
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Надсемейство Rotaliidea Reuss, 1860
[nom. transl. Glaessner, 1945 (ex Rotaliidae Reuss, 1860)]
Раковина свободная, трохоидная, на поздней стадии иногда спирально-плоскостная, редко в последующем однорядная, у всех с системой каналов, иногда с двойными септами. Устье без губы, септальное, в виде щели или ряда пор, или ситовидное; у трохоидных - в основании септальной поверхности, которая в то же время является брюшной стороной камер, у спирально-плоскостных - медианное или септальное у основания септы или по всей септальной поверхности, у развернутых - конечное. Устье иногда первоначально отсутствует и образуется путем резорбции стенки у основания септы, перед образованием следующей камеры. Сообщение между камерами происходит через межкамерные форамены, простые или усложненные вторичными перегородками камер. Последняя камера у некоторых всегда тонкостенная и у ископаемых редко сохраняется. Стенка радиально-лучистая, слоистая; дополнительные скелетные образования в виде пупочного диска, бугорков, киля, мостиков, шовных утолщений и пр. Два семейства: Rotaliidae и Elphidiidae. В. мел - ныне.
Раковина трохоидная, линзовидная или коническая, с выпуклой спинной и уплощенной брюшной сторонами. Межкамерные форамены усложнены иногда перегораживающим их желобообразным тяжем (аналог зубной пластинки Хофкера). На обращенной к пупку стороне каждая камера с пупочным отверстием (пупочное устье Смаута, 1954), по которому осуществляется соединение полости камеры с пупочным каналом или при его отсутствии непосредственно с пупочной областью. Шесть родов. В. мел - ныне.
Rotalia Lamarck, 1804. Тип рода - Rotalites trochidiformis Lamarck, 1804; cp. эоцен, Франция. Раковина обычно с простыми камерами; у некоторых видов есть дополнительные камерки близ периферии. В центре брюшной стороны пупочная шишка, разбитая трещинами на многочисленные столбики, расположенные ярусами; в каждом обороте свой ряд столбиков. Сверху пупочная сторона покрыта сплошь скелетным веществом, под которым проходит пупочный канал; у некоторых видов имеются внутрисептальные каналы, впадающие в пупочный, у других камеры через пупочное отверстие непосредственно по щели соединяются с пупочным каналом (рис. 697-699). Десятки видов. В. сенон - ныне. Широко распространенный род.
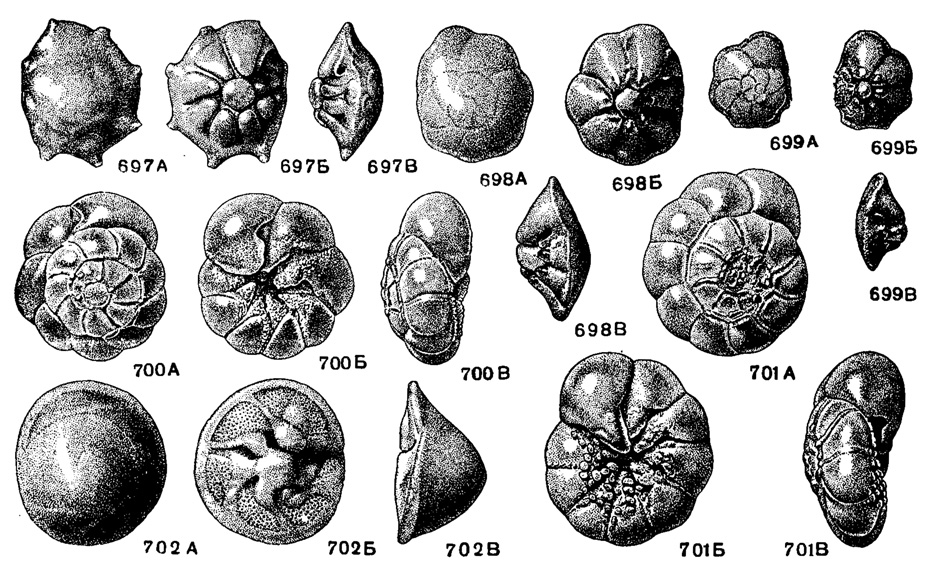
Рис. 697-702. Сем. Rotalliidae. 697 А-В. Rotalia (Pararotalia) armala (Orbigny); внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 47, ср. эоцен, бучакский ярус, Украина, Никопольский р-н (колл. М. В. Ярцевой). 698-699. Rotalia (Pararotalia) inermis (Terquem); 698 А-В, внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 47; 699 А-В. внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 47, лютетский ярус, Парижский бассейн (колл. Н. Н. Субботиной). 700-701. Streblus beccarii (Linne); 700 А-В (форма А), внешний вид: А - со спинной стороны, Б - с брюшной стороны. В - с периферического края; 701 А-В (форма В), внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 33, современный. Черное море у Карадага (колл. Н. Н. Субботиной). 702 А-В. Rotorbinella fungiformls Subbotlna; внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 72, в. эоцен, Абхазия (Субботина, 1958)
Подрод: Pararotalia Le Calvez, 1949.
Streblus Fisher, 1817. Тип рода - Nautilus beccarii Linne, 1758; современный, Средиземное море. Отличается от Rotalia отсутствием сплошного покрова скелетного вещества, закрывающего пупок и пупочные щели около камер. К центру каждая камера на брюшной стороне вытягивается в виде лопасти, покрытой многочисленными мелкими столбиками. Настоящего устья нет. Имеется межкамерный форамен. Перед образованием новой камеры септальная поверхность, судя по неровным краям и ассиметричной форме щели, по-видимому, прорывается. В дальнейшем образуется желобообразный тяж (зубная пластинка Хофкера), соединяющий брюшную и спинную части камеры; он расположен немного впереди септы. Сообщение с внешней средой происходит через открытую пупочную область. Имеются пупочные отверстия на пупочных концах камер, ведущие через щель (аналог пупочного канала) в пупочную область (рис. 700-701). Несколько разновидностей одного вида. Миоцен - ныне. Широко распространен, особенно на юге Европы и в США.
Rotorbirtella Bandy, 1944. Тип рода - R. calliculus Bandy, 1944; эоцен, Орегон. Раковина с высоковыпуклой спинной и уплощенной брюшной сторонами, иногда с пластинчатым килем, в центре брюшной стороны со сплошной пупочной шишкой. Швы изогнутые, часто с изломом посредине, валиковидные, расширенные около шишки и суженные к периферическому краю. Устье - щель без губы, протягивается вдоль краевого шва, немного не достигая периферического края (рис. 702). Примерно 10 видов. Третичные Крыма, Абхазии, З. Европы, С. Америки. Современные.
Вне СССР: Lockhartia Davies, 1932; Dictyoconoides Nuttall, 1925; Sakesaria Davies, 1937.
[nom. transl. Sigal, 1952 (ex Elphidiinae Galloway, 1933)]
Раковина свободная, асимметричная трохоидная, реже полуинволютная или двусторонне-симметричная, большей частью инволютная, редко эволютная. Система каналов, большей частью с выходами наружных отростков меридиональных каналов в ямки, или на швах, или параллельно им. Для некоторых форм характерны длинные межсептальные мостики с соответствующими им межсептальными ямками, для других - очень короткие утолщения между углублениями на швах (септальные мостики и ямки). Устье часто отсутствует; форамен в виде округлых отверстий, реже щелевидный, в основании септальной поверхности, иногда с редкими добавочными отверстиями на септах или ситовидный. 13 родов. Третичные - ныне. Происходит, вероятно, от представителей сем. Rotaliidae путем развития инволютной раковины и двусторонне-симметричной системы каналов.
Характеризуется развитием ретральных отростков камер с полыми трубками внутри, открывающимися с одного конца в полость камер и замкнутыми с другого конца; на поверхности стенки им соответствуют возвышения - межсептальные мостики. Отростки меридиональных каналов открываются в межсептальные ямки вдоль швов; иногда имеется килевой канал. Устье часто отсутствует. Форамен в виде одного ряда отверстий в основании септ, иногда с добавочными отверстиями на септах. Пять родов.
Faujasina Orbigny, 1839. Тип рода - F. carinata Orbigny, 1839; в. мел, Голландия (?). Раковина трохоидная, с плоской спинной и выпуклой брюшной сторонами (рис. 703). Один вид. По Орбиньи, в. мел Голландии; по Кешмэну (Cushman, 1948) - с плиоцена до ныне.

Рис. 703-710. Сем. Elphidiidae (подсем. Elphidiinae). 703 А-В. Faufasina carinaia Orbigny; внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 75, в. мел, Голландия (Orbigny, 1846). 704-705. Faujasinella semiinvoluta Mjatliuk; 704 А-В, внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, 705 - внешний вид со спинной стороны, × 75, в. эоцен, Предкарпатье (Мятлюк, 1956). 706 А-В. Notorotalia australis (Cushman); внешний вид: А - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 45, современный, Австралия (Cushman, 1939). 707. Elphidium crisрum (Linne); схематизированное аксиальное сечение (схема строения каналов): кк - килевой канал, ск - спиральный канал, пк - пупочные каналы, мк - меридиональные каналы, ро - ретральные отростки, но - наружные отростки мернд. канала, ф - форамен, × 70. 708. Elphidium macellum (Fichtel et Moll); строение межсептальных мостиков и ямок: ро - ретральные отростки, оно - отверстие наружного отростка мерид. канала, × 102, сармат, С. Кавказ (колл. А. К. Богдановича). 709 А-Б. Elphidium macellum (Fichtel et Moil); внешний вид: А - сбоку, Б - с периферического края, × 35, миоцен, зап. обл. Украины (колл. А. К. Богдановича). 710 А-Б. Planoelphidium laminalum (Terquem); внешний вид: А - со спинной стороны, Б - с периферического края, × 50, плиоцен, Албания (колл. Е. А. Храмой)
Faujasinella Voloshinova, 1958. Тип рода - Elphidium semiinvolutum Mjatliuk, 1956; н. олигоцен, Закарпатье. Раковина полуинволютная, не вполне симметричная, двояковыпуклая (рис. 704-705). Один вид. В. эоцен Восточных Карпат и н. олигоцен Закарпатья.
Notorotalia Finlay, 1939. Тип рода - N. zealandica Finlay, 1939; современный, у берегов Новой Зеландии. Раковина трохоидная, большей частью двояковыпуклая. В основании септ расположен ряд округлых отверстий. На поверхности с обеих сторон выпуклая сетчатая скульптура (рис. 706). Несколько видов. Эоцен - ныне. Новая Зеландия, Америка, Современные - в Индо-Тихоокеанской обл. Систематическое положение рода не вполне ясно.
Elphidium Montfort, 1808 (Geophonus Montfort, 1808; Sporilus Montfort, 1808; Themeon Montfort, 1808; Polystomella Lamarck, 1822). Тип рода - Nautilus macetlus Fichtel et Moll, 1798; современный, Средиземное море. Раковина инволютная, чечевицеобразная, двусторонне-симметричная, часто килеватая, иногда с периферическими шипами, с 10-30 седлообразными камерами в последнем обороте (рис. 707-709). Около 50 видов. В. эоцен (?) - ныне. Повсеместно.
Planoelphidium Voloshinova, 1958. Тип рода - Polystomella laminata Terquem, 1878; в. плиоцен, о. Родос. Раковина двусторонне-симметричная, плоская, полностью эволютная (рис. 710). 1-2 вида. Плиоцен - ныне. Средиземноморская область и южная часть Тихого океана.
Ретральные отростки и соответствующие им межсептальные мостики отсутствуют; швы большей частью либо слегка углубленные с септальными ямками и мостиками, либо плоские, без мостиков, с двойным рядом отверстий отростков меридиональных каналов, расположенных по обе стороны от шва. У некоторых родов имеются внутренние отростки меридиональных каналов. Устье выражено редко. Форамен большей частью в виде ряда отверстий, реже щелевидный в основании септальной поверхности, часто ситовидный. Семь родов.
Porosorotalia Voloshinova, 1958. Тип рода - Notorotalia clarki Voloshinova, 1952; миоцен, Сахалин. Раковина трохоидная, двояковыпуклая, с зернистостью в пупочной области и с выпуклыми швами. Спиральный канал и длинные анастомозирующие пупочные каналы. Форамен в виде ряда округлых отверстий в основании септ (рис. 711). 2-3 вида. Эоцен Австралии, миоцен Сахалина; плиоцен Америки.
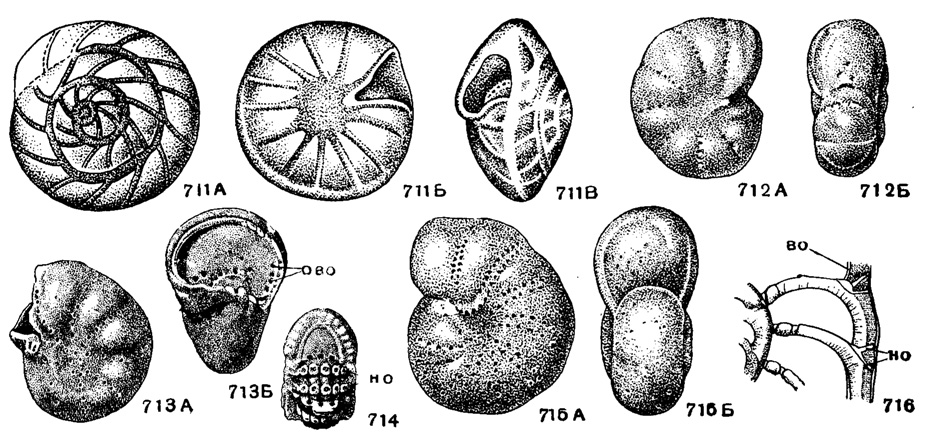
Рис. 711-716. Сем. Elphidiidae (подсем. Cribroelphidiinae). 711 А-В. Porosorotalia clarki (Voloshinova); внешний вид: A - со спинной стороны, Б - с брюшной стороны, В - с периферического края, × 30, миоцен, Сахалин (Волошинова, 1952). 712 А-Б. Cribroelphidium vulgare (Voloshinova); внешний вид: А - сбоку, Б - с периферического края, × 50, миоцен, Сахалин (Волошинова, 1952). 713 А-Б. Cribroelphidium subglobosum (Voloshinova); внешний вид экземпляра с поломанной последней камерой: А - сбоку, Б - с периферического края, ово - отверстия внутренних отростков меридиональных каналов, × 50, миоцен, Сахалин (Волошинова, 1956). 714. Cribroelphidium sp.; строение наружных отростков меридиональных каналов: но - наружные отростки мерид. каналов, × 50, миоцен, Сахалин (Волошинова, 1956). 715-716. Elphidiella arctica (Parker et Jones); 715 А-Б, внешний вид: А - сбоку, Б - с периферического края, × 35; 716 - строение отростков меридиональных каналов: во - внутренние отростки мерид. каналов, но - наружные отростки мерид. каналов, × 100, современный, Карское море (Волошинова, 1956)
Cribroelphidium Cushmanet Bronnimann, 1948. Тип рода - C. vadescensCushman et Bronnimann, 1948; современный, Карибское море. Раковина инволютная, двусторонне-симметричная, с 8-12 камерами в последнем обороте. Швы углубленные, с ямками и отчетливыми септальными мостиками. Внутренние отростки меридиональных каналов открываются в камеру над септой, наружные отростки не разветвляются, выходят в ямки септальных швов. Устье и форамен в виде ряда округлых отверстий, реже очень короткой щели в основании септ, часто ситовидное; устье обычно отсутствует (рис. 712-714). Отличиями от сходного рода Cribrononion Thalmann, 1946 из сем. Nonionidae служат: микроструктура стенки, присутствие внутренних отростков меридиональных каналов и характер устья - у Cribrononion оно не бывает ситовидное. Не менее 20 видов. Эоцен (?), олигоцен - ныне. Сахалин, Япония, Америка.
Elphidiella Cushman, 1936. Тип рода - Polystomella arctica Parker et Jones, 1864; современный, С. Ледовитый океан. Раковина инволютная, редко на взрослых стадиях слегка эволютная, обычно крупных размеров (1-3 мм), часто с большим количеством камер в последнем обороте (15-30). Отростки меридиональных каналов многочисленные; наружные имеют тенденцию к раздваиванию, в последнем случае вдоль швов образуется два ряда отверстий; в случае наличия одного ряда наружных отростков каналов, последние открываются на швах на дне многочисленных очень мелких и частых ямок, разделенных неясными перемычками; внутренние отростки меридиональных каналов открываются внутри камер, как у рода Cribroelphidium. Форамен часто ситовидный, с многочисленными отверстиями в основании устьевой поверхности, иногда разрывающимися и образующими подобие длинной неровной щели (рис. 715-716). Не менее 15-20 видов. Палеоцен - ныне. Сахалин, З. Сибирь, Америка.
Вне СССР: Elphidioides Cushman, 1945. Parrellina Thalmann, 1951; Polystomellina Yabe et Hanzawa, 1923, Ozawaia Cushman, 1931
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'