят роды с инволютной раковиной.
Раковина спирально-плоскостная, двусторонне-симметричная на всех стадиях развития, большей частью инволютная. Камеры увеличиваются равномерно, образуя низкую, плотно навитую спираль. Устье медианное щелевидное, или в виде округлых отверстий, расположенных в один ряд или в 2-3 параллельных ряда (ситовидное). Шесть родов.
Planomalina Loeblich et Tappan, 1946. Тип рода - Р. apsidostroba Loeblich et Tappan, 1946; н. мел, Техас. Раковина более или менее эволютная. Устье щелевидное. Стенка грубопористая, без скульптуры (рис. 595-596). Один вид в н. мелу США. Возможный предок инволютных нонионин.

Рис. 595-615. Сем. Nonionidae (подсем. Nonioninae, Nonionellinae). 595-596. Planomalina apsidostroba Loeblich et Tappan; 595 - внешний вид сбоку, 596 А-Б, внешний вид: А - сбоку, Б - с периферического края, × 46, н. мел, Техас (Loeblich and Tappan, 1946). 597 А-Б. Pullenia sextacamerata Pischvanova; внешний вид: А - сбоку, Б - с периферического края, × 50, миоцен, зап. обл. Украины (колл. Е. Д. Скворцовой). 598-599. Nonion incrassatus (Fichtel et Moll); 598 А-Б, взрослая особь, 599 A-Б, молодая особь, внешний вид: А - сбоку, Б - с периферического края, × 50, плиоцен, Албания (колл. Е. А. Храмой). 600. Nonion laevis (Terquem); аксиальное сечение: ск - спиральные каналы, пк - пупочные каналы, ф - форамен, × 50. эоцен, окр. Парижа (колл. Н. А. Волошиновой). 601-602. Porosononion subgranosus (Egger); 601 А-Б, внешний вид: А - сбоку, Б - с периферического края; 602 - аксиальное сечение: ск - спиральные каналы, пк - пупочные каналы, ф - форамен, × 50, сармат, С. Кавказ (колл. А. К. Богдановича). 603 А-Б. Cribrononion heteropora (Egger); внешний вид: А - сбоку, Б - с периферического края, × 50, четвертичный, Прикаспийская впадина (колл. Н. А. Волошиновой). 604-605. Astrononion gallowayi Loeblich et Tappan; 604 - аксиальное сечение, вскрыты камеры наружного оборота, × 70; 605 А-Б, внешний вид: А - сбоку, Б - с периферического края, × 50, современный, Баренцево море (колл. Н. А. Волошиновой). 606 А-В. Nonionella mlocenica Cushman; внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 50, миоцен, Сахалин (Волошинова, 1952). 607-608. Nonionellina labradorica (Dawson); 607 А-В, молодая трохоидная особь, внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края; 608 А-Б, взрослая инволютная особь, внешний вид: А - сбоку, Б - с периферического края, × 50, современный, Охотское море (колл Х. М. Саидовой). 609-610. Florllus boueanus (Orbigny); 609 А-Б, внешний вид: А - сбоку, Б - с периферического края, × 50; 610 - обломанный экземпляр, видно внутреннее строение, × 75, миоцен (тарханский горизонт), С. Кавказ (колл. А. К. Богдановича). 611 А-В. Florilus boueanus (Orbigny) var. janiformis Jones; внешний вид: A - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 50, миоцен (тортон), зап. обл. Украины (колл. А. К. Богдановича). 612 А-Б. Melonis melo (Orbigny); внешний вид: А - сбоку, Б - с периферического края, × 50, миоцен, зап. обл. Украины (колл. А. К. Богдановича). 613-614. Melonis soldanti (Orbigny); 613 А-В, начальная трохоидная стадия, внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края; 614 - аксиальное сечение, × 50, миоцен, Венский бассейн (колл. Н. А. Волошиновой). 615 А-Б. Paranonion venezuelanum Logue et Haas; внешний вид: A - сбоку, Б - с периферического края, × 50, в. миоцен, Венесуэла (Logue and Haas, 1943)
Pullenia Parker et Jones, 1862. Тип рода - Nonionina sphaeroides Orbigny, 1826 (Nonionina bulloides Orbigny, 1846); миоцен, Венский бассейн. Раковина шаровидная или сдавленная с боковых сторон; в обороте 4-8 камер, доходящих до центра пупочной области. Скульптура отсутствует. Устье щелевидное, длинное. Форамен более короткий, так как его краевые части зарастают (рис. 597). Более 30 видов. В. мел - ныне. Довольно широко распространенный род, но в небольших количествах экземпляров.
Nonion Montfort, 1808. Тип рода - Nautilus incrassatus Fichtel et Moll, 1798; современный, Средиземное море. Раковина сжатая с боковых сторон. В последнем обороте - 6-15 камер. Пупочная область большей частью выпуклая, со скульптурой. Имеются спиральный и пупочные каналы. Устье щелевидное, короткое, или в виде одного ряда отверстий в основании септ (рис. 598-600). 10-12 видов. В. мел - ныне. Широко распространен с неогена.
Porosononion Putrja, 1958. Тип рода - Nonionina subgranosa Egger, 1857; миоцен, Австрия. Подобен роду Nonion, но отличается ситовидным устьем и радиально-лучистой структурой стенки (рис. 601-602). Несколько видов. Миоцен (тарханский горизонт) - ныне. Европейская часть СССР и З. Европа.
Cribrononion Thalmann, 1946 (Canalifera Krascheninnikoff, 1953). Тип рода - Nonionina heteropora (Egger), 1857; миоцен, Бавария. Подобен роду Nonion, но отличается наличием отверстий каналов на септальных швах. От рода Elphidium Montfort, 1808 из сем. Elphidiidae отличается отсутствием ретральных отростков и зернистой микроструктурой стенки (рис. 603). Более 10 видов. Палеоцен Донбасса (?), в. эоцен - ныне СССР и З. Европа; н. эоцен Америки (?). Astrononion Cushman et Edwards,
1937. Тип рода - Nonionina stelligera Orbigny, 1839; современный, у берегов Канарских о-вов. Раковина с добавочными мелкими камерками в пупочной области, чередующимися с более крупными основными. Устье основных камер щелевидное (медианное). Дополнительные камеры открываются наружу посредством округлого или щелевидного отверстия, расположенного на периферическом крае (рис. 604-605). Около 10 видов. В. мел (?), эоцен - ныне. В ископаемом состоянии распространен широко, но в виде очень редких экземпляров. Принадлежность к сем. Nonionidae сомнительна.
Раковина, по меньшей мере на ранних стадиях, асимметричная, трохоидная, реже полностью симметричная, инволютная, с быстрым возрастанием в высоту оборотов спирали. Устье щелевидное, медианное, у трохоидных иногда протягивается на брюшную сторону. Три рода.
Nonionella Cushman, 1926 (Pseudononion Asano, 1936). Тип рода - Nonionella miocenica Cushman, 1926; миоцен, Калифорния. Раковина асимметричная, с брюшной стороны инволютная, со спинной - полуинволютная или эволютная. В некоторых случаях набрюшной стороне пупочный конец последней камеры прикрывает в виде лопасти пупочную область (рис. 606). Более 30 видов. В. мел - ныне. Широко распространен в Тихоокеанской области. В СССР (около 10 видов) встречается редко. В. мел Кавказа, Ср. Азии, Прикаспия; палеоген Донбасса, Ср. Азии, Кавказа; миоцен Сахалина.
Nonionellina Voloshinova gen. n. Тип рода - Nonionina labradorica Dawson, 1860; современный, залив св. Лаврентия. Раковина на ранних стадиях трохоидная, на поздних становится вполне инволютной; пупочная область углубленная, без скульптуры (рис. 607-608). Один вид. Миоцен Сахалина, Калифорнии. Современные.
Florilus Montfort, 1808. Тип рода - Nautilus asterizans Fichtel et Moll, 1798; современный, Средиземное море. Раковина, за исключением редких трохоидных форм, инволютная на всех стадиях развития. Пупочная область со скульптурой; развита система каналов (рис. 609-611). Около 10 видов. Миоцен Европейской части СССР и З. Европы; эоцен (?) Америки. Современные.
Раковина на ранних стадиях трохоидная, асимметричная, позднее двусторонне-симметричная, инволютная или слегка эволютная; характеризуется навиванием оборотов в низкую спираль с углубленной пупочной областью, обычно без скульптуры. Два рода.
Melortis Montfort, 1808. Тип рода - Nautilus pompilioides Fichtel et Moll, 1798; современный, Средиземное море. Раковина на поздних стадиях развития инволютная; пупок большей частью глубокий. Устье щелевидное, длинное, протягивается на боковые стороны. Форамен часто короче, так как у некоторых видов зарастает с боков (рис. 612-614). Примерно 20 видов. Эоцен - ныне. Широко распространенный род.
Paranonion Logue et Haas, 1943. Тип рода - Р. venezuelanum Logue et Haas, 1943; в. миоцен, Венецуэла. Раковина напоминает Melonis, но не вполне инволютная, двусторонне-симметричная. Устье у молодых особей в виде щели, протягивается от основания устьевой поверхности, у взрослых - овальное отверстие в средней части устьевой поверхности (рис. 615). Один-два вида. Миоцен Подолии (?), Венецуэлы (Ю. Америка).
Вероятно, к сем. Nonionidae относится Cribropullenia Thalmann, 1937, в СССР неизвестная.
Раковина прикрепленная спинной стороной в начальной стадии, иногда свободная у взрослых особой; начальная часть спирально-плоскостная или слабо трохоидная, в дальнейшем камеры нарастают кольцевидными рядами и радиально, чередуются в смежных рядах, вытягиваются и образуют неправильную сетку; нагромождаясь на предыдущие, образуют неправильно-шаровидную или конусовидную массу. Устья отчетливые, с губами с двух сторон каждой камеры на периферии дисковидных раковин или в виде крупных пор на поверхности конусовидных и шаровидных, в виде маленьких сосочков по краям удлиненных камер в сетчатых оборотах. Стенка обычно тонкая и равномерно крупнопористая; у ранних камер тонкий хитиноидный прослой. Иногда над центральной частью развит стекловатый натек (рис. 616 А). Столбиков и системы каналов нет. Эоцен - ныне. Шесть родов.
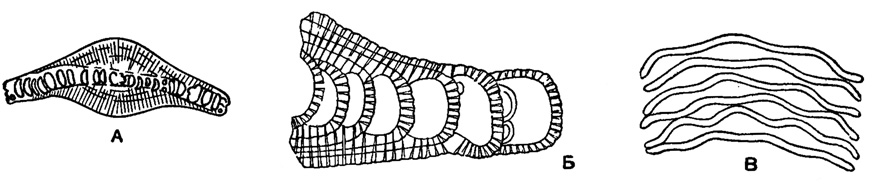
Рис. 616. Сем. Planorbulinidae. А-В Linderina brugesi Schlumberger; А - двояковыпуклая, изогнутая однослойная раковина с утолщением стекловатого вещества над начальной частью с обеих сторон раковины; Б - поровые каналы пронизывают перегородки и добавочное стекловатое вещество, расширяясь кверху; поверхность раковины шероховатая; В - разрез перпендикулярный осн, волнообразно изогнутые перегородки (Schlumberger, 1893)
Дискоидальная форма, бугорки на поверхности у родов Planorbulinella и Linderina, а также расширяющиеся к поверхности поровые каналы, пронизывающие толстую стенку камер второго рода, напоминают признаки родов сем. Orbitoididae. Однако, спиральное или трохоидное нарастание начальных камер и характер устьев у Planorbulinella указывают на принадлежность этих родов к сем. Planorbulinidae. Характер строения начальной стадии и наличие хитиноидного прослоя говорят о генетической близости описанного семейства к семействам Rotaliidae и Anomalinidae. Характерны для теплых и мелководных бассейнов.
Planorbulina Orbigny, 1826. Тип рода - Р. mediterranensis Orbigny, 1826; современный, берег Средиземного моря. Раковина прикрепленная, в начальной стадии спиральная, трохоидная, во взрослой стадии камеры расположены кольцеобразно, часто радиально, иногда в последнем обороте беспорядочно нагромождаются друг на друга; эволютная с обеих сторон или более или менее инволютная с брюшной стороны, на которой камеры более выпуклые, чем на спинной стороне. Устье вначале у внутреннего края последней камеры, низкоарковидное. Каждая камера кольцевой стадии на периферии имеет два устья с губами. Стенка тонкая, равномерно пористая (рис. 617-619). Около 12 видов. Эоцен - ныне. Мангышлак, З. Европа, о. Гаити.
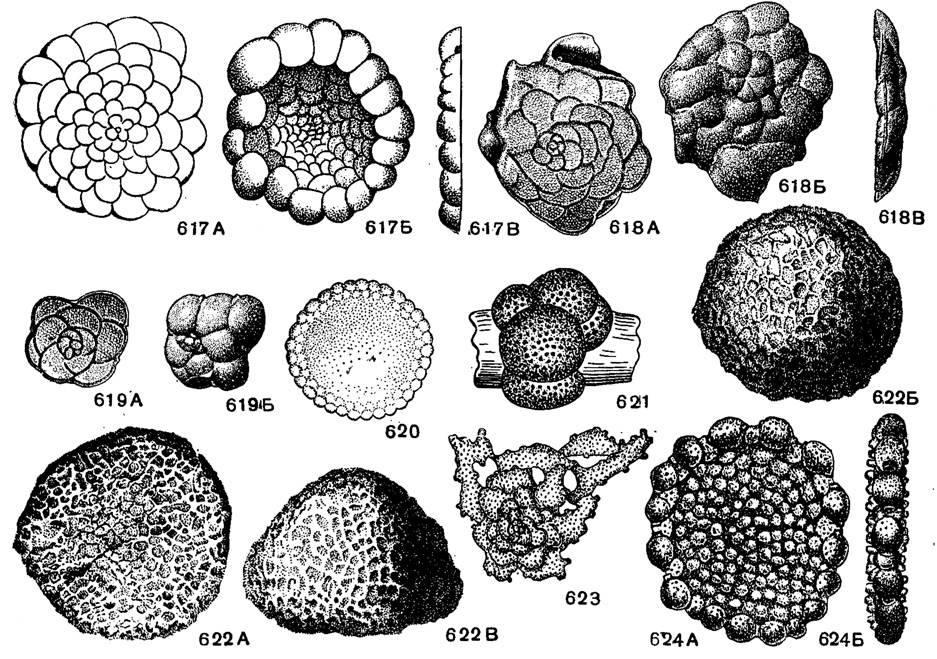
Рис. 617-624. Сем. Planorbulinidae. 617 А-В. Planorbulina mediterranensis Orbigny; внешний вид: А - со стороны прикрепления (спинной), Б - со свободной (брюшной) стороны, В - с периферического края, × 35, современный, Средиземное море (Orbigny, 1826). 618-619. Planorbulina mangyschlakensis Vassilenko; A-В, внешний вид: A - со стороны прикрепления (спинной), Б - со свободной (брюшной) стороны, В - с периферического края; 619 А-Б, внешний вид: А - со стороны прикрепления (спинной), Б - со свободной (брюшной) стороны, × 50, олигоцен, н. Майкоп, п-ов Мангышлак (В. П. Василенко, 1958). 620. Linderina brugesi Schlumberger; внешний вид, × 18, эоцен, Франция (Schlumberger из Galloway, 1933). 621. Acervulina inhaerens Schultze; внешний вид, × 60, современный, Италия (Schultze из Cushman, 1927). 622 А-В. Gypsina vesicularis (Parker et Jones); внешний вид: A - со стороны прикрепления, Б - со свободной стороны, В - с периферического края, × 30; современный, Атлантический океан (Brady, 1884). 623. Planorbullnoides retinaculata (Parker et Jones); внешний вид, × 7, современный. Индийский океан (Parker and Jones, 1865). 624 A-Б. Planorbulinella larvata (Parker et Jones); внешний вид: A - с боковой стороны, Б - с периферического края, × 15, современный, Индийский океан (Parker and Jones, 1865)
Linderina Schlumberger, 1893. Тип рода - L. brugesi Schlumberger, 1893; в. эоцен, Франция. Раковина чечевицеобразная, в начале спирально-плоскостная с волнообразно изогнутыми перегородками (рис. 616 В), не всегда образующими ясные камеры; позднее камеры нарастают в один ряд кольцеобразно, перемежаясь в соседних рядах. Начальные обороты закрыты толстым слоем прозрачного скелетного вещества (рис. 616 А). Стенка толстая, гладкая или шероховатая; перегородки пронизаны тонкими каналами, расширяющимися к поверхности в крупные поры (рис. 616 Б), на периферическом крае они являются устьями (рис. 620). Единичные виды. Кампан Франции; эоцен Европы, Африки, Индии, Японии, о-ва Борнео.
Acervulina Sсhultzе, 1854. Тип рода - A. inhaerens Schultze, 1854; современный, Италия. Раковина свободная или прикрепленная, в начальной стадии плоско-спиральная, более поздние камеры сильно вздутые, шаровидные, нагромождаются друг на друга и закрывают собой начальные камеры. Устья образуются крупными порами. Стенка тонкая, крупнопористая (рис. 621). Шесть видов. В. мел - современные. Германия, Флорида, Филиппинские о-ва.
Gypsina Carter, 1877. Тип рода - Orbitolina vesicularis Parker et Jones, 1860; современный, Атлантический океан. Раковина дискоидальная, полусферическая или слабо коническая, в начальной стадии прикрепленная, с крупной шаровидной камерой, окруженной округлыми спирально нарастающими камерами; в поздней стадии они нарастают кольцеобразно и радиально, временами чередуясь и расширяясь к периферии (рис. 625 А), соединяясь непористой частью стенки (625 Б, б), иногда с просветом (625 Б, в, в1). Устья в виде крупных пор. Стенка в верхней части многослойная (рис. 622; 625 В). Около 10 видов. Эоцен Карпат, Польши, Перу; олигоцен - миоцен Италии, Египта, о. Гаити, Австралии. Современные.
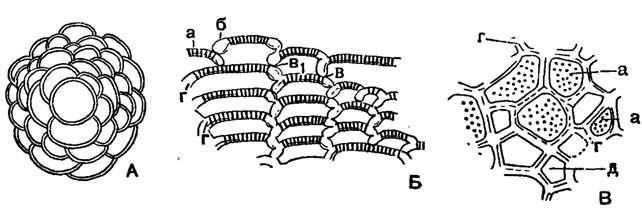
Рис. 625. А-В Gypsina globulosa Reuss: А - нарастание камер начальной части; Б - пористая (а) и не пористая (б) части стенки и соединение смежных камер; без просвета (в) и с просветом (в1), внутренний слой в стенке камер (г); В - поперечный шлиф через нижнюю часть камер; многоугольная форма камер (д), пористость (а) и многослойность (г) их стенки (Uhlig, 1886)
Вне СССР: Planorbulinoides Cushman, 1928 (рис. 623); Planorbulinella Cushman, 1927 (рис. 624).
Род Borodinia Hanzawa, 1949 в семейство не включен, так как его систематические признаки не достаточно ясны. Роды Eoanularia Cole et Bermudez, 1944 и Archaecyclina Silvestri, 1910, судя по строению их начальной части, относятся к сем. Orbitoididae.
Раковина на ранней стадии трохоидная, прикрепленная плоской или вдавленной спинной стороной, которая у некоторых родов изменяется в непористый бесструктурный диск; поздние камеры нарастают трохоидно, конусом или по булиминоподобной спирали, столбиком, однорядно или двурядно. Устье щелевидное или полукруглое, с губой, в основании септальной поверхности последней камеры на брюшной стороне; у взрослых особей неправильно-округлое на вершине конуса, конечное с губой или на конце шейки. Стенка толстая, равномерно крупнопористая, в начальной части многослойная, гладкая, редко на поверхности покрыта посторонними частицами. В. мел?, эоцен - ныне. Четыре рода. В СССР неизвестны.
Трохоидное строение начальной части, отсутствие системы каналов и эволютное или инволютное нарастание камер на брюшной стороне указывают на родство этого семейства с сем. Anomalinidae; однако, сидячий образ жизни привел к большому разнообразию в нарастании поздних камер, к развитию устья в начальной стадии только на брюшной стороне и к преобразованию конечного устья на поздней стадии, а также к сильному утолщению и видоизменению стенки спинной стороны. Роды Bermudezella и Neocarpenterella, видимо, тесно связаны с Carpenteria, но являются менее специализированными по сравнению с ним и родом Rupertia. Представители характерны для холодных или теплых, но обычно мелких вод.
Carpenteria Gray, 1858 (Dujardinia Gray, 1858). Тип рода - Carpenteria balaniformis Gray, 1858; современный, Филиппинские о-ва. Раковина в ранней стадии трохоидная, прикрепленная широко распластанной спинной стороной. Поздние камеры неправильно-треугольные или округлые, вздутые, нарастают по булиминоподобной спирали однорядно или двухрядно, или конусом. Устье вначале щелевидное, на вершине конуса неправильно округлое или округлое на конце шейки у камер однорядной стадии. Стенка грубопористая, толстая, многослойная, внутри камер губчатая (рис. 626-628). Единичные виды. Эоцен Карпат, Польши и С. Каролины; олигоцен Новой Зеландии; миоцен о-вов Порто-Рико, Гаити. Современные.
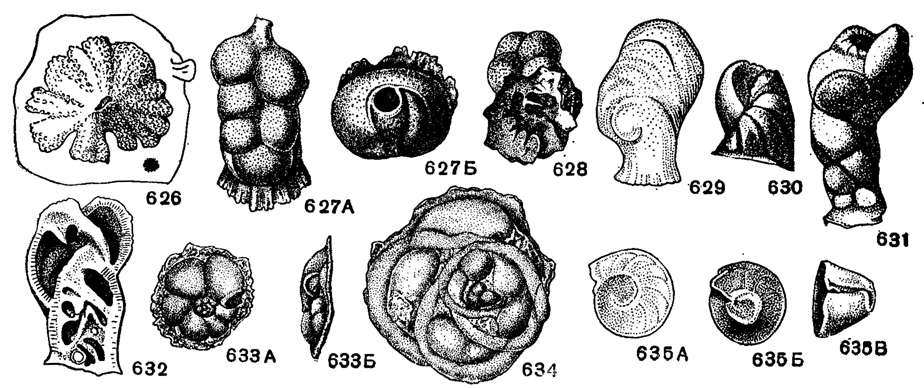
Рис. 626-635. Сем. Rupertiidae. 626. Carpenteria balaniformis Gray; внешний вид раковины, прикрепленной к субстрату, × 21, современный, Филиппинские о-ва (Gray из Galloway, 1933). 627-628. Carpenteria lithotamnica (Uhlig); 627 А-Б, внешний вид: А - сбоку, Б - с устьевого конца; 628 - вид со стороны прикрепления (спинная), × 10, эоцен, Карпаты Польши (Uhlig, 1886). 629-632. Rupertia stabilis Wallich; 629 - внешний вид сбоку, × 21, современный, сев. часть Атлантического океана, Гренландия (Wallich из Galloway, 1933); 630-631 различные стадии роста раковины; 632 - продольное сечение, × 18, современный. Атлантический океан (Brady, 1884). 633-634. Neocarpenteria cubana Cushman et Bermudez, 633 A-Б, внешний вид: A - с брюшной стороны, Б - с периферического края, 634 - внешний вид с брюшной стороны, × 33, в. эоцен, о-в Куба (Cushman and Bermudez, 1936). 635 А-В. Bermudezella truncata (Bermudez); внешний вид: А - со стороны прикрепления (спинная), Б - со свободной (брюшной) стороны, В - с периферического края, × 15, олигоцен, о-в Гаити (Bermudez, 1949)
Rupertia Wallich, 1877. Тип рода - R. stabilis Wallich, 1877; современный, Гренландия. Раковина с резко развитым диском, видоизменившим начальную трохоидную часть. Поздние камеры нарастают по булиминоподобной спирали в виде столбика. Устье в ранней стадии щелевидное, в основании последней камеры, в поздней - открытое, конечное, с губой. Стенка толстая, неравнопористая, многослойная, гладкая или покрыта посторонними частицами (рис. 629-632). Единичные виды. Эоцен Карпат, Польши, Австрии, Флориды. Современные, Гренландия.
В семейство также входят Neocarpenteria Cushman et Bermudez, 1936 (рис. 633-634); Bermudezella Thalmann, 1950 (рис. 635).
Раковина трохоидная, прикрепленная в начальной стадии. По оси навивания образуется пустая полость, закрытая у взрослых особей поздними оборотами; последующие камеры нарастают по конической или трохоидной спирали. Устье щелевидное или треугольное, с губой, иногда закрыто пластинкой с порами или ситовидное. Стенка и септы вначале многослойные; в последней стадии стенка двухслойная. Поровые каналы отчетливые, в виде конических пучков; между ними сплошные столбики, образующие на поверхности бугорки; вдоль углубленных швов иногда выходы септальных каналов. Три рода. Эоцен - ныне. В СССР неизвестны.
Трохоидное строение начальной стадии и спирально-коническое или трохоидное нарастание поздних, чаще вздутых камер, а также наличие столбиков и поровых каналов позволяют предполагать связь этих родов друг с другом и с родом Carpenteria. Однако все указанные признаки изучены еще очень плохо; выделение этого семейства принято условно.
Eorupertia Yabe et Hanzawa, 1925 (Uhligina Yabe et Hanzawa, 1922). Тип рода - Uhligina boninensis Yabeet Hanzawa, 1922; ср. эоцен, Япония. Раковина в начальной стадии прикрепленная, трохоидная. Поздние камеры нарастают по конической или трохоидной спирали, закрывая пустую полость вдоль оси навивания. Устье щелевидное, вдоль внутреннего края камер или одно для нескольких камер одного оборота, с губой и покрыто пластинкой с порами (рис. 641 В). Стенка, пластинка над устьем и септы многослойные; каналы пор сужаются к поверхности и чередуются со сплошными столбиками и каналами вдоль септальных швов (рис. 636-638). Поры на поверхности почти неразличимы, а столбики заканчиваются ясными бугорками (рис. 636-641). Единичные виды. Ср. эоцен Карпат, Польши, Франции, Японии, Ирака, Турции и Кубы.
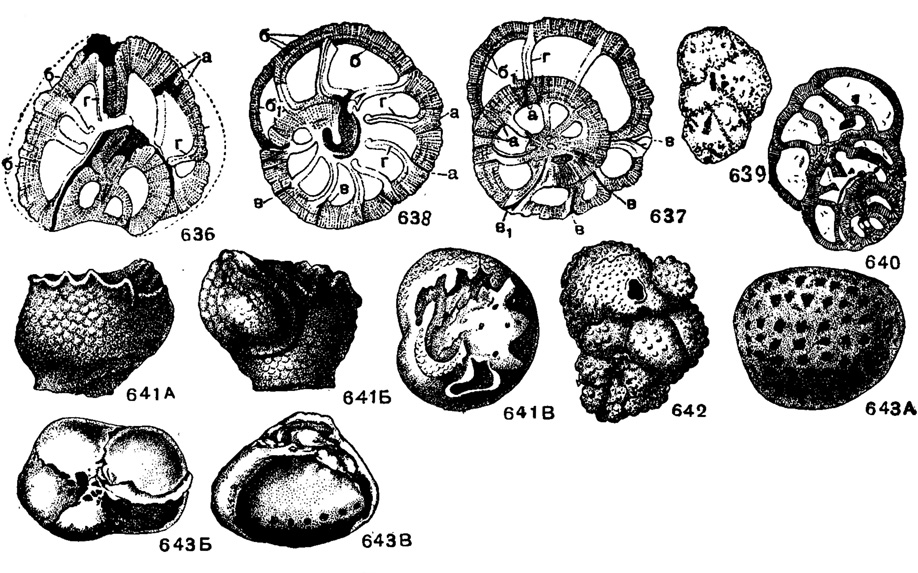
Рис. 636-643. Сем. Victoriellidae. 636-638. Eorupertia incrassata (Uhlig). 636 - продольное сечение через всю раковину; 637 - поперечное сечение через начальную часть раковины; 638 - поперечное сечение через более поздние камеры: а - пучки пор и между ними более плотные столбики, б - слои в спиральной перегородке, разделенные тонкими линиями, б - в септе последней камеры только два слоя, в - круглые, грубые септальные каналы, разветвляющиеся кнаружи, вх - септальный канал, проходящий через два оборота, г - септальные перегородки состоят из двух пластинок, разделенных темной линией. 639-640. Eorupertia boninensis (Yabe et Hanzawa); 639 - вид с боковой стороны, × 8; 640 - продольное сечение, × 12, ср. эоцен, Оки-мура, Япония (Yabe and Hanzawa из Galloway, 1933). 641 А-В. Eorupertia incrassata (Uhlig); внешний вид: А-Б с боковых сторон, В - со стороны устья, × 15, эоцен, Карпаты Польши (Uhlig, 1886). 642. Victoriella plecie (Chapman); внешний вид, × 12, миоцен, Австралия (Chapman, 1921). 643. А-В Hofkerina semiornata (Howchin); внешний вид: А - с нижней поверхности раковины, Б - со стороны пупка, В - с периферического края, × 30, олигоцен, Австралия (Howchin, 18)
Victoriella Chapman et Crespin, 1930. Тип рода - Carpenteria proteiformis Goes var. plectae Chapman, 1921; миоцен, Австралия. Раковина свободная, коническая; камеры немногочисленные, вздутые. Устье полулунное в основании последней камеры, позднее треугольное и лопастное, с губой. Стенка крупнопористая, толстая, в ранней части двуслойная, а в поздней - однослойная; ее пересекают сплошные столбики, образующие на поверхности крупные бугорки (рис. 642). Единичные виды. Олигоцен Франции; миоцен Австралии.
К семейству относится Hofkerina Chapman et Parr, 1931 (рис. 643).
Раковина на ранних стадиях трохоидная, прикрепленная, иногда с мелкими концентрически нарастающими камерами. В поздней стадии растет неправильной массой вверх, разветвляясь на короткие и длинные отростки. Стенка грубо прободенная, имеет каналы и столбики. Специальных устьев нет, их заменяют крупные поры, иногда прикрытые пористыми пластинками (рис. 645). Современные и даже некоторые ископаемые формы окрашены в красный, оранжевый или розовый цвет. Три рода. В. мел - ныне. В СССР неизвестны.
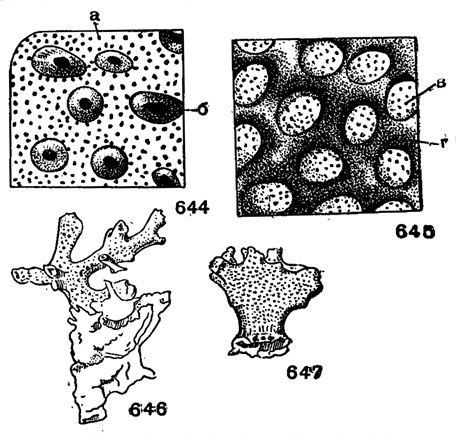
Рис. 644-647. Сем. Homotremidae. 644, 646. Miniacina miniacea (Pallas). 645, 647. Homotrema rubrum (Lamarck). 644-645. Детали поверхности: а - мелкие поры, б - столбики и крупные поры, в - пластинки с мелкими порами, г - непористые промежутки. 646-647 внешний вид, × 3, современные, Индийский океан (Hickson, 1911)
Трохоидное строение начальной стадии раковины и ее прикрепление плоской эволютной стороной указывают на связь этого семейства с формами типа Cibicides; резкая специализация признаков и окраска раковин настолько отличают эту группу фораминифер, что очень долго их считали кораллами. В современных бассейнах встречаются в мелких тропических водах, в особенности в области коралловых рифов.
Miniacina Galloway, 1933 (Polytrema Risso, 1826). Тип рода - Millepora miniacea Pallas, 1776; современный, Индийский океан. Раковина прикрепленная, вначале напоминает Cibicides, поздние мелкие камеры нарастают концентрическими раковина стелется по субстрату. В последней стадии камеры растут неправильной ветвистой) массой с тонкими отростками. Стенка со столбиками и крупными порами; мелкие поры рассыпаны между ними (рис. 644). Цвет красный, яркорозовый или белый (рис. 646). Несколько видов. В. мел. Триполитании. Современные.
К семейству относятся Homotrema Hickson, 1911 (рис. 647) и Sporodotrema Hickson, 1911.
Раковина свободная, на ранних стадиях трохоидная, на поздних - правильно или неправильно циклическая, с многочисленными, часто шаровидными камерами, расположенными в один или несколько оборотов; последняя камера иногда сильно раздута. Устье на ранних стадиях одно, брюшнокраевое, на поздних - множественное, в виде маленьких шовных пор на обеих сторонах, либо ситовидное. Стенка у некоторых на ранних стадиях, по-видимому, хитиноидная. Пять родов. Сенон - ныне. Европа, Америка. В СССР неизвестны.
Cymbalopora Нagеnow, 1851. Тип рода - С. radiata Hagenow, 1851; маастрихт, Голландия. Раковина низкоконическая, на ранних стадиях трохоидная, на поздних - с кольцевым или неправильным расположением камер; на спинной стороне поздние камеры частично объемлют ранние, на брюшной - камеры уплощенные и вытянутые радиально по направлению от периферии к широко открытому пупку. Устье на ранних стадиях одно, брюшнокраевое, на поздних - устьев много, в виде пор в углублениях швов и на внутреннем пупочном крае камер. Стенка на ранних стадиях хитиноидная, на поздних - известковая, грубопористая (рис. 648-649). Единичные виды. Сенон Европы, миоцен Египта. Современные.

Рис. 648-657. Сем. Cymbaloporidae. 648-649. Cymbalopora radiata Hagenow; 648 А-Б, внешний вид: А - сверху, Б - снизу; 649 - ранняя часть раковины, × 22, мел, Голландия (Cushman, 1948). 650 А-В. Cymbaloporetta squamosa Orbigny; внешний вид: А - сверху, Б - снизу, В - сбоку, × 50, современный (Brady, 1884). 651-652. Cymbaloporella tabellaeformis (H. B. Brady); 651 - внешний вид, × 28; 652 А-Б, внешний вид: А - сверху, Б - сбоку, × 40, современный (Brady, 1884). 653-657. Tretomphalus bulloides (Orbigny); внешний вид: 653 А - сверху, 653 Б - сбоку, 654 - сбоку, 655 - снизу, 656-657 - сбоку, × 60, современный (Brady, 1884)
Cymbaloporetta Cushman, 1928. Тип рода - Rosalina squamosa Orbigny, 1839; современный, З. Атлантика. Раковина высококоническая, на ранних стадиях трохоидная, на поздних - с кольцевым расположением камер. На спинной стороне камеры частично объемлющие, между поздними камерами на брюшной стороне имеются углубления, заполняемые камерами следующего кольцевого ряда. Пупок закрыт пластинкой. Устье у взрослых из ряда мелких округлых отверстий по шовным краям камер и на их пупочном крае, ниже пластинки, прикрывающей пупок. Стенка на ранних стадиях хитиноидная, на поздних - известковая (рис. 650). Единичные виды. Миоцен - ныне. Индо-Тихоокеанская обл.
Cymbaloporella Cushman, 1927. Тип рода - Cymbaloporatabellaeformis Н. В. Brady, 1884; современный, Индийский океан, воет, побережье Мадагаскара. Раковина уплощенная, на ранних стадиях трохоидная, с частично объемлющими камерами, на поздних - с кольцевым расположением камер, вытягивающихся в спинно-брюшном направлении (вертикальные камеры). Пупок открытый. Устье у взрослых состоит из многочисленных мелких отверстий на брюшной стороне, вдоль швов. Стенка на ранних стадиях хитиноидная, коричневая, на поздних - известковая, грубопористая (рис. 651-652). Эоцен - ныне. Индо-Тихоокеанская обл.
Tretomphalus Мoеbius, 1880. Тип рода - Rosalina bulloides Orbigny, 1839; современный, Атлантический океан, около о-ва Гаити. Раковина на ранних стадиях роталоидная, на поздних - с большой шаровидной камерой, содержащей внутреннюю камеру с тонкой стенкой, которая может заполняться газом. Устье - ряд округлых отверстий, типа пор, на наружной поверхности шаровидной камеры. Стенка на ранних стадиях хитиноидная, на поздних - известковая, пористая (рис. 653-657). Специализированный пелагический род. Единичные виды. Третичные отложения. Современные.
Род Pyropilus Cushman, 1934, известен пока только в современных морях. Америка, Тихий и Атлантический океаны.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'