
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Отряд Nummulitida (И. В. Качарова)
Раковина крупная (до 160 мм), чечевицеобразная или дисковидная, обычно округлого очертания или звездчатая, гладкая или с гранулами, иногда с лучами. Строение раковины спирально-плоскостное, иногда со спирально-трохоидным расположением камер на ранних стадиях развития раковины или концентрическим на поздних. Раковина пористая, обычно со вторичными скелетными образованиями и сложной системой каналов. В. мел - ныне. Семейства: Nummulitidae, Miogypsinidae, Orbitoididae, Discocyclinidae и Lepidocyclinidae.
У концентрических форм в хорошо ориентированном экваториальном сечении, проходящем по плоскости симметрии, различают стадии роста раковины: 1) эмбриональную, 2) непионическую и 3) неаническую с эфебической.
Эмбриональная стадия. Эмбрион (нуклеоконх) у форм В (или микросферических) однокамерный, у форм А (макросферических) он часто двухкамерный; первая камера, протоконх, сферической или субсферической формы, обычно более маленькая, чем вторая камера - дейтероконх. Они отделены друг от друга прямой или изогнутой перегородкой (стенкой) (рис. 717).
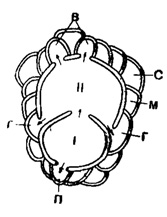
Рис. 717. Схематическое строение эмбриона и периэмбриональных камер: I - протоконх; II - дейтероконх; стрелки показывают столоны, соединяющие камеры эмбриона с главными вспомогательными камерами (г), побочными (адоксилярными) вспомогательными камерами (б) и акцессорной (протоконхальной) камерой (n); остальные камеры - межвспомогательные: с - симметричная межвспомогательная камера, м - асимметричная межвспомогательная камера (Bronnimann, 1940)
Двухкамерный эмбрион обычно одного из следующих типов: а) изолепидиноидный, состоящий из протоконха и дейтероконха приблизительно равного размера; б) нефролепидиноидный - с дейтероконхом, охватывающим половину окружности протоконха; в) эвлепидиноидный - с дейтероконхом, охватывающим протоконх целиком, за исключением места прикрепления последнего к внутренней стенке дейтероконха; г) триблиолепидиноидный, похожий на эвлепидиноидный, но со стенкой протоконха, перпендикулярной к стенке дейтероконха (рис. 718). Встречаются и многокамерные эмбрионы.
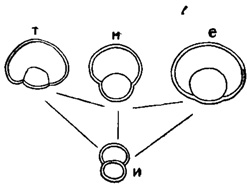
Рис. 718. Эмбриональные камеры Lepidocyclina в экваториальном сечении: и - изолепидиноидная, н - нефролепидиноидная, е - эвлепидиноидная, т - триблиолепидиноидная (Glaessner, 1948)
Непионическая стадия. Первое кольцо камер, называемых периэмбриональными (или непионическими), окружает эмбрион частично или полностью (рис. 717). По расположению и характеру связи их с нуклеоконхом можно различить:
А. Вспомогательные камеры, связанные с нуклеоконхом отверстиями (столонами). Среди них различаются: а) главные вспомогательные камеры, соприкасающиеся базальными частями своей стенки и протоконха и дейтероконха, но сообщающиеся только с последним; б) побочные (адоксилярные) вспомогательные камеры, связанные с дейтероконхом; в) акцессорные, или протоконхальные камеры, имеющие связь только с протоконхом.
Б. Межвспомогательные камеры, образующиеся от вспомогательных камер. Одна базальная часть стенки такой камеры, носящей название асимметричной, касается нуклеоконха, а другая - стенки вспомогательной или предшествующей межвспомогательной камеры. Последняя, если стенка ее соприкасается со стенками двух смежных с ней межвспомогательных камер, носит название симметричной камеры.
Непионические камеры (непионт) с одной главной вспомогательной камерой, из которой развиваются межвспомогательные камеры, образующие одну спираль (Miogypsina, рис. 742) считаются примитивными. Двухспиральный непионт характеризуется одной главной вспомогательной камерой, которая дает начало двум спиралям, направляющимся в противоположные стороны.
Трех- и четырехспиральные непионты имеют две (асимметричные или симметричные) главные вспомогательные камеры, из которых выходят три или четыре спирали (Orbitocyclinoides). Бывают и многоспиральные непионты, в сложении которых принимают участие и побочные вспомогательные камеры. Непионт и эмбрион вместе составляют ювенариум.
Неаническая (и эфебическая) стадия. За периэмбриональными камерами следуют экваториальные камеры, которые расположены кольцеобразно; последние с эмбриональными и периэмбриональными камерами вместе образуют средний, или экваториальный, слой раковины.
Камеры экваториального слоя расположены концентрическими рядами и имеют различную форму: дугообразную, стрельчатую, лопатообразную, ромбовидную или гексагональную (рис. 719, 720). В экваториальном сечении некоторых форм видно, что стенка камер состоит из двух слоев: внутреннего и внешнего, отграниченных друг от друга "темной линией" (Discocyclina, рис. 754).
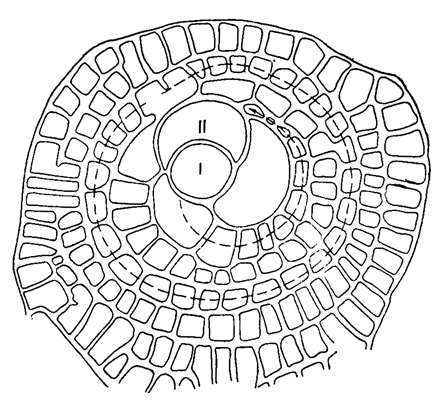
Рис. 719. Cycloclypeus gumbelianus Brady. I-II - начальные камеры (эмбрион); камеры, отмеченные стрелкой - гетеростегиноидные; большая камера, расположенная между последними и эмбрионом - оперкулиноидная; периферические камеры расположены циклично (Renz und Küpper, 1946)
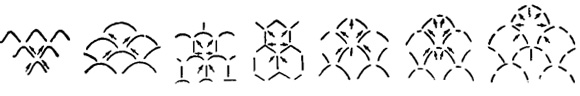
Рис. 720. Форма экваториальных камер и столоны; (слева направо); ромбовидная с 4 столонами (2 диагональных); дугообразная с 4 столонами (2 диагональных); лопатообразная с 6 столонами (2 диагональных + 1 кольцевой); гексагональная с 6 столонами (2 диагональных + 1 кольцевой); стрельчатая с 6 или 8 столонами (2 диагональных + 1 или 2 радиальных); стрельчатая с 8 столонами (2 диагональных + 1 кольцевой + 1 радиальный) (Sigal, 1952)
По обеим сторонам экваториального слоя иногда развиты боковые камеры, неправильно полигональные в сечении, параллельном горизонту или на поверхности раковины, и щелевидные, округлые или прямоугольные в вертикальном сечении (рис. 721-723).
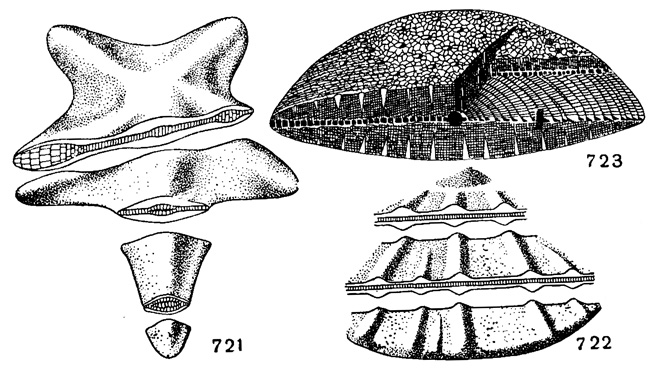
Рис. 721-723. Схемы, показывающие соотношение между экваториальными и боковыми слоями и боковыми камерами. 721. Asterocyclina. 722. Aklinocyclina; незаштрихованная часть поперечного сечения соответствует слоям боковых камер (Bronnimann, 1946). 723. Discocyctina; соотношение между экваториальными и боковыми камерами (Van der, Vlerk Umbgrove, 1927)
Связь между камерами осуществляется через столоны. В филогенетическом ряде форм отмечается возрастание количества столонов: к обычным диагональным столонам прибавляются радиальные, кольцевые, иногда и вертикальные. Различаются следующие типы (рис. 720) систем столонов: а) система из четырех диагональных столонов, б) система из шести столонов (диагональных и кольцевых), в) система из шести - восьми столонов (диагональных и радиальных) и г) система из восьми столонов (диагональных, радиальных и кольцевых). Столоны хорошо различимы в препаратах, пропитанных каким-нибудь красящим веществом или канадским бальзамом (Lepidocyclina, рис. 763).
Связь между камерами происходит и через поры, пронизывающие пол и потолок камер; при этом количество пор всегда больше количества столонов; поперечное сечение последних всегда больше поперечного сечения первых.
В экваториальном сечении спирально-плоскостных форм видны следующие главные морфологические элементы раковины: спиральная пластинка, спиральный канал, ограниченный смежными спиральными пластинками и разделенный перегородками, состоящими из двух слоев. Камеры соединены между собой отверстиями, расположенными в плоскости симметрии раковины у основания перегородок. Начальная камера у форм В весьма маленькая, у форм А она крупная, сферической или субсферической формы, иногда двухраздельная. Форма камер в экваториальном сечении зависит от высоты спирального канала, от наклона и изгиба перегородок и от расстояния между ними.
Дополнительные скелетные образования. Помимо основного скелета, образующего раковину, наблюдаются и вторичные (дополнительные) скелетные образования: краевая хорда (или спиральный валик), столбики и перегородочные ребрышки (или следы септ). Хорда развита в стенке раковины у перегиба спиральной пластинки; она состоит из непрободенных известковых пластинок, сложенных веерообразно; они хорошо различимы в поперечном сечении раковины. В сложении столбиков принимают участие конические или призматические известковые тельца волокнистого строения и соединяющее их известковое вещество. Столбики могут зародиться на разных уровнях завивания спиральной пластинки. Они на поверхности раковины заканчиваются бугорками или гранулами, связанными с перегородочными ребрышками или расположенными в промежутках между ними (рис. 724).
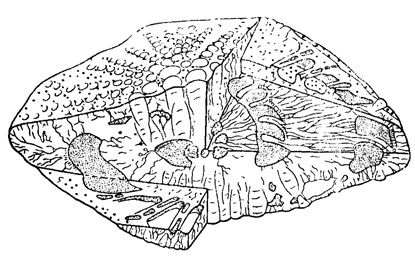
Рис. 724. Схематическая блокдиаграмма Siderolites (S. heracleae Arni) (Sigal, 1952)
Различные части раковины некоторых форм содержат систему канальцев, о строении которой могут дать ясное представление отливы, полученные в результате растворения раковины, пропитанной каким-нибудь нерастворяющимся веществом (канадским бальзамом или парафином). В раковине оперкулины (рис. 725) ясно наблюдаются системы канальцев: внутрихордовая или спиральная (краевая) и внутриперегородочная (интрасептальная). Последняя, соединяя хордовые канальцы соседних спиральных пластинок, образует систему радиальных связей в раковине. В некоторых формах отмечено наличие и других канальцев. Иногда головки столбиков, развитых на местах пересечения перегородочных ребрышек (следов септ) с хордой предшествующего оборота, располагаются на поверхности раковины спирально и образуют так называемую трансверсальную пластинку. В стенке некоторых форм различимы короткие и тонкие ответвления перегородочных ребрышек, так называемые поперечные трабекулы, соответствующие вторичным перегородкам гетеростегинид и представляющие собой начальную стадию усложнения перегородочных ребрышек.
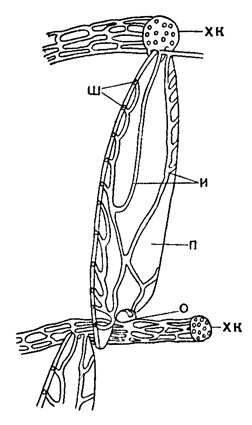
Рис. 725. Система канальцев у Operculina: хк - хордальные канальцы, и - интрасептальные канальцы, ш - шовные поры, п - перегородка, о - перегородочное отверстие (Sigal, 1952)
Простыми (радиальными) считаются перегородочные ребрышки (или следы септ), отходящие от центра раковины и направляющиеся к ее периферии. Они могут быть прямые, волнистые и вихреобразно закрученные. Более сложны меандрические перегородочные ребрышки, образующиеся в результате обособления на поверхности раковины пучков сильно извилистых и параллельных ребрышек с несколькими центрами расхождения. На поверхности некоторых форм эти параллельно тянущиеся ребрышки, сходясь местами, образуют сетчато-меандрический тип ребрышек, состоящий из редких и удлиненных петель неправильной формы. Наиболее сложно построенным типом считаются сетчатые перегородочные ребрышки с короткими и близко друг к другу расположенными петлями правильной формы. Эти ребрышки у мегалосферических форм и форм В на ранней стадии развития простые, а у взрослых микросферических - более сложные.
Важным критерием для установления крупных подразделений в группе спирально - плоскостных форм считается степень инволютности спиральной пластинки и камер (рис. 726), а также характер строения камер и спирального валика хорды.
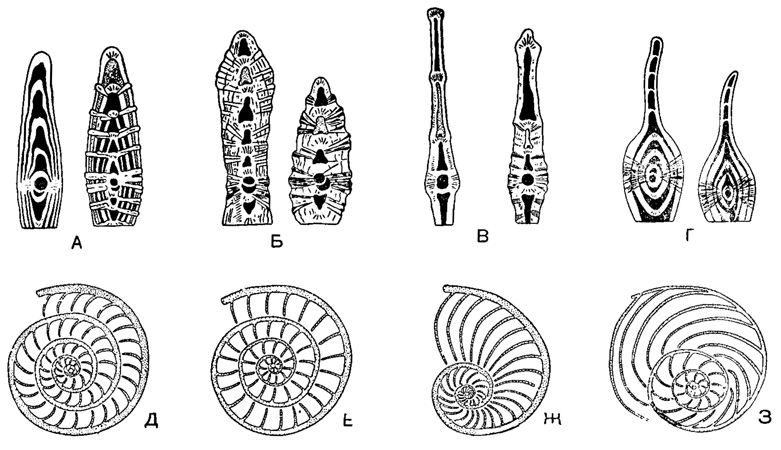
Рис. 726. Строение раковин спирально-плоскостных форм подсем. Nummulitinae в осевых (А-Г) и экваториальных (Д-З) сечениях: А-Д - Nummulites; Б, Е - Assilina, В, Ж - Operculina; Г, З - Operculinella (по Немкову, 1955)
Для систематики группы с концентрическим расположением камер разные авторы применяют различные морфологические элементы. Так, например, Ван де Гейн (Van de Geyn) и Ван дер Флерк (Van der Vlerk) для классификации использовали систему канальцев, Шеффен (Scheffen) - боковые камеры. Однако дальнейшие исследования показали, что каждый из этих признаков меняется даже у представителей одного и того же вида; кроме того, некоторые из этих морфологических элементов еще недостаточно хорошо изучены. Поэтому более правильной приходится признать систематику, основанную на использовании нескольких признаков вместе.
Представители отряда Nummulitida имеют важное стратиграфическое значение. Верхнемеловые представители этого отряда развивались в зоне Тетиса и только в З. Европе они дошли до Голландии. В третичном периоде их ареал распространения несколько расширился: кроме Тетиса, присутствие их отмечено в южных частях Русской платформы до Днепропетровска, в Ср. Азии и в Приаралье, а также в отложениях Центр. Европы, Ю. Австралии, Мадагаскара, Новой Зеландии.
(Раздел "Семейство Nummulitidae" составлен Г. И. Немковым.)
[nom. transl. Schubert, 1920 (ex Nummulitinae Carpenter, 1859)]
Раковина двусторонне-симметричная, чечевицеобразная или дисковидная, состоит из ряда оборотов, разделенных септами на многочисленные камеры. Обороты инволютные и эволютные, на ранних стадиях роста в большинстве случаев инволютные. Камеры располагаются обычно спирально, редко концентрически. Неперфорированные элементы раковины (септы, спиральный валик, столбики и гранулы) имеют сложную систему мелких канальцев. В. мел - ныне. Подсемейства: Nummulitinae, Miscellaneinae, Siderolitinae и Heterostegininae.
Филогения сем. Nummulitidae изучена недостаточно. Имеющиеся схемы филогении нуммулитид, предложенные Р. Шубертом (Schubert, 1909), Д. Кешмэном (Cushman, 1928) и Д. Гэлловеем (Galloway, 1933), содержат серьезные ошибки и не могут быть приняты даже за основу. Наши представления о филогении сем. Nummulitidae показаны на приведенной ниже схеме (рис. 727).
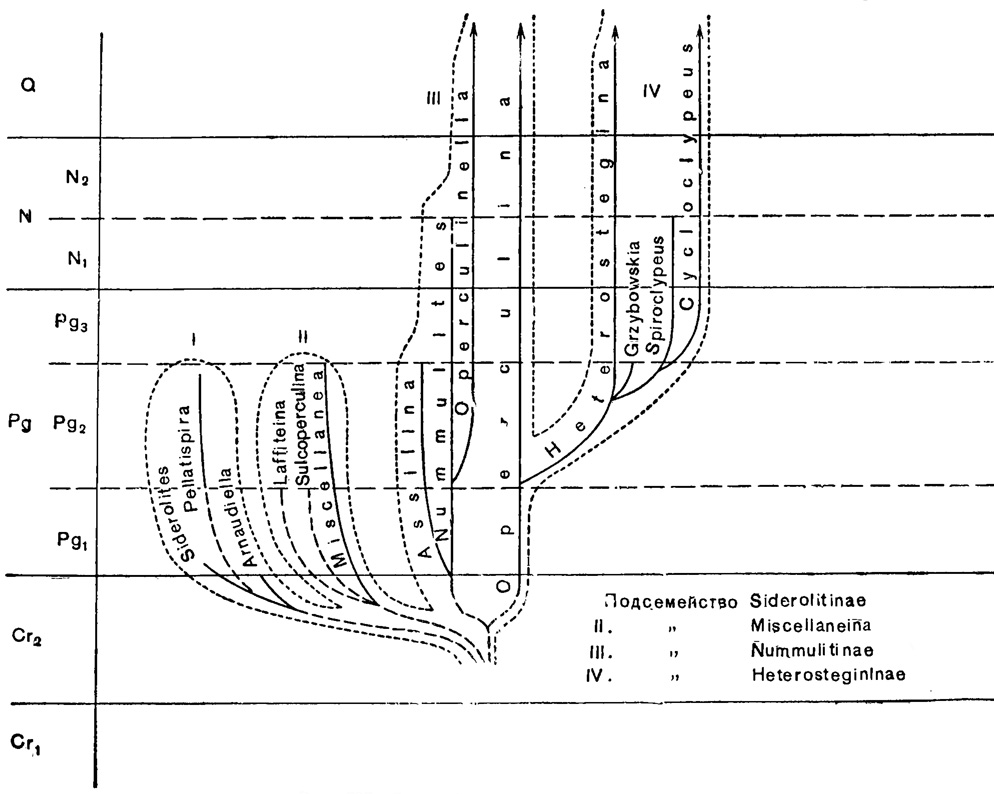
Рис. 727. Схема филогении сем. Nummulitidae
Полость спирального канала раковины разделена многочисленными септами на простые камеры; спиральный валик хорошо развит. Четыре рода (рис. 726).
Nummulites Lamarck, 1801* (Camerina Bruguiere, 1792; Lenticulites Lamarck, 1804; Egeon Montfort, 1808; Nummularia Sower by, 1826; Nummulina Orbigny, 1826). Тип рода - Catnerina laevigata Bruguiere, 1792; средний эоцен, Франция. Раковина крупная (от 1 до 160 мм), чечевицеобразная (мелкие формы) или дисковидная (крупные), вздутая в различной степени в центре. Обороты и полости камер инволютны. Многочисленные спиральные обороты разделены септами на простые камеры (рис. 728, 732). Септы обычно дугообразно изогнуты, редко прямые. На поверхности оборотов в местах прикрепления септ образуются прямые или извилистые линии в виде швов или ребер, называемые следами септ. По характеру следов септ различается три группы нуммулитов: а) с радиальными следами септ, б) с меандровидными, в) с сетчатыми (рис. 733). По форме камер выделяются три основные группы нуммулитов: а) с оперкулиноидными камерами, б) с ассилиноидными, в) с нуммулитоидными (рис. 734). У некоторых нуммулитов имеется промежуточный скелет в виде столбиков, оканчивающихся на поверхности бугорками-гранулами, располагающимися беспорядочно или спирально (табл. XIII, фиг. 1, 2). Многочисленные виды. Широко распространен в палеогене (особенно в эоцене) юга СССР; в. мел - олигоцен от Пиренейского полуострова и Англии до Новой Зеландии; палеоген Центр. Америки. Подроды: (?) Operculinoides Hanzawa, 1935 и (?) Ranikothalia Gaudri, 1944, в СССР неизвестны.
* (Родовое название Camerina имеет приоритет перед Nummulites и другими названиями. Однако Международная комиссия зоологической номенклатуры в 1945 г. постановила сохранить название Nummulites как nomen conservandum из-за того, что в геологической литературе укоренились такие термины, как нуммулитовая система, нуммулитовый известняк и т. д.)
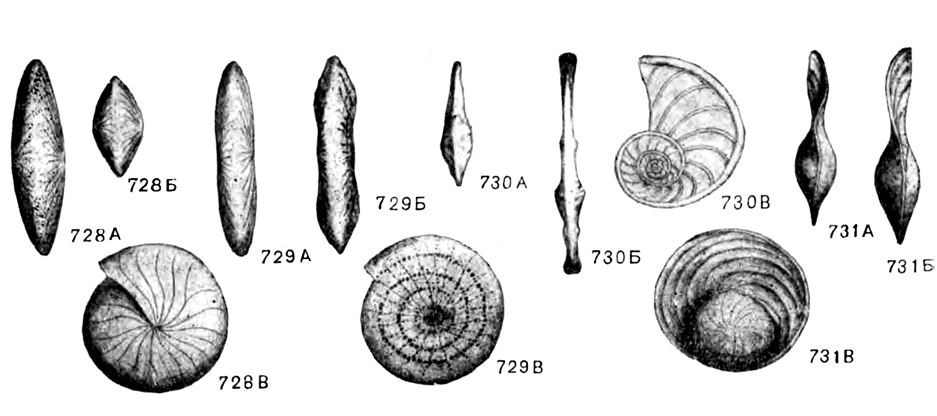
Рис. 728-731. Схематический внешний вид основных родов подсем. Nummulitinae. 728 А-В. Nummulites Lamarck. 729 А-В. Assilina Orbigny. 730 А-В. Operculina Orbigny. 731 А-В. Operculinella Yabe (Немков, 1955)
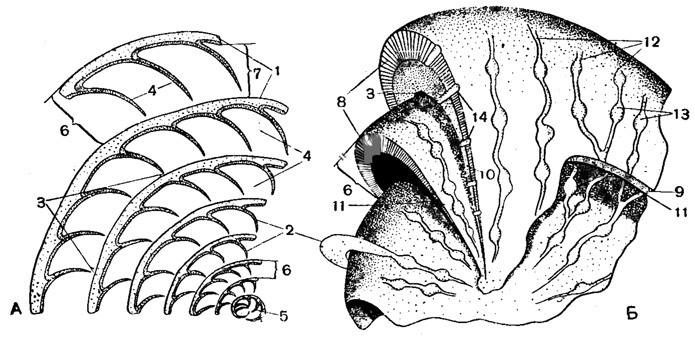
Рис. 732. Схема строения раковины Nummulites: А - экваториальное сечение, Б - вид сбоку; 1 - спиральная полоса; 2 - септы; 3 - устье; 4 - камеры; 5 - начальная камера; 6 - высота оборота или шаг спирали; 7 - высота канала; 8 - спиральный валик; 9 - боковые стенки оборотов; 10 - боковые продолжения септ; 11 - боковые продолжения камер; 12 - следы септ; 13 - гранулы; 14 - столбики гранул (Немков, 1955)
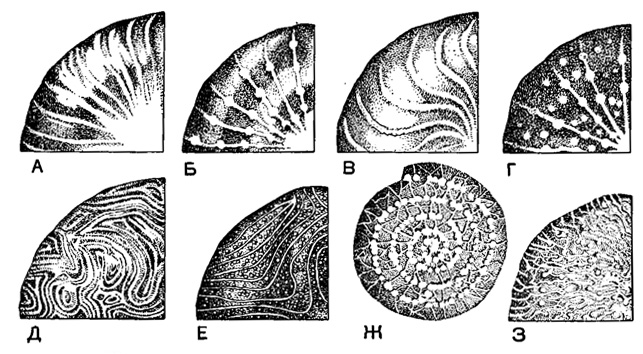
Рис. 733. Типы следов септ (перегородочных ребрышек) у нуммулитов (схема): А-Г - радиальный; Д, Е - меандровый; Ж, З - сетчатый (Немков, 1955)
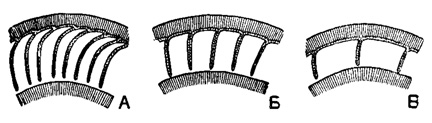
Рис. 734. Различные типы строения камер у нуммулитов: А - оперкулиноидный тип; Б - ассилиноидный; В - нуммулитоидный (Немков, 1955)
Operculinella Yabe, 1918. Тип рода - AmphisteginacutningiiCarpenter, 1859; современный, Индо-Тихоокеанская обл. Раковина дисковидная, вздутая в умбональной части. На ранней стадии роста обороты инволютные, септы радиальные, камеры четырехугольные, слабо серповидные, на поздней стадии - обороты эволютные, септы сильно изогнутые, камеры очень высокие, резко серповидные (рис. 726, Г, З, рис. 731). Очень малочисленные виды. Эоцен Крыма и С. Америки.
Assilina Orbigny, 1826. Тип рода - Nummulites spira Roissy, 1805; ср. эоцен, Франция. Раковина на ранней стадии роста чечевицеобразная с инволютными оборотами и эволютными полостями камер, на поздней стадии - плоская дисковидная, иногда вогнутая в центре, обычно эволютная (редко с инволютными оборотами). Септы радиальные, камеры прямоугольные. У эволютных форм - гранулы, у инволютных - пустулы (бугорки с канальцами в центре) (рис. 726, Б, Е; 729, табл. XIII, фиг. 4). Видов немного. Широко распространен. Эоцен юга СССР; палеоцен - эоцен Ю. Европы, С. Африки, Индии, Индонезии.
Operculina Orbigny, 1826. Тип рода - Lenticulitescomplanata Defrance, 1822; ср. эоцен, Франция. Раковина плоская, с малым количеством быстро возрастающих оборотов, разделенных аркообразными септами на высокие серповидные камеры. Все обороты хорошо видны на поверхности, так как раковина полностью эволютная (редко на ранней стадии роста обороты инволютны). Иногда развиты гранулы на следах септ (рис. 726, В, Ж, 739, 735). Немногочисленные виды. В. мел - ныне. Эоцен юга СССР; третичные отложения Европы, Индии, Индонезии, Японии, Австралии, Америки. Подрод Nummulitoides Abrard, 1956.
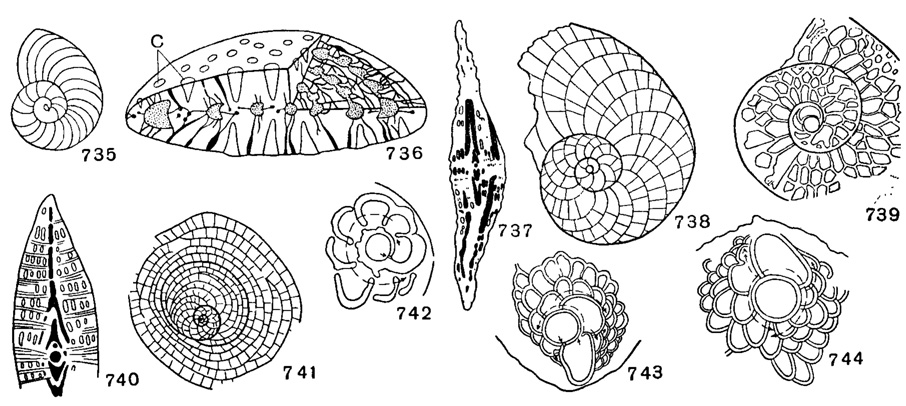
Рис. 735-744. Nummulitidae и Miogypsinidae: 735. Operculina complanata Defrance; × 7, миоцен, З. Европа (Glaessner, 1948). 736. Pellatispira; реконструкция, с - столбики (Sigal, 1952). 737. Arnaudiella grossouvrei H. Douville; (Sigal, 1952). 738. Heterostegina complanata sculpiurata Papp et Kupper; × 17, Ангола (Papp, 1956). 739. Grzibowskia multifida Bieda; × 18, Карпаты (Bieda, 1950). 740. Spiroclypeus; схема (Sigal, 1952). 741. Cycloclypeus posteidae Tan Sin Hok; форма В, × 19 (Sigal, 1952). 742. Miogypsina borneensis Tan Sin Hok; с односпиральным непионтом (Glaessner, 1948). 743. M. bifida Tan Sin Hok; с четырехспиральным непионтом (Glaessner, 1948). 744. M. indonesiensis Tan Sin Hok; с четырехспиральным непионтом (Glaessner, 1948)
Полость спирального канала раковины разделена септами на простые камеры, спиральный валик отсутствует или развит очень слабо. Сильно развиты гранулы. Три рода.
Miscellanea Рfеndеr, 1934. Тип рода - Nummulites miscella Archiac et Haime, 1853; палеоцен, Индия. Раковина чечевицеобразная, как у Nummulites, но с крупными гранулами в центре и более мелкими вдоль следов септ. Обороты и полости камер инволютные. Септы изогнутые, камеры высокие, слабо серповидные. Спиральный валик отсутствует, на его месте развит округлый вторичный спиральный канал (табл. XIII, фиг. 3). Несколько видов. Палеоцен - эоцен юга СССР; в. мел - эоцен Ю. Европы, Индии, Центр. Америки.
Вне СССР: (?) Sulcoperculina Thalmann, 1938; (?) Laffiteina Marie, 1945.
Полость спирального канала раковины разделена септами на простые камеры; спиральный валик развит очень сильно, так что ширина спиральной пластинки равна или превышает высоту камер соответствующего оборота. Сильно развиты гранулы и система каналов. Три рода.
Pellatispira Вoussac, 1906. Тип рода - Р. douvillei Boussac, 1906; приабонский ярус, в. эоцен, Италия. Раковина чечевицеобразная или плоская, дисковидная с крупными гранулами расположенными спирально. Обороты инволютные, полости камер эволютные. Спиральный валик сильно развит, спиральная пластинка толстая. Многочисленные спиральные, радиальные и межсептальные канальцы (рис. 736). Видов мало. Эоцен Закавказья, Индо-Тихоокеанской обл. Подроды: Vacuolispira Tan Sin Hok, 1936; Biplanispira Umbgrove, 1937 (= Heterospira Umbgrove, 1936).
Siderolites Lamarck, 1801 (Siderolina Defrance, 1824). Тип рода - Siderolites calcitrapoides Lamarck, 1801; Маастрихт, Голландия. Раковина чечевицеообразная или с радиальными лучами, сильно гранулированная. Обороты инволютные, полости камер эволютные; на ранней стадии - трохоидное навивание, позже - спирально-плоскостное. Спиральный валик сильно развит, спиральная пластинка толстая. Многочисленные спиральные, радиальные, межсептальные и латеральные канальцы (рис. 724, табл. XIII, фиг. 6, 7). Мало видов. В. мел - ныне. Маастрихт Закавказья и Средиземноморской области.
(?) Arnaudiella Н. Douville, 1907. Тип рода - A. grossouvrei Н. Douville, 1907; кампан, Франция. Раковина дисковидная, сильно гранулированная, спиральный валик хорошо развит. На ранней стадии роста обороты и полости камер инволютные, камеры простые; на поздней - обороты полуинволютные, полости камер эволютные, развиваются вторичные камерки (рис. 737). Монотипный род. Сенон Кавказа и кампан Франции.
Обороты спиральные или концентрические. Камеры в начальной стадии роста простые, затем они подразделяются на более мелкие вторичные камеры. Спиральный валик развит только в начальных оборотах. Четыре рода.
Heterostegina Orbigny, 1826. Тип рода - H. depressa Orbigny, 1826; современный, побережье о-ва Св. Елены. Раковина уплощенная дисковидная, похожая на оперкулину. На ранних стадиях роста инволютная, на поздних - эволютная. Камеры в начальных оборотах оперкулиноидные - высокие и серповидные, а в последующих подразделены на вторичные прямоугольные камерки. Устье - ряд округлых отверстий; между камерками одной и той же камеры нет связи (рис. 738). Видов мало. Эоцен - ныне. Эоцен Закавказья, Средиземноморской и Тихоокеанской областей.
Grzibowskia Bieda, 1950. Тип рода - Grz. multifida Bieda, 1950; в. эоцен, Карпаты. Раковина чечевицеобразная, выпуклая в умбональной части с инволютными оборотами. Камеры простые в начальных оборотах, в последующих подразделены на многочисленные вторичные камерки различной формы (рис. 730). Видов мало. В. эоцен Карпат и Армении.
Spiroclypeus Н. Douville, 1905. Тип рода - S. orbitoideus Н. Douville, 1905; в. олигоцен, о. Борнео. Раковина инволютная, напоминающая в экваториальном сечении гетеростегину, в поперечном - лепидоциклину. Камеры оперкулиноидные, подразделяются на вторичные камерки почти непосредственно за мегасферой. Развиты боковые камеры и столбики, как у орбитоидов (рис. 740). Мало видов. В СССР не встречен. В. эоцен - н. миоцен Индо-Тихо океанской и Средиземноморской областей и Венецуэлы.
Cycloclypeus Carpenter, 1856. Тип рода - С. carpenteri Brady, 1884; современный, у о-ва Борнео. Раковина плоская, дисковидная, сильно похожая на гетеростегину в начальных оборотах, но в последующих камеры становятся концентрическими, подразделенными на вторичные прямоугольные камерки (рис. 741). Видов мало. Олигоцен - ныне. Индо-Тихоокеанская обл. Подроды; Heteroclypeus Schubert, 1906; Radiocycloclypeus Tan Sin Hok, 1932; Katacycloclypeus Tan Sin Hok, 1932.
Раковина чечевицеобразная, более или менее симметричная, от треугольного до округлого очертания. Эмбрион у форм А двухкамерный. Ювенариум форм А спирально-плоскостной или слегка трохо-спиральный, центральный, субцентральный, субпериферический или периферический; у форм В ювенариум спиральный. Непионт одно-, двух- или четырехспиральный. Столоны: радиальные (дистальные и проксимальные), диагональные и проксимальные, диагональные и кольцевые. Экваториальные камеры: дугообразные, ромбовидные, лопатообразные и гексагональные; боковые камеры обычно хорошо развиты. Система канальцев: интрасептальные, радиальные, спиральная; хорды нет. По положению ювенариума, а также по наличию или отсутствию боковых камер семейство делится на роды и подроды. Олигоцен - миоцен. Два рода.
Miogypsina Sacco, 1893 (Ftabelliporus Dervieux, 1900; Lepidosemicyclina Rutten, 1911). Тип рода - Nummulites globulina Michelotti, 1841; H. миоцен, Италия. Эмбрион двухкамерный, непионт периферический; ранние камеры спиральные, позднее рост происходит только в одном секторе, вследствие чего получается раковина треугольного очертания; имеются спиральный и интрасептальные канальцы; экваториальные камеры ромбовидные или гексагональные с четырьмя столонами; имеются боковые камеры. Столбики оканчиваются на поверхности раковины в виде сосочков (рис. 742-744). Видов много. Олигоцен - миоцен Европы, Америки, Индии, С. Африки. Подроды: Miogypsinopsis Hanzawa, 1940; Miolepidocyclina Silvestri, 1907 (Heterosteginoides Cushman, 1919).
Вне СССР: род Miogypsinoides Yabe et Hanzawa, 1928 с подродами Conomiogypsinoides Tan Sin Hok, 1936 и Miogypsinella Hanzawa, 1940.
[nom. transl. Schubert, 1920 (ex Orbitoidinae Prever, 1904)]
Раковина дисковидная или чечевицеобразная, звездчатая или седловидная, выпуклая или вогнутая с двух сторон. Поверхность раковины гладкая, сетчатая, гранулированная или лучистая. Начальные камеры форм В часто спиральные, а у форм А эмбрион, иногда спиральный, много- или двухкамерный; экваториальные камеры дугообразные или гексагональные; обычно присутствуют боковые камеры, правильно или неправильно расположенные вокруг столбиков. В. мел - палеоцен? Подсемейства: Omphalocyclininae, Orbitoidinae, Pseudorbitoidinae и Lepidorbitoidinae.
Эмбрион четырехкамерный; у некоторых форм имеются боковые камеры, которые трудно отличимы от экваториальных. Три рода.
Omphalocyclus Bronn, 1852. Тип рода - Orbulites macroporus Lamarck, 1801; Маастрихт, Голландия. Раковина дискоидальная, двояковогнутая; у форм В эмбриональные камеры спирально-плоскостные, а у форм А эмбрион четырехкамерный. В центре раковины один или два слоя экваториальных камер; в периферической части между экваториальными камерами вклиниваются боковые; наружные стенки экваториальных камер, расположенные концентрически, почти прямые или слегка изогнутые; периферические камеры открываются наружу краевыми порами (рис. 745). Видов мало. Маастрихт юга СССР, З. Европы, Индии, Кубы.
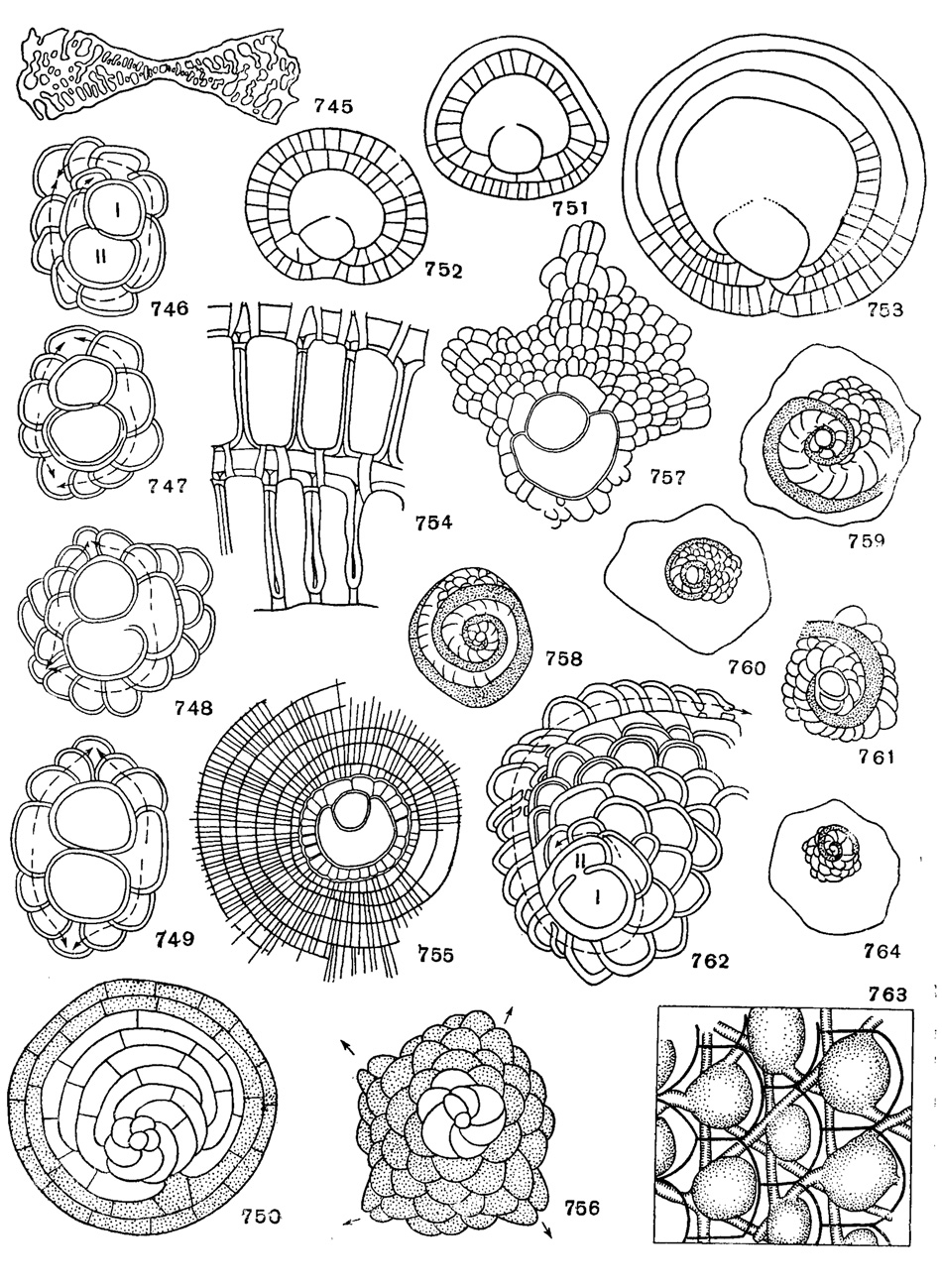
Рис. 745-764. Сем. Orbitoididae, Discocyclinidae, Lepidocyclinidae: 745. Omphalocyclus macroporus (Lamarck); вертикальный разрез, × 8, Маастрихт, Франция (Cushman, 1948). 746. Orbitocyclinoides scheticki Brönnimann; с трехспиральным непионтом, × 90, верхний мел, Иран (Bronnimann, 1944). 747. Оrbitocyclinoides schenckl Bronnimann; с асимметричным четырехспиральным непионтом, × 90, верхний мел, Иран (Brönnimann, 1944). 748. Orbitocyclinoides schencki Brönnimann, с асимметричным четырехспиральным непионтом, × 90, верхний мел, Иран (Brönnimann, 1944). 749. Orbltocyclinoides schencki Brönnimann; с симметричным четырехспиральиым непионтом, × 90, верхний мел, Иран (Brönnimann, 1944). 750. Discocyclina раругасеа (Boube); форма В, в центре - начальная камера, незаштрихованные камеры - непионические, камеры с точками - неанические (Brönnimann, 1946). 751-752. Discocyclina angusiae van der Weijden; расположение периэмбриональных камер (Renz u. Küpper, 1946). 753. Discocyclina aff. varians (Kaufmann) (Renz u. Küpper, 1946). 754. Discocyclina striati-emanuelis Brönnimann; разрез экваториальных камер по плоскости симметрии, в стенках камер видны 'темные линии' и столоны, × 300 (приблизительно); эоцен, Крессенберг. 755. Aciinocyclina radians (Archiac); × 40, эоцен, Верона (Brönnimann, 1946). 756. Asterocyclina stellaris ((Brunner); форма В, четырехлучевой экземпляр в центре - начальная камера; незаштрихованные камеры - непионические; камеры с точечками - неанические (Brönnimann, 1946). 757. Asterocyclina stellaris (Brunner); форма A; × 40, В. эоцен Марокко (Brönnimann, 1946). 758. Helicostegina gyralis Barker et Grimsdale; схематическое строение спиральных камер (Brönnimann, 1944). 759. Helicostegina dimorpha Barker et Grimsdale; схематическое строение спиральных камер (Brönnimann, 1944). 760 Helicolepidina paucispira Barker et Grimsdale; схематическое строение спиральных камер (Brönnimann, 1944). 761. Helicolepidina polygyralis Barker; схематическое строение спиральных камер (Bronnimann, 1944). 762. Helicolepidina spiralis Tobler; двухспиральный непионт с главной спиралью, начиная с центра, × 80, Венесуэла (Brönnimann, 1944). 763. Lepidocyclina (Eulepidina) favosa Cushman; жирные линии - очертания камер (Glaessner, 1948). 764. Eulinderina guyabalensis Barker et Grimsdale; схематическое строение спиральных камер (Brönnimann, 1944)
Вне СССР: Torreina Palmer, 1934 и Monolepidorbis Astre, 1927.
Эмбрион четырех- или двухкамерный (редко больше), окруженный толстой стенкой; экваториальные камеры дугообразные, с четырьмя столонами; имеются и кольцевые столоны; боковые камеры отличаются от экваториальных Три рода.
Orbitoides Orbigny, 1847 (Orbitella Н. Douville, 1915). Тип рода - Orbitolites media Archiac, 1835; в. сенон, юг Франции. Раковина чечевицеобразная, симметричная или асимметричная, орнаментирована червеобразными столбиками или радиальными ребрами. Эмбрион, вначале состоящий из 2-4 камер, позже становится двухкамерным вследствие атрофии двух из них, тогда большая камера объемлет меньшую; боковые камеры низкие; экваториальные камеры с выпуклой наружной стенкой и сходящимися ковнутри боковыми стенками; радиальный диаметр их короче поперечного (табл. XIII, фиг. 6). Несколько видов. В. сенон юга СССР, З. Европы, Индии, Америки.
Simplorbites de Gregorio, 1882. Тип рода - S. cupulimis de Gregorio, 1882; в. мел, Сицилия. В отличие от Orbitoides большая эмбриональная камера (более 2 мм), яйцевидной формы, окруженная толстой пористой стенкой и подразделенная на камерки неправильного очертания. Видов мало, встречаются совместно с Orbitoides.
Clypeorbis Н. Douville, 1915. Тип рода - Orbitoides mamillata Schlumberger, 1902; Маастрихт, Франция. Раковина асимметричная, чечевицеобразная или низкоконическая, с закругленной верхушкой; эмбриональный аппарат состоит из сферической начальной камеры, под которой развиты 3-4 камеры, образующие розетку; эмбриональные камеры завиты в коническую спираль (табл. XIII, фиг. 7). Видов мало. Маастрихт Европы.
Раковина чечевицеобразная, поверхность гранулированная. Ювенариум у форм В спиральный, а у форм А эмбрион двухкамерный; непионт односпиральный, неправильно завитый; экваториальные камеры неправильные, полигональные или гексагональные, у некоторых форм расположенные в один слой около центра и в несколько слоев близ периферии. Два рода. В. мел.
Вне СССР: Pseudorbitoides Н. Douville, 1922 и Vaughanina Palmer 1934.
Эмбрион у форм А двухкамерный, маленький, редко изолепидиноидного типа, обычно нефролепидиноидного или переходного между ними. Экваториальные камеры лопатообразные, гексагональные или дугообразные (в центре раковины). Четыре рода.
Lepidorbitoides Silvestri, 1907. Тип рода - Orbitoides socialis Leymerie, 1851; Маастрихт, Франция. Раковина чечевицеобразная, округлого очертания, поверхность гранулированная, иногда лучистая. Эмбрион двухкамерный: дейтероконх слегка почковидный и охватывает частично субсферический протоконх (нефроэмбрион); непионт четырехспиральный. Экваториальные камеры, слегка вытянутые в радиальном направлении, в центре раковины дугообразные, в периферической же части лопатообразные; стенка боковых камер тонкая (табл. XIII, фиг. 5). Видов мало. Маастрихт юга СССР, З. Европы, Индии; мел Мексики, Луизианы, Миссисипи, Флориды, Кубы. Маастрихтские подроды Orbitocyclina Vaughan, 1929 и Orbitocyclinoides Bronnimann, 1944 (рис. 746-749), в СССР не встречены.
Вне СССР: Cryptasterorbis Rutten, 1935; Asterorbis Vaughan et Cole, 1932; Hellenocyclina Reichel, 1949.
Раковина дисковидная, округлого или звездчатого очертания. Поверхность раковины орнаментирована гранулами, иногда и лучами. У форм В в экваториальном сечении различимы: протоконх, спиральный непионт и концентрически расположенные неанические камеры; нуклеоконх у форм А состоит из сферического протоконха, полностью или частично объемлемого дейтероконхом. У некоторых (нефролепидиноидных) форм можно различить три типа взаимоотношения между эмбрионом и периэмбриональными камерами: а) эмбрион расположен между двумя кольцами периэмбриональных камер; б) эмбрион вклинивается между камерами первого кольца периэмбриональных камер; в) первое кольцо периэмбриональных камер частично прерывается эмбрионом. Экваториальные камеры, расположенные в концентрические круги, дугообразные или кольцевые; последние подразделены на прямоугольные или слегка гексагональные камерки. Развиты канальцы: интрасептальные, радиальные и кольцевые. Радиальные столоны (два или один) проксимальные и дистальные, в зависимости от того, чередуются между собой камеры или нет; в периэмбриональных камерах, исключая вспомогательные, по одному проксимальному столону; по одному столону отмечено и между дейтероконхом и периэмбриальными камерами; между дейтероконхом и протоконхом один или иногда и несколько столонов. Палеоцен - эоцен. Подсемейства: Discocyclininae и Orbitoclypeinae.
[nom. transl. Bronnimann, 1946 (ex Discocyclinidae Vaughan et Cole, 1940)]
У форм В непионт спиральный, последние камеры непионта подразделены на камерки; неанические камеры расположены циклично, они подразделены на камерки, которые в соседних кольцах чередуются друг с другом; камерки с проксимальными столонами; имеется система интрасептальных и внутристенных канальцев. Шесть родов.
Discocyclina Gümbel, 1870. Тип рода - Orbitolites pratti Michelin, 1840; в. эоцен, юг Франции. Раковина дисковидная, плоская или вздутая, округлого очертания, украшена гранулами. У форм В протоконх сферический, непионт состоит из камер, тангенциально удлиненных; неанические камеры расположены циклично и подразделены на прямоугольные камеры; камеры одного кольца сообщаются с камерами соседних колец (рис. 750-754, табл. XIII, фиг. 8). Видов много. Палеоцен - в. эоцен юга СССР, З. Европы, Мал. Азии, Индии, С. Африки, Америки, Новой Зеландии.
Aktinocyclina Gümbel, 1870. Тип рода - Orbitolites radians Archiac, 1848; в. эоцен, юг Франции. Раковина сходна с раковиной Discocyclina, от которой отличается присутствием на поверхности лучей, состоящих из боковых камер. Кольцевые столоны проксимальные (рис. 755, табл. XIII, фиг. 9). Ср. и в. эоцен Европы и Америки.
В не СССР: Proporocyclina Vaughan et Cole, 1940; Pseudophragmina Н. Douville, 1923; Athecocyclina Vaughan etCole, 1940; Asterophragmina Rao, 1942.
Экваториальные камеры не подразделены на камерки; форма камер, расположенных в общем концентрически, от дугообразной до прямоугольной. Два рода.
Orbitoclypeus Silvestri, 1907 (Exagonocyclina Checchia-Rispoli, 1908). Тип рода - Orbitoclypeus himerensis Silvestri, 1907; эоцен, Сицилия. Раковина дискоидальная, с чередующимися экваториальными камерами. Видов мало. Эоцен З. Европы.
Asterocyclina Gümbel, 1870. Тип рода - Calcarina (?) stellata Archiac, 1846; эоцен, юг Франции. Раковина с многочисленными (5-12) лучами, образованными увеличением числа экваториальных камер; формы камер: дугообразные лопатообразные, прямоугольные (рис. 756-757), Несколько видов. Ср. и в. эоцен юга СССР. З. Европы, М. Азии, Ирана, С. Африки.
Для древних представителей этого семейства характерным является наличие в начальной стадии развития раковины амфистегиноидных камер, которые у более молодых форм постепенно редуцируются; вместе с этим у молодых форм отмечается концентрическое расположение кваториальных камер. Этот признак является критерием для подразделения семейства на два подсемейства. Эоцен - олигоцен. Подсемейства: Helicolepidininae и Lepidocyclininae.
Раковина в начальной стадии роста характеризуется явно выраженным спирально-трохоидным расположением камер; более поздние камеры с умбональной стороны делятся на камерки. Форма камер от дугообразной до шестиугольной, с диагональными, иногда и радиальными столонами; сообщение с боковыми камерами через поры и столоны. Устья спиральных камер суббазальные, с просифонами. Дейтероконх меньше протоконха (подобное явление наблюдается и у некоторых лепидоциклин, а также у Actinosiphon). В родах Helicolepidinoides, Helicolepidina и Helicocyctina отмечается уменьшение количества амфистегиноидных камер, так что у молодых форм все экваториальные камеры походят друг на друга (проявление сходства с Lepidocyclina). Непионт у нижне- и среднеэоценовых форм односпиральный, у верхнеэоценовых - двух- и четырехспиральный. Несколько родов.
Вне СССР: Helicostegina Barker et Grimsdale, 1936; Helicolepidinoides Tan Sin Hok, 1936) (рис. 758-759); Pseudolepidina Barker et Grimsdale, 1937; Helicolepidina Tobler, 1922 (Helicocyclina Tan Sin Hok, 1936) (рис. 760-762); Eoconuloides Cole et Bermudez, 1944; Boreloides Cole et Bermudez, 1947, emend. Reichel, 1949.
Раковина обычно чечевицеобразная, по очертанию округлая, седловидная или звездчатая Поверхность раковины сетчатая, гранулированная, бугорчатая или лучистая. Толстостенный эмбрион состоит из двух камер (редко больше); только у одного рода в центре раковины развиты неясно выраженные амфистегиноидные камеры. Экваториальные камеры у форм подсемейства, в отличие от Helicolepidininae, расположены концентрически; камеры соседних колец чередуются друг с другом; формы камер дугообразные, ромбовидные, полигональные, лопатообразные и гексагональные. Столоны диагональные и радиальные. Система канальцев отсутствует (?). Два рода.
Lepidocyclina Gümbel, 1870 (Isolepidina H. Douville, 1915). Тип рода - Nummulites mantelli Morton, 1833; олигоцен, Алабама. Раковина без амфистегиноидных камер. Нуклеоконх у мегасферических форм нескольких типов: а) изолепидиноидный, б) нефролепидиноидный, в) триблиолепидиноидный, г) евлепидиноидный, д) полилепидиноидный, состоящий из восьми крупных камер, две из которых (иногда больше) приблизительно равного размера, е) плиолепидиноидный, состоящий из двух равных камер, рядом с которыми расположены мелкие, ж) мультилепидиноидный с большой центральной камерой, окруженной кольцом, состоящим из 5-10 камер. Непионт одно-, двух-, четырех- и многоспиральный (рис. 763,табл. XIII, фиг. 10). Много видов. Ср. эоцен - н. миоцен Европы, Индии, Америки. Подроды: Polylepidina Vaughan, 1942; Pliolepidina Н. Douville, 1915 emend. Vaughan et Cole, 1941; Nephrolepidina H. Douville, 1911; EulepidinaH. Douville, 1911; MultilepidinaHanzawa, 1932; Lepidocyclina Gumbel, 1870.
Вне СССР: Actinosiphon Vaughan, 1929; Triplalepidina Vaughan et Cole, 1938; Eulinderina Barker et Grimsdale, 1936 (Eolepidina Tan, 1939) (рис. 764).
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'