
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Экология современных радиолярий и их географическое распространение (А. А. Стрелков)
Радиолярии - исключительно морские животные, живущие в водах океанической солености (32-38‰) и ведущие пелагический, планктонный образ жизни. Единственным исключением является описанная Шрёдером (Schröder, 1907) в Антарктических водах прикрепляющаяся с помощью стебелька к мшанкам, иглам морских ежей и т. д. Podactinellius sessilis из отрядов Acantharia (рис. 40). Однако Шевяков (1926) с известным основанием высказывает сомнение вообще о принадлежности данной формы к радиоляриям. Радиолярий нет совсем во внутренних морях, Черном и Каспийском, где соленость сильно понижена - до 10-22‰.
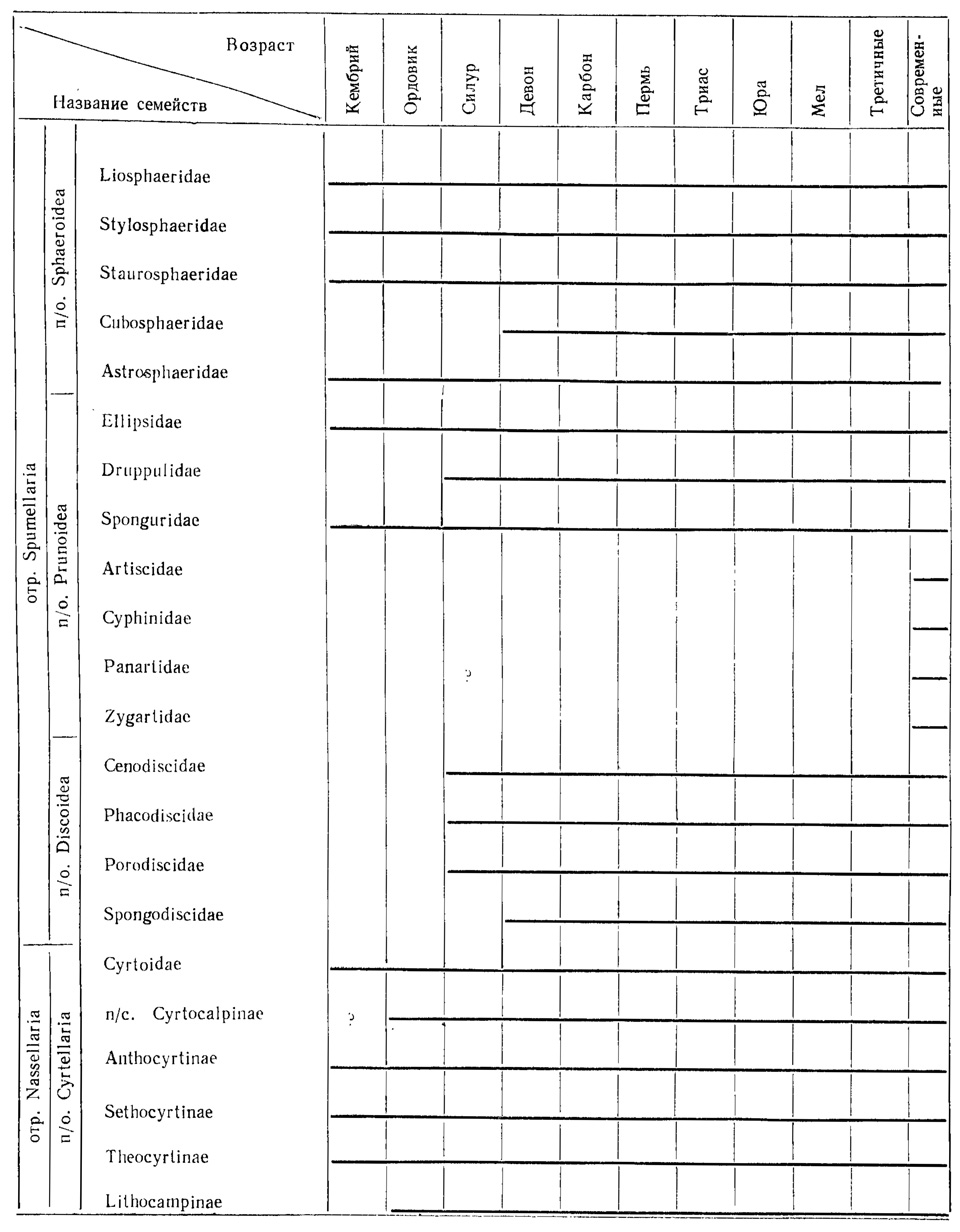
Геологическое распространение некоторых семейств радиолярий
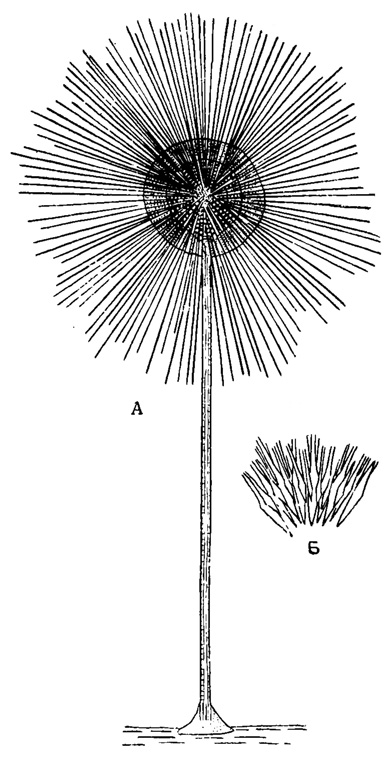
Рис. 40. Podactinelius sessilis Schröder (Astrolophidae); сидячая радиолярия с глубин Антарктических вод: А - общий вид радиолярии, × 75; Б - соединение радиальных игл скелета в центре тела, × 750 (Schröder, 1907)
Подобный образ жизни сочетается с рядом приспособлений, обусловливающих плавучесть радиолярий в толще воды.
Внешний облик радиолярий с торчащими во все стороны иглами скелета и псевдоподиями явно указывает на увеличенную поверхность трения, на усиление парящей способности тела этих животных. Вакуолистость протоплазмы ведет к уменьшению удельного веса тела и также способствует замедлению погружения радиолярий. Особенное же значение приобретают в этом плане имеющиеся в их теле жировые включения, капли которых, выполняя роль поплавков, облегчают плавающее состояние радиолярий.
Радиолярии отличаются способностью населять всю толщу вод океанов, начиная от поверхностных слоев и кончая самыми глубинными зонами. В. Геккер (Haecker, 1908) обнаруживал живых радиолярий на глубинах 4000 и даже 5000 м. В материалах экспедиционного судна "Витязь", собранных в Курило-Камчатской впадине, живые радиолярии были найдены на глубинах более 8000 м (Решетняк, 1955).
Громадное большинство радиолярий встречается в открытых частях морей и океанов. Лишь единичные представители свойственны прибрежным районам и могут считаться неритическими формами.
Из ныне живущих радиолярий представители отр. Acantharia и Sticholonchea свойственны преимущественно поверхностным слоям воды 50-200 м, хотя и встречаются на глубине до 1000 м.
В других же отрядах, наряду с формами сравнительно немногочисленными и свойственными верхним горизонтам воды, имеется большое число видов - обитателей глубинных вод. В целом отр. Spumellaria, Nassellaria и Phaeodaria по их современному распространению могут быть охарактеризованы как довольно глубоководные батипелагические формы.
Геккеру (Haecker, 1908) принадлежит характеристика различных горизонтов толщ вод Мирового океана по преимущественному нахождению в них тех или иных радиолярий. Их именем Геккер и называет соответствующий горизонт:
0-50 м - "слой с коллидами" ("Collidenschicht"): типичные формы Collozoumy Sphaprozoum, Thalassophysa Thalassicolla (из отр. Spumellaria); к ним добавляются Auloscena, Cannosphaera, Sagenoscena (из отр. Phaeodaria).
50-400 м - "слой с челленджеридами" ("Challengeridenschicht"): типичные формы Challengeria, Protocystis; к ним добавляются Conchidium, Ccnchoceros, Conchophacus (все из отр. Phaeodaria).
400-1500 м - "слой с тускароридами" ("Tuscarorenschicht"): преимущественное обитание различных видов сем. Tuscaroridae, Aulacanthidae, Aulosphaeridae, Castanellidae (из отр. Phaeodaria).
1500-5000 м - "слой с фарингеллами" ("Pharyngellenschicht"): типичные формы - Pharyngella, Porcupina, Conchopsis и другие представители отр. Phaeodaria.
Этим горизонтам, по Геккеру, соответствуют вертикальные зоны Ло-Бианко, установленные для Средиземного моря. Ло-Бианко дает выразительные характеристики, основанные на степени освещенности этих зон, а следовательно, и на возможностях ассимиляции, отраженных в распределении диатомовых водорослей:
0-50 м - светлая (освещенная) зона "Phaoplankton";
50-400 м - затененная (полуосвещенная) зона "Knephoplankton";
400-1800 м - темная (малоосвещенная) зона "Skotoplankton";
и 1500-5000 м - ночная (неосвещенная) зона "Nyctoplankton".
Схема Геккера в последующем подверглась уточнению. Обильные материалы, собранные экспедиционным судном "Витязь" в районе Курило-Камчатской впадины (Решетняк, 1955), позволили для этого района Мирового океана прежде всего выделить две экологических категории радиолярий, отсутствовавшие у Геккера - стенобатные формы, приуроченные строго к определенным горизонтам, и эврибатные формы, связанные с несколькими горизонтами или же распределенные во всей толще океанической воды.
Среди обнаруженных в районе Курило-Сахалинской впадины 100 с лишним видов радиолярий, около половины (45%) может быть отнесено на долю стенобатных форм, среди которых выделяются следующие группы:
I. Поверхностные радиолярии, приуроченные к горизонту 0-50 (4% всех стенобатных форм).
II. Субповерхностные радиолярии, приуроченные к горизонту 50-200 м (16% всех стенобатных форм).
III. Среднеглубинные радиолярии, приуроченные к горизонту 200-1000 м (20% всех стенобатных форм).
IV. Батипелагические радиолярии, приуроченные к горизонту 1000-2000 м (32% всех стенобатных форм).
V. Абиссальные радиолярии, приуроченные к горизонту 4000-8000 м (12% всех стенобатных форм).
Ко II и III группам радиолярий следует добавить 16% переходных форм, живущих на глубине 50-1000 м.
Рис. 41 дает наглядное представление о распределении обычных видов, типичных для соответствующего горизонта вод Курило-Камчатской впадины.
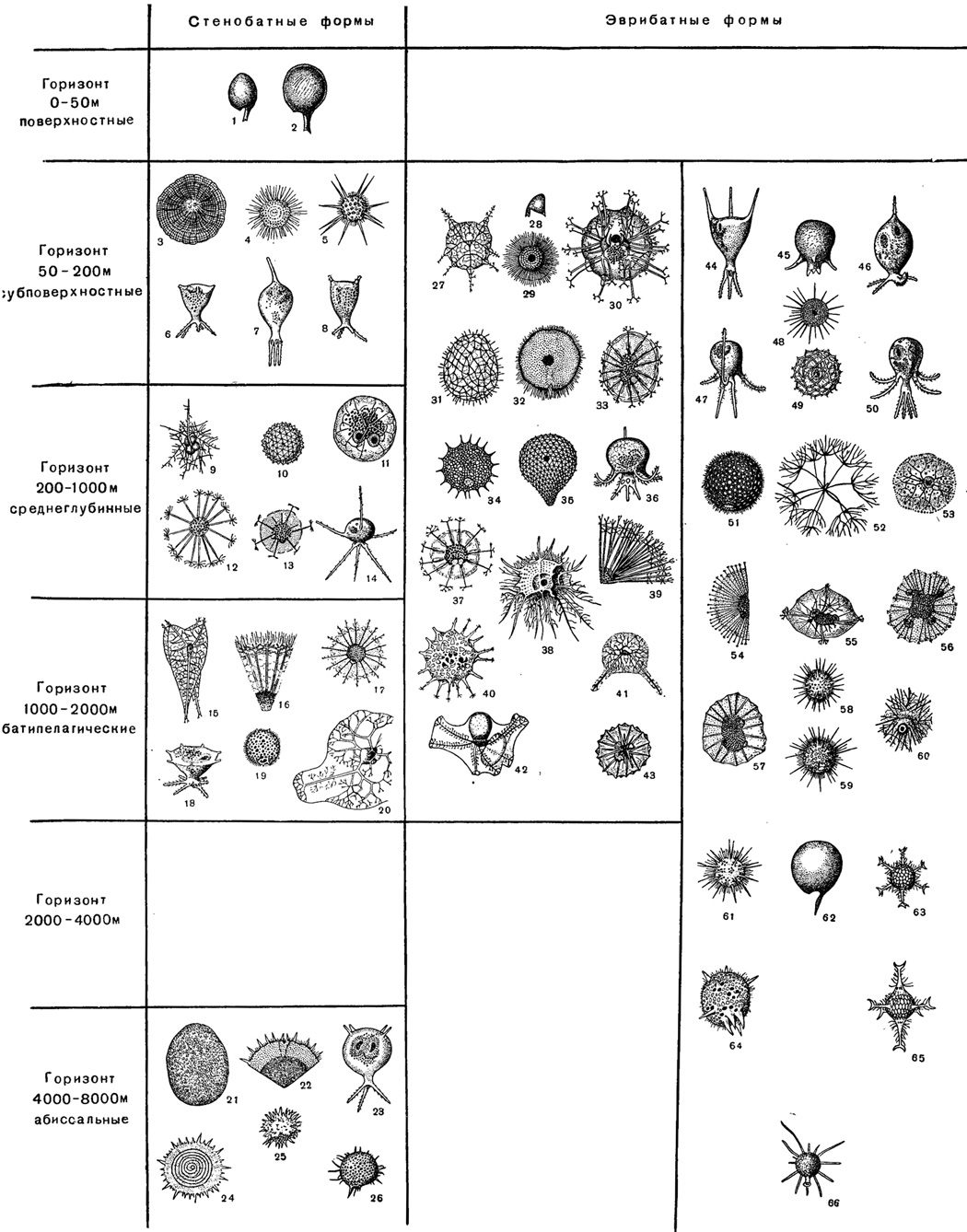
Рис. 41. Основные формы радиолярий, распределенные по глубинным зонам океанических вод Куримо-Камчатской впадины (Из Решетник, 1955). Среди эврибатных радиолярий местоположение той или иной формы соответствует массовому ее нахождению на указанных глубинах. 1 - Protocystis tridens; 2 - Protocystis thomsoni Murray; 3 - Litharachnium epeira Haeckel; 4 - Lithelius spiralis Haeckel; 5 - Heliosoma delicatulum Doglel (отр. Spumellarla); 6 - Tuscarilla wationalis (Borgert) (отр. Phaeodaria); 7 - Tuscarantha lucia Haecker (отр. Phaeodaria); 8 - Tuscarilla scutellum (Haecker) (отр. Phaeodaria); 9 - Thalassothamnus ramosus Haecker (отр. Spumellarla); 10 - Aulosphaera elegantissima Haeckel (отр. Phaeodaria); 11 - Aulographis triangulum Haeckel (отр. Phaeodaria); 12 - Aulographonium candelabrum Haeckel (отр. Phaeodaria); 13 - Auloceros arborescens birameus (Immermann) (отр. Phaeodaria); 14 - Tuscarora bisternaria Murray (отр. Phaeodaria); 15 - Coelographis regina Haeckel (отр. Phaeodaria); 16 - Aulographonium bicorne Haecker (отр. Phaeodaria); 17 - Aulospathis variabilis tetrodon Haecker (отр. Phaeodaria); 18 - Tuscarilla campanella Haecker (отр. Phaeodaria); 19 - Castanissa megastoma Reschetnjak (отр. Phaeodaria); 20 - Coelodiceras spinosum Haecker (отр. Phaeodaria); 21 - Plegmosphaera maxima Haeckel (отр. Spumellaria); 22 - Schlzodiscus stylotrochoides Doglel (отр. Spumellaria); 23 - Tuscarilla ampulla Hacker (отр. Phaeodaria); 24 - Stylotrochus geddesi Haeckel (отр. Spumellaria); 25 - Styptosphaera stupacea Haeckel (отр. Spumellaria); 26 - Castanura primlliva Reschetnjak (отр. Phaeodaria); 27 - Coeloplegma murrayanum Haeckel (отр. Phaeodaria); 28 - Protocystis ornitocephala Reschetnjak (отр. Phaeodaria); 29 - Actissa peinceps Haeckel (отр. Spumellaria); 30 - Coelanthemum auloceroides Haecker (отр. Phaeodaria); 31 - Aulosphaera triodon Haeckel (отр. Phaeodaria); 32 - Schizodiscus spatangus Doglel (отр. Spumellaria); 33 - Auloceros arborescens subelegans Haecker (отр. Phaeodaria); 34 - Castanidium variabile podacontium Reschetnjak (отр. Phaeodaria); 35 - Sagenoarium chuni Borgert (отр. Phaeodaria); 36 - Tuscaretta globosa (Borgert) (отр. Phaeodaria); 37 - Auloceros arborescens dichodendrum (Immermann) (отр. Phaeodaria); 38 - Oroscena regaiis Borgert (отр. Spumellaria); 39 - Aulospathis variabilis triodon Haecker (отр. Pnaeodarila); 40 - Polypyramis fenestrata Reschetnjak (отр. Phaeodaria); 41 - Coelothyrsus cypripedium Haecker (отр. Phacodaria); 42 - Planktonetta atiantica Borgert (отр. Phaeodaria); 43 - Aulacantha scolymantha typica Haecker (отр. Phaeodaria); 44 - Tuscarantha braueri Haecker (отр. Phaeodaria); 45 - Tuscaretta сера Haecker (отр. Phaeodaria); 46 - Tuscaridium cyg neum (Murray) отр. Phaeodaria); 47 - Tuscaretta tubulosa (Murray) (отр. Phaeodaria); 48 - Trochodiscus helioides Cleve (отр. Spumellaria); 49 - Coelacantha dogieli Reschetnjak (отр. Phaeo darla); 50 - Tuscaretta belknapi (Murray) (отр. Phaeodaria); 51 - Casianella bulbospinosa Reschetnjak (отр. Phaeodaria); 52 - Cytdcladus dogieli Reschetnjak (отр. Spumellaria); 53 - Coelodendrum ramoslssimum Haeckel (отр. Phaeodaria); 54 - Aulographis stellatta Haeckel (отр. Phaeodaria); 55 - Aulokteptes ramosus Immermann (orp. Phaeodaria); 56 - Aulokleples flosculus (Haeckel) (отр. Phaeodaria); 57 - Aulographis pandora Haeckel (отр. Phaeodaria); 58 - Haeckeliana irregularis Haecker (отр. Phaeodaria); 59 - Castanidium variabile Borgert (отр. Phaieodarla); 60 - Aulacantha scolymantha bathybia Haecker (отр. Phaeodaria); 61 - Haeckeliana porcellana Murray (отр. Phaeodaria); 62 - Challengeria naresi (Murray) (отр. Phaeodaria); 63 - Circospathis sexfurea trifida Reschetnjak (отр. Phaeodaria); 64 - Haeckeliana megalodonta Reschetnjak (отр. Phaeodaria); 65 - Circospathis sexf urea Haeckel (orp. Phaeodaria); 66 - Porospathis holostoma (Cleve) (отр. Phaeodaria)
Эврибатные радиолярии Курило-Камчатской впадины либо распределены во всей толще воды, начиная с 50 м и до наибольших глубин (до 8000 м и глубже), таких было встречено около 60% всех эврибатных форм; либо они обладают меньшей степенью эврибатности и распространяются вглубь только до 2000 м (рис. 41). Однако и эврибатные радиолярии впадины обнаруживают известное тяготение к определенным горизонтам, встречаясь в одних чаще и в большем количестве, чем в других. Указанная тенденция графически отражена на рис. 41, где отдельные виды изображены около того горизонта, к которому они особенно приурочены.
Отличие приведенной схемы от данных Геккера заключается именно в том, что у названного автора зоны выделялись с применением менее четкого критерия большей или меньшей встречаемости типичных форм в отдельных слоях воды. У Геккера не хватало фактических данных для выделения стенобатных форм. Один и тот же вид нередко фигурировал как типичный для разных горизонтов. Применение стенобатных форм бесспорно сделало характеристику горизонтов более точной.
Наибольшее видовое разнообразие радиолярий в водах Курило-Камчатской впадины приходится на долю среднеглубинных и батипелагических видов (слой 200-2000 м). В этом же горизонте наблюдается и количественное обилие радиолярий, позволяющее говорить о радиоляриевом планктоне в этом слое воды. Многие пробы, добытые планктонными сетями с глубин 200-2000 м в основном содержали массу тел крупных представителей сем. Aulosphaeridae, Aulacaiithidae, Tuscaroridae (из отр. Phaeodaria). Исключительное обилие радиолярий придавало этим пробам вид "лягушечьей икры". К поверхности и к большим глубинам океана видовой состав радиолярий резко обедняется и количество их падает; следует, однако, отметить что на больших глубинах протозойный планктон почти исключительно составлен из радиолярий; других Protozoa там почти нет.
В отличие от данных Геккера, категории по вертикальному распределению радиолярий в районе Курило-Камчатской впадины могут быть характеризованы типичными для них водными массами, установленными для этого района Морошкиным с определенной температурой (t) и соленостью (s).
I. Поверхностные радиолярии приурочены к поверхностной водной массе со средней t° = 2,6° и s = 33,0‰.
II. Субповерхностные радиолярии населяют холодную промежуточную водную массу с t° = 0,3° и s = 33,0‰.
III. Среднеглубинные радиолярии распределены в теплой промежуточной водной массе с t° = 3,5° и s = 34,1‰.
IV. Батипелагические радиолярии живут в глубинной водной массе с t° = 1,7° и s = 34,7‰.
V. Абиссальные радиолярии приурочены к придонной водной массе с t° = 1,5-2,0° и s =34,7‰.
Иначе говоря, среди в основном тепловодных форм вклинивается небольшое число видов радиолярий, предпочитающих холодную водную прослойку (субповерхностные виды, 16% всех стенобатных радиолярий).
Сезонные изменения, и тем более суточные, в распределении радиолярий почти неизвестны.
У глубинных форм их и ожидать трудно; например, стабильные условия температуры в течение года на значительных глубинах не окажут влияния на изменение распределения. Однако в Средиземном море (Шевяков, 1926) спумеллярии и насселлярии в более холодное время года (с октября по май) поднимаются с глубин к поверхности. Они в это время собираются массами в слое 0-50 м, в то время как акантарии, наоборот, из поверхностных слоев перемещаются в зону 50-200 м.
Опреснение воды чувствительно сказывается на распределении радиолярий. После сильных дождей летом акантарии опускаются в Средиземном море на глубины 100-200 м, скопляясь снова у поверхности воды уже через 1-2 дня.
Волнение моря (средней степени) угоняет акантарий с поверхности моря на глубину 5-10 м; при сильном же волнении скопления акантарий наблюдаются на глубине 50-100 м.
Наконец, в Средиземном море можно назвать несколько "зимних" видов Acantharia, появляющихся и размножающихся в значительных количествах в декабре, январе.
Подобными, еще в значительной степени отрывочными, данными пока ограничиваются наши сведения об экологии современных радиолярий.
Основная масса радиолярий связана с теплыми водами, на что ясно указывает даже распространение массовых скоплений скелетов радиолярий в донных отложениях исключительно в тропической области Мирового океана (см. карту, на которой показаны радиоляриевые илы - с содержанием кремнистых панцирей радиолярий более 20% состава осадка).
В имеющейся литературе описано свыше 6000 видов ныне живущих радиолярий. Из них значительная часть имеет циркумтропическое распространение, ограничиваясь обитанием в тропической зоогеографической области Мирового океана.
По мере продвижения в область умеренных вод, как северных (бореальных), так и южных (нотальных), число ныне живущих видов радиолярий резко падает. Для Атлантического океана, например, в отношении представителей отр. Acantharia известно, что в полосе экватора число видов в 10 раз и более превышает таковое в более северных районах (Popofsky, 1904) (рис. 42). Тоже обнаружено и для северной части Тихого океана, где, в результате обработки многочисленных сборов судна "Витязь" в настоящее время определено всего 185 видов радиолярий (Догель и Решетняк, 1955). В северополярных морях число видов падает до весьма небольшого; например, в Карском море известно всего не более 15 видов радиолярий [Бернштейн (Bernstein, 1932), Хмызникова, 1936]. В морях антарктических известно всего несколько десятков видов (Popofsky, 1903-1913; Schröder, 1913).
Рис. 42. Графическое изображение изменения числа видов акантарий в Атлантическом океане. Ширина заштрихованной полосы показывает число видов из расчета 3 вида на 2 мм (Popofsky, 1904)
Однако такая картина распространения может быть безоговорочно отнесена лишь к радиоляриям, населяющим поверхностные слои океанов, т. е. главным образом к представителям отр. Acantharia.
В отношении же глубинных форм грань между зоогеографическими областями Мирового океана в известной степени стирается прежде всего в силу наличия одинаковых гидрологических условий на больших глубинах как в тропических, так и в умеренных, областях. Известная часть глубинных радиолярий может быть названа панбатипелагическими. К таким, несомненно, принадлежит большинство глубоководных феодарий, обнаруженных в водах Курило-Камчатской впадины (Решетняк, 1956).
Среди циркумтропических видов часть, вероятно, будет развенчана дальнейшими исследованиями; установление этой категории видов было связано с материалами, добытыми знаменитыми глубоководными экспедициями "Челледжер"(1872-1876 гг.) и "Вальдивия" (1898-1899 гг.), изучавшими в основном тропические районы Мирового океана.
При обработке материалов экспедиционного судна "Витязь", собранных в с.-з. части Тихого океана, в настоящее время обнаружилось, что из найденных там 100 видов из отр. Phaeodaria более 20% оказались "тропическими" видами (т. е. до сих пор относились к циркумтропическим формам). Обитание таких форм на больших глубинах и в значительных количествах в районе Курило-Камчатской впадины, в Охотском и Беринговом морях (см. карту рис. 43), заставляет развенчать их "тропическую" природу и включить их в группу панбатипелагических видов.
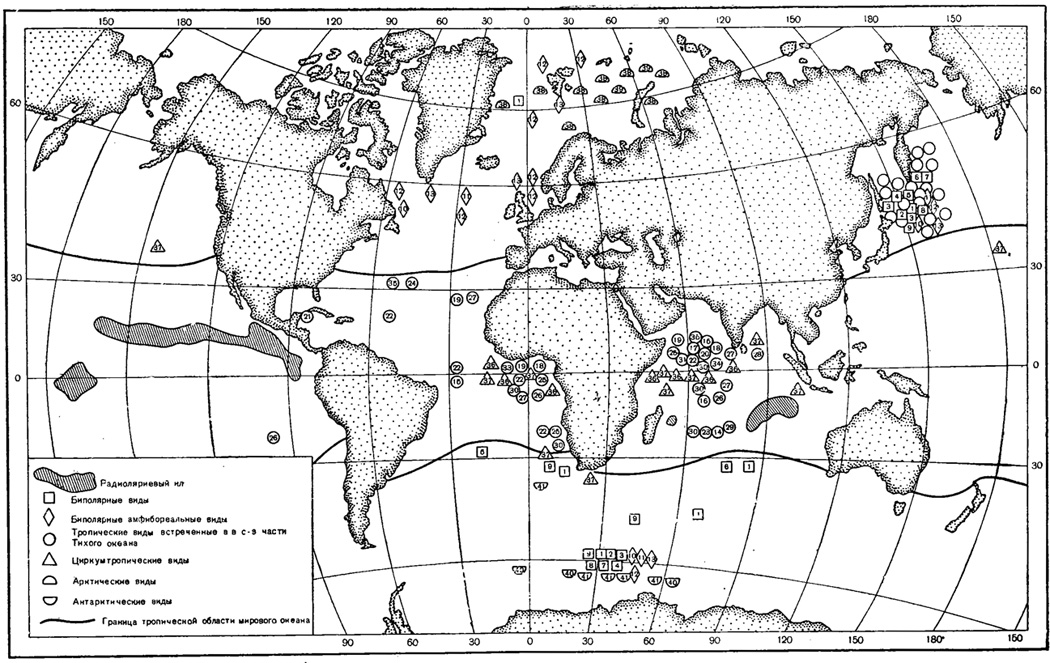
Рис. 43. Карта распределения современных радиолярий в пределах Мирового океана (ориг.): 1 - Attlodendron antarcticum Haeckel; 2 - Auloceros arborescens irregularis Haecker - (отр. Phaeodaria); 3 - Aulographonium antarcticum Haecker (отр. Phaeodaria); 4 - Aulodendron verticillatum Haecker (отр. Phaeodaria); 5 - Auloscena pyramidalis Haeckel (отр. Phaeodaria); 6 - Castanarium favosum Haecker (отр. Phaeodaria); 7 - Caslanidium spinosum Schroder (отр. Phaeodaria); 8 - Coelographis antarctica Haecker (отр. Phaeodaria) 9 - Aulospathis variabilis diodon Haecker (отр. Phaeodaria); 10 - Aulastrum spinosum Borgert (отр. Phaeodaria); 11 - Sagenoscena irmingeriana Borgert (отр. Phaeodaria); 12 - Protocystis Iridens Haeckel (отр. Phaeodaria); 13 - Aulacantha laevissima Haeckel (Phaeodaria); 14 - Aulographis arcuata Haecker (отр. Phaeodaria); 15 - Aulographis pentaslyla Haecker (отр. Phaeodaria); 16 - Auloceros arborescens dichodendrum (Immermann) (отр. Phaeodaria); 17 - Aulographis furcula Haeckel (отр. Phaeodaria); 18 - Aulographis triangulum Haeckel (отр. Phaeodaria); 19 - Aulographonium indicum Haeckee (отр. Phaeodaria); 20 - Aulographonium candelabrum Haeckel (отр. Phaeodaria); 21 - Aulodendron heterocanthum Haecker (отр. Phaeodaria); 22 - Aulospathis variabilis monodon Haecker (отр. Phaeodaria); 23 - Auloscena robustissima Haecker (отр. Phaeodaria); 24 - Sagenoarium verticillatum Haecker (отр. Phaeodaria); 25 - Sagenoarium dicranon Haecker (отр. Phaeodaria); 26 - Castanissa circumvallata Schmidt (отр. Phaeodaria); 27 - Castanea henseni (Borgert) (отр. Phaeodaria); 28 - Cirkocastanea margarita Haecker (отр. Phaeodaria); 29 - Tuscarilla scutellum (Haecker) (отр. Phaeodaria; 30 - Tuscarora bisternaria Murray (отр. Phaeodaria); 31 - Tuscaretta belknapi (Murray) (отр. Phaeodaria); 32 - Tuscaridium cygneum (Murray) (отр. Phaeodaria); 33 - Atlanticella bicornis Haecker (отр. Phaeodaria); 34 - Nationaletta valdivia Borgert (отр. Phaeodaria); 35 - Euphysetta elegans Borgert (отр. Phaeodaria); 36 - Protocystis slogetti Haeckel (отр. Phaeodaria); 37 - Tuscaretta tubulosa (Murray) (отр. Phaeodaria); 38 - Botryopyle setosa Cleve (отр. Nassellaria); 39 - Dorataspis heleropora Bernstein (отр. Acantharia); 40 - Tuscaretta globosa (Borgert) (отр. Phaeodaria); 41 - Tuscaretta passercula Haecker (отр. Phaeodaria)
Наряду с указанными категориями радиолярий, небольшое число видов приурочено к жизни только в холодных районах Мирового океана. Мы можем говорить об арктических и антарктических радиоляриях.
Точно так же, среди радиолярий наших дальневосточных морей работами на "Витязе" в последнее время обнаружен большой процент эндемичных видов - почти 30% по отношению ко всем 185 найденным видам радиолярий (Догель и Решетняк). Это обстоятельство подтверждает правильность отнесения дальневосточных морей к Северотихоокеанской умеренной (более реальной) зоогеографической области, установленной на основании изучения других групп животных.
Географическое распространение радиолярий может быть связано с системой течений Мирового океана. Ряд планктонных обитателей арктических вод, несомненно, приносится из северной Атлантики Гольфстримом и его ветвями (Хмызникова, 1936). В Дальневосточных морях представителей отр. Acantharia не обнаружено. Но в материалах "Витязя" 1955 г., собранных в струе Куро-Сиво, найдены акантарии в больших количествах. Они принесены этим теплым течением из тропической области.
Более полную картину зависимости распространения радиолярий от океанических течений сейчас не представляется возможным дать за отсутствием достаточных сведений.
Для ряда холодолюбивых видов современных радиолярий Геккер (Haecker, 1908) отмечает своеобразие их распределения, в зависимости от широты обитания, В антарктических водах эти виды (из отр. Phaeodaria) преимущественно свойственны поверхностным слоям, в то время как по мере продвижения на север, в умеренную или тропическую зону, эти же формы спускаются на значительные глубины. Таким видам Геккер дает название погружающихся ("submergent"). Точно такая же картина наблюдается и для ряда видов Phaeodaria, обнаруженных в с.-з. части Тихого океана, где они населяют верхние горизонты (Решетняк, 1955), тогда как Геккер для них же в более южных широтах указывает глубины значительно большие.
Наконец, у современных радиолярий в ряде случаев удается заметить разрывы в распространении. Чаще эти разрывы связаны с так называемым биполярным распространением, когда одни и те же виды обитают в умеренных областях обоих полушарий и полностью отсутствуют в тропических водах. На карте показаны примеры таких разрывов - одни и те же виды встречены в антарктических водах и в. Североатлантических и Северотихоокеанских. Л. С. Берг (1920), объясняя данное явление, говорит, что биполярность распространения организмов есть следствие ледниковых эпох. В эпохи оледенений охлаждением были затронуты не только умеренные широты, но и тропические и животные умеренных областей могли расселиться во всем. Мировом океане. Когда же ледниковый период кончился, в тропиках температура поднялась, и подобные формы либо должны были здесь вымереть, либо удалиться к северу и югу.
Второго рода разрывы распространения носят характер амфибореальных, когда известные формы (см. карту) встречаются в сев. Атлантике и в сев. части Тихого океана, отсутствуя полностью в арктических и тропических морях. Берг (1934) объясняет и амфибореальные разрывы влиянием условий ледниковой эпохи, когда в до- и послеледниковое время происходило потепление арктических вод, стало быть, имелась возможность обмена фаунами между северными частями Атлантического и Тихого океанов. С последующим похолоданием эти формы были вытеснены из Арктического бассейна и получился разрыв в когда-то бывшем сплошном ареале. Иначе говоря, разрывы в современном распространении радиолярий находят объяснение в истории развития фаун.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'