
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Строение протоплазматического тела современных радиолярий
Протоплазма. Существование у радиолярий характерной для них центральной капсулы приводит к тому, что у громадного большинства форм резко различается протоплазма, заключенная в центральную капсулу (внутрикапсулярная протоплазма, интракапсулярная) и лежащая вне ее (внекапсулярная, экстракапсулярная). Внутрикапсулярная протоплазма часто гомогенного строения, плотная; у феодарий и низших спумеллярий в ней имеются вакуоли. Ядерный аппарат радиолярий всегда помещается внутри центральной капсулы.
У большинства радиолярий во внутрикапсулярной протоплазме сосредоточены запасные питательные вещества (жиры, белковые гранулы), заключены кристаллические продукты обмена веществ, а у некоторых спумеллярий кристаллографически устанавливается присутствие кристаллов целестина. У феодарий, однако, во внутрикапсулярной протоплазме запасных питательных веществ обнаружить не удалось (Решетняк, 1955) в силу сосредоточения их, как будет видно ниже, в пределах так называемого феодия. Наконец, у многих радиолярий внутри центральной капсулы заключены зерна пигмента, отчего она окрашивается в яркие красные, синие, зеленые и другие цвета.
Во внекапсулярной протоплазме, в свою очередь, можно различить три слоя. Непосредственно к центральной капсуле прилегает слой зернистой протоплазмы, обычно сравнительно тонкий - саркоматрикс, но у феодарий, особенно среди высших их представителей, достигающий значительной толщины. Поверх саркоматрикса располагается мощный слой студенистой протоплазмы, или калиммы; калимма у акантарий однородная, прозрачная. В других отрядах радиолярий калимма имеет грубовакублистую структуру. Поверхность калиммы покрыта третьим тончайшим сетчатым слоем - саркодиктиумом (рис. 3).
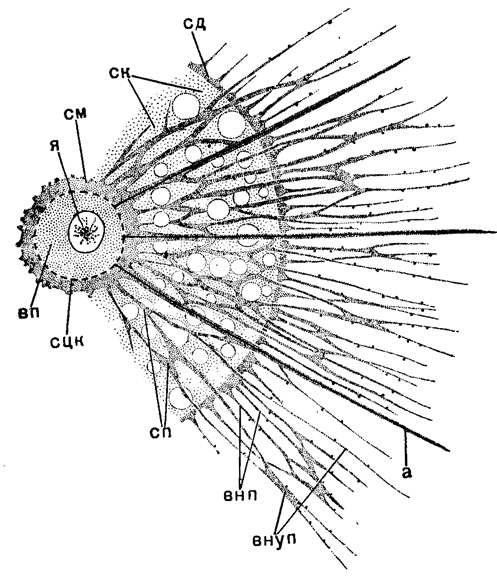
Рис. 3. Схема расслоения протоплазмы радиолярий: сд - саркодиктиум; ск - студенистая калимма; см - саркоматрикс; я - ядро; вп - внутрикапсулярная протоглазма: сцк - пористая стенка центральной капсулы; сп - саркоплегма (сеть внутрь калиммарных псевдоподий в толще калиммы); внп - внекалиммарные псевдоподии; внуп - внутрикалиммарные псевдоподии; а - аксоподии (орнг.)
В связи с подобным расслоением внекапсулярной протоплазмы стоит возникновение псевдоподий. Псевдоподии отходят от саркоматрикса, пронизывают толщу калиммы и выходят наружу (внутрикалиммарные псевдоподии). От саркодиктиума также отходят самостоятельно более короткие псевдоподии (внекалиммарные). Обычно эти псевдоподии имеют характер сетчатых филоподий - острых, тонких, разветвленных, анастомозирующих друг с другом. Их сеть, пронизывающая калимму, образует протоплазматическую основу последней (саркоплегма). Кроме таких филлоподий, у Acantharia, Spumellaria и Sticholonchea имеются особые прямые аксоподии, с осевой нитью внутри, берущие начало также от саркоматрикса (это название и означает - слой зарождения псевдоподий). У Sticholonchea осевая нить (вернее - стержень, ибо он плотной консистенции) берет начало от особых трубчатых коротких образований, связанных шарнирами со стенкой центральной капсулы (Догель, 1951). У некоторых спумеллярий (Sphaerellaria) описывается схождение осевых нитей аксоподий внутри ядра в особом тельце - аксопласте, подобно некоторым солнечникам. Аксоподии выступают наружу, и у акантарий в саркодиктиуме вокруг скелетных игл правильным образом располагаются отверстия для аксоподий, придающие многоугольный вид поверхности тела этих радиолярий. Саркодиктиум у акантарий становится более плотным и упругим благодаря проходящей там системе фибриллей (рис. 4).
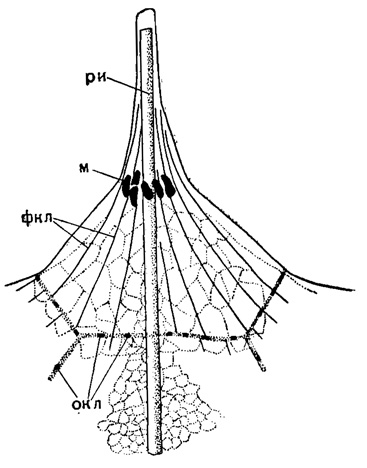
Рис. 4. Acantholithium dicopum Haeckel (Acantharia, Astrolithidae); современный вид из Средиземного моря. Участок калиммы около иглы, ×750. (Schewiakoff, 1926): ри - радиальная игла; м - мионемы; фкл - фибриллы калиммы; окл - отверстия в калимме, через которые выходят наружу аксоподии
Наконец, у низших акантарий (Holacantha) и у некоторых спумеллярий (Porodiscinae с трехлопастным телом) имеются особые флагеллоподии, способные совершать медленные гребные движения (рис. 5).
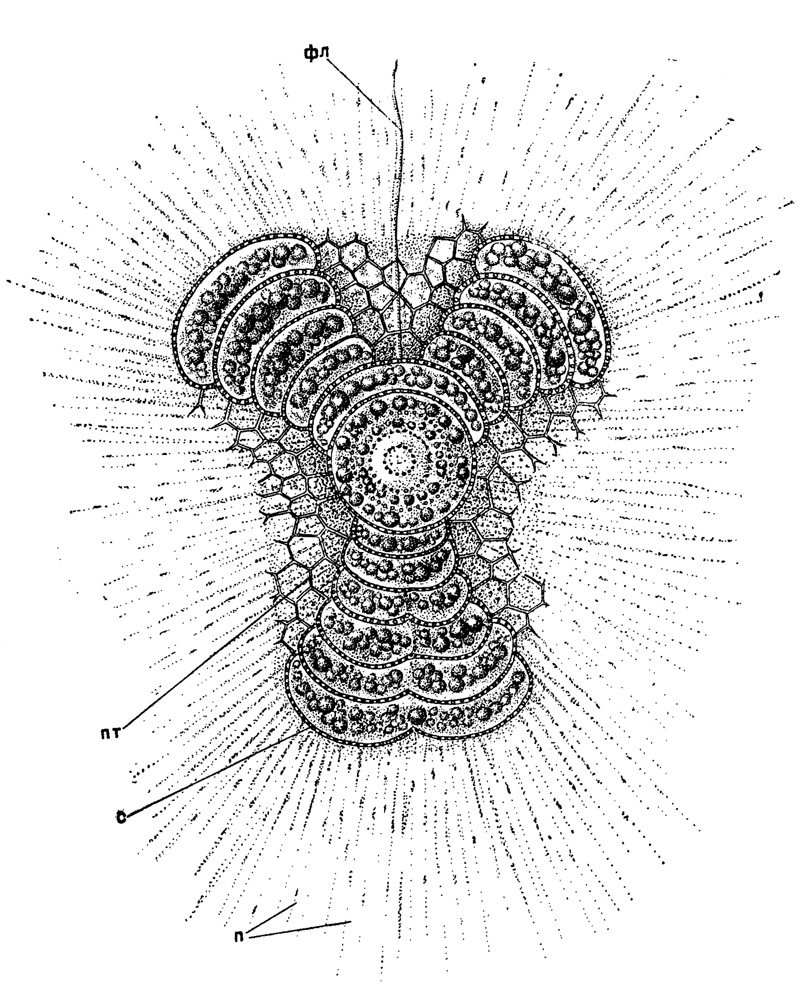
Рис. 5. Euchitonia virchowii Haeckel (Spumellaria, Porodiscidae); живой экземпляр из Средиземного моря, × 350. (Haeckel, 1862): фл - флагеллоподия, пт - ячеи патагия; с - скелет; п - псевдоподии
Основное назначение псевдоподий у радиолярий - захват пищи. Радиолярии плотоядны, захватывают псевдоподиями различные мелкие планктонные организмы, причем есть данные, что подвижные формы парализуются прикосновением к ним псевдоподий. Вокруг захваченной добычи сливается протоплазма из нескольких псевдоподий, и переваривание пищи начинается еще вне тела радиолярии. Далее псевдоподии укорачиваются, добыча втягивается внутрь тела и переваривание заканчивается в саркоматриксе.
Центральная капсула, однако, свойственна вовсе не всем радиоляриям; в отряде акантарий низшие его представители (около 1/3 видов отряда) лишены центральной капсулы. У Низших акантарий не приходится говорить о внутри- и внекапсулярной протоплазме, но калимма развита и у них, и псевдоподии отходят от поверхности внутренней протоплазмы, граничащей с калиммой, т. е. общие отношения и у них могут быть, сведены к общей схеме разделения тела радиолярий на выше разобранные слои.
Скелетные элементы у громадного большинства радиолярий залегают во внекапсулярной протоплазме. Только у акантарий радиальные иглы пронизывают стенку центральной капсулы, к сходятся в центре тела, сочленяясь определенным образом.
Саркоматрикс внекапсулярной плазмы содержит многочисленные включения. Самыми важными из них являются названные уже пищевые частички. У феодарий в области главного отверстия центральной капсулы в саркоматриксе выделяется особое скопление различных включений - феодий. Основным признаком этого особенного органоида тела феодарий является обилие пигментных зерен коричневого и зеленого цвета, придающих характерную окраску этому участку протоплазмы. В области феодия концентрируются жировые капли, зерна гликогена, белковые гранулы; здесь же наблюдаются скопление хондриозом, и тут же лежат особые тельца - феоделлы. Наконец, феодий переполнен пищевыми включениями, остатками панцирей захваченной пищи. В целом феодий выполняет функцию обмена веществ, осуществляя переваривание пищи, видимо, за счет секретирования пищеварительных соков из феоделл и накапливая здесь запасные питательные вещества, а также кристаллические экскреторные зерна.
Среди включений внекапсулярной плазмы особое место занимают зооксантеллы - симбиотические одноклеточные водоросли с хроматофорами желтого цвета. Они распределяются равномерно по всей калимме и саркоматриксу (у акантарий они сосредоточены, наоборот, в центральной капсуле). Зооксантеллы у глубоководных радиолярий, в частности у феодарий, отсутствуют. Зооксантеллы принадлежат или к безжгутиковым Peridinea (род Xatttella) или к Cryptomonadina (род Chrysidella). Число зооксантелл у отдельных видов может быть, весьма различным (от 6-10, 20-40 до 150-300 и более у Acantharia). При делении радиолярий зооксантеллы распределяются между дочерними особями.
Развивающиеся после полового процесса зиготы заражаются, видимо, какими-то стадиями зооксантелл извне. Симбионты извлекают пользу из радиолярий, получая защиту, но при голодании радиолярий они перевариваются; при помещении радиолярий в темноту зооксантеллы выбрасываются из тела радиолярий.
Наконец, здесь же следует упомянуть о широко распространенном среди радиолярий паразитизме в их теле разнообразных Peridinea. В экстракапсулярной протоплазме многих спумеллярий паразитируют около десятка видов рода Merodinium. Внутри центральной капсулы и в ядре обитают представители родов Syndinium и Soletiodinium. Живущих в Acantharia и Sticholonchea видов Amoebophrya Chatton относит в особую группу Coelomastigina близкую к Peridinea.
Мионемы: Особая, свойственная только акантариям система сократительных волокон - мионем - представляет, наряду со скелетом, пример достижения наивысшей сложности организации протозойного тела. Мионемы (миофриски Геккеля, 1887) располагаются звездой около каждой скелетной иглы там, где иглы пронизывают поверхностный слой калиммы. Мионемы - это нитевидные или лентовидные образования с поперечной полосатостью, способные сокращаться. К каждой игле они прикрепляются тонкой фибриллой сухожильного характера, и такие же нити продолжаются от мионем под поверхностью калиммы. Число мионем варьирует от 2-4 у низших акантарий, до 40-60 у высших. Сокращение мионем вызывает натяжение поверхности калиммы, которая растягивается между иглами, увеличивая объем тела радиолярии. Способность акантарий совершать миграции в толще воды (см. раздел "Экология") связана с гидростатической регуляцией при помощи мионем - увеличение объема тела при их сокращении повышает парящую способность тела радиолярии, и она пассивно может подниматься кверху. Наоборот, удлинение мионем ослабляет натяжение калиммы между иглами, объем тела становится меньше и радиолярия станет тонуть. Сокращение мионем удается вызвать искусственно всякими раздражающими воздействиями, в том числе и электрическим током рис. 6).
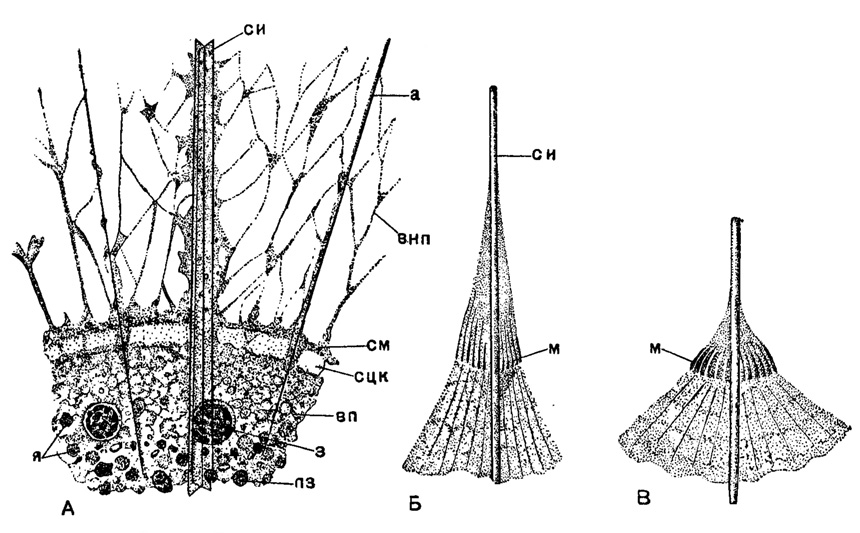
Рис. 6. Детали цитологии живых акантарий из Средиземного моря: А - участок протоплазмы Phyllostaurus cuspidatus Haeckel (Phyllostauridae, × 750); Б - участок калиммы около иглы Acanthomeira pelluclda J. Müller (Acanthometrldae, × 700) с вытянутыми мионемами; В - то же с сократившимися мионемами (Schewiakoff, 1926); си - скелетная игла; а - аксоподия; внп - внутрикалиммарные псевдоподии; см - саркоматрикс: сцк - стенки центральной капсулы; вп - внутрикапсулярная протоплазма; з - зооксантеллы; пз - пигментные зерна; я - ядра; м - мионемы
Возникновение мионем В. Т. Шевяков (1926) связывает в онтогенезе с делением первичного ядра. Часть возникших вторичных "соматических" ядер, по Шевякову, подходит к поверхности калиммы и преобразуется в мионемы. Однако в последнее время высказываются сомнения в возможности преобразования ядер в мионемы, поскольку последние не дают положительной нуклеальной реакции по Фёльгену; картины подобного преобразования, изображенные Шевяковым, признаются неубедительными.
Центральная капсула. Характернейшей особенностью радиолярий следует считать наличие у них центральной капсулы, стенка которой разделяет тело на два основных слоя. Однако сразу же приходится оговориться, что центральная капсула радиолярий не является однородным образованием у всех групп; в этом отношении акантарии отличаются от прочих радиолярий.
У акантарий единственная центральная капсула, как уже указывалось, развита не у всех представителей (имеется у 2/3 видов всего отряда). Химически оболочка центральной капсулы не отличается от остальной протоплазмы и представляет ее уплотненный слой, без пор, залегающий вблизи внутренней границы калиммы.
У прочих радиолярий оболочка центральной капсулы слагается из хитиноидного материала. Холл (Hall) и Дофлейн (Doflein) в составе стенки центральной капсулы указывают тектин (белковое вещество с углеводными компонентами, входящее в состав оболочек цист простейших).
Размеры и форма центральной капсулы весьма варьируют (рис. 7). Форма ее примерно соответствует форметела. У многоосных,шарообразных радиолярий она шарообразна и занимает сравнительно небольшое место в теле радиолярии. Если же тело вытянуто, то вытягивается соответственно и центральная капсула; если на теле имеются выросты, то такие же выросты появляются и на центральной капсуле. У крупной Cytocladus (Spumellaria) центральная капсула образует длинные, дихотомические ветвления, расположенные параллельно ветвям скелета. У колониальных Spumellaria центральная капсула может быть червеобразной. Число центральных капсул у одиночных форм может варьировать от одной до нескольких. Множественность центральных капсул в ряде случаев типична для отдельных групп радиолярий (среди феодарий, например, Aulacaitha нормально имеет 2-4 центральных капсулы, Aulospathis - 8-16, Aulocleptes - 6-8 и т. д.).
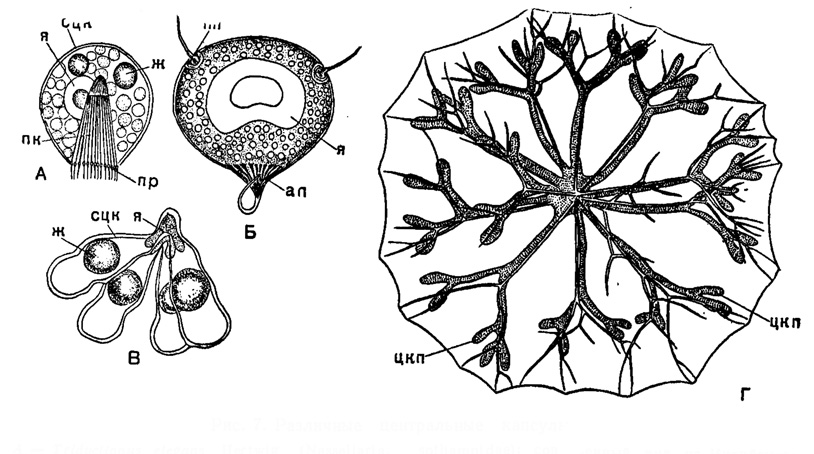
Рис. 7. Различные центральные капсулы: А - Tridyctiopus elegans Hertwig (Nassellarla, Cyrtoldae); простая капсула, × 500 (Hertwig, 1879). Б - Tuscaridium cygneum (Murrey) (Phaeodaria, Tuscarondae); × 50; В - Theopilium cranoides Haecke) Nassellarla, Cyrtoidae); × 500, четырехраздельная капсула, (Hertwig, 1879). Г - Cytocladus tricladus Haecker (Spumellaria, Thalassothamnldae); современный вид на Индийского океана; разветвленная капсула, × 15 (Haecker, 1908): сцк - стенка центральной капсулы; я - ядро; пк - подоконус; пр - поры; ж - капли жира; пп - парапиль; ап - астропиль; цкп - разветвления центральной капсулы
Центральная капсула в исключительно редких случаях может разрастаться, приходя в соприкосновение со скелетным панцирем и вызывая его редукцию (см. Medusettidae из Phaeodaria). При этом поверхность протоплазмы может покрываться пристающими к ней посторонними частицами - панцирями диатомей, силикофлагеллят.
Громадная центральная капсула свойственна среди насселярий сем. Nassellidae - ее покрывает только тонкий слой протоплазмы с отходящими псевдоподиями (рис. 8).
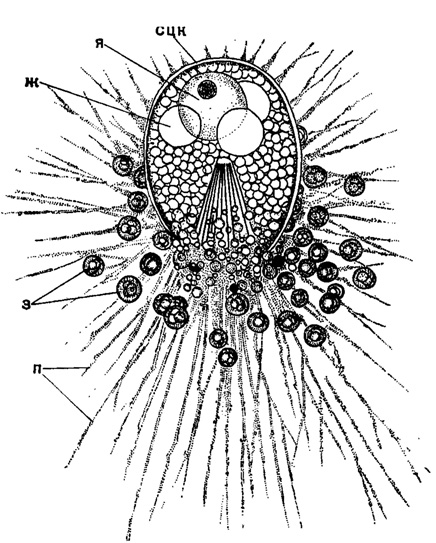
Рис. 8. Cistidium princeps Haeckel (Nasseltaria, Nassellidae); живой экземпляр из Индийского океана. × 350 (Haeckel, 1887): сцк - стенка центральной капсулы; я - ядро; ж - капли жира; з - зооксаителлы; п - псевдоподии
Стенка центральной капсулы пронизана порами, через которые внутрикапсулярная протоплазма сообщается с внекапсулярной. По устройству и по расположению отверстий в стенке мы различаем три типа центральных капсул.
1. У спумеллярий мелкие поры во множестве равномерно пронизывают однослойную стенку центральной капсулы; центральная капсула многоосна.
2. У насселлярий центральная капсула вытянута, становится одноосной, радиально-симметричной; большая часть ее плотной однослойной стенки лишена пор, за исключением порового поля на одном из полюсов капсулы, пронизанного несколькими десятками мельчайших отверстий; от них тончайшие плазматические тяжи уходят внутрь центральной капсулы, образуя конический подоконус.
3. У феодарий центральная капсула становится двустороннесимметричной и приобретает двуслойную толстую стенку; общая шарообразная форма нарушается легким сплющиванием ее полюсов и тем, что стенки ее пронизываются у громадного большинства форм тремя отверстиями (почему их называют еще и Tripylea). Главное из отверстий (астропиль), более крупное, лежит на плоской стороне капсулы и прикрыто крышечкой с радиальной исчерченностью, заканчивающейся трубчатым хоботком. Два других отверстия, побочных (парапили), располагаются на противоположном полюсе, занимают боковое положение и также снабжены короткими трубочками. В редких случаях у феодарий сохраняется одно главное отверстие (сем. Caementellidae) или этих главных в редуцированном состоянии может быть несколько (Globicella, из сем. Medusettidae).
Ядро. В громадном большинстве случаев у радиолярий в покоящемся состоянии в центральной капсуле бывает заключено одно ядро. Исключением являются все акантарии, обладающие многими ядрами.
Радиоляриям свойственны два типа ядер - первичные и вторичные.
Первичное ядро - крупное, полиэнергидное, при размножении. простым делением на двое проделывает типичный митоз, дает громадное число хромозом (например 1000-1500 у Aulacantha, рис. 9, из Phaeodaria). Форма его соответствует контурам центральной капсулы (чаще округлая). Хроматиновые элементы концентрируются в виде тяжа с боковыми отростками, дающего реакцию Фёльгена (Решетняк, 1955).

Рис. 9. Деление ядра у радиолярий: А-В - деление ядра и центральной капсулы при бесполом размножении у Aulacantfia scolymantha Haeckel, × 120; Г - покоящееся ядро A. scolymantha, × 250; Д - профаза ядра A. scolymantha при бесполом размножении, × 250; Е - метафаза при делении ядра А. scolymantha вовремя бесполого размножения, × 250; Ж - анафаза того же деления, × 250; З - хромозомы при бесполом размножении A. scolymantha (большое увеличение), × 700; И - телофаза при делении ядра Thalassicola nucleata Huxley перед образованием гамет, × 1350. Рисунки сделаны с препаратов срезов, окрашенных железным гемотоксилином (А-З - Borgert, 1901; И - 1926)
Вторичные ядра - мелкие, моноэнергидные; их образование путем последовательного митотического многократного деления первичного ядра приводит в конце концов к формированию ядер гамет, участвующих в половом процессе. Число хромозом при делении вторичных ядер не превышает 20 (рис. 9, И).
У акантарий первичное ядро формируется на очень ранней стадии онтогенеза. По мере роста тела и развития скелета это первичное ядро быстро делится многократно, приводя к обычному многоядерному состоянию, в котором акантарии и пребывают длительно. Митозов у акантарий не описано, а сообщаемое Шевяковым (1926) возникновение особых трофических ядер, преобразующихся якобы в мионемы, в последнее время, как указывалось, подвергается сомнениям (Hollande, 1953).
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'