
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Краткая характеристика современных синезеленых водорослей
Микроорганизмы, объединенные под названием синезеленые водоросли, или цианобактерии, насчитывают свыше 1600 видов разнообразных одноклеточных, колониальных, нитчатых форм, характеризующихся следующими основными признаками: l) способностью осуществлять фотосинтез с выделением кислорода, в отличие от фотосинтезирующих бактерий, у которых фотосинтез протекает в анаэробных условиях и не сопровождается выделением кислорода; 2) свойством пигментного аппарата цианобактерии включать, наряду с хлорофиллом и каротиноидами, водорастворимые пигменты красного и голубого цвета, обладающие высокой способностью поглощения света, что и определяет их некоторые уникальные свойства; 3) отсутствием оформленного ядра и особенностями структуры клетки, которые отражаются и на физиологических свойствах представителей Cyanofoacteria; это своеобразие в строении клетки служит основанием для отнесения их к надцарству Procaryota (безъядерные организмы) и царству Monera /Whittaker, 1969; Маргелис, 1983/; 4)отсутствием полового процесса, дифференцированных органов воспроизведения и жгутиковых стадий; 5) способностью всех синезеленых иметь микроскопические размеры /Еленкин, 1936; Голлербах и др., 1953; Гусев, Кирикова, 1982/.
Современные синезеленые по морфологическим признакам делятся на три класса. Их краткая характеристика взята из работы С. В. Горюновой и др. /1969/.
1. Chroococcophyceae - одноклеточные и колониальные водоросли. Колония у хроококковых возникает в случае объединения нескольких разрозненных клеток посредством слизи в одно целое (рис. II. 1). Расположение клеток в колониях беспорядочное, очень редко нитевидное, но ни ложнопаренхимных слоевищ, ни настоящих нитей не образуется. Эндоспоры, экзоспоры, гетероцисты отсутствуют.
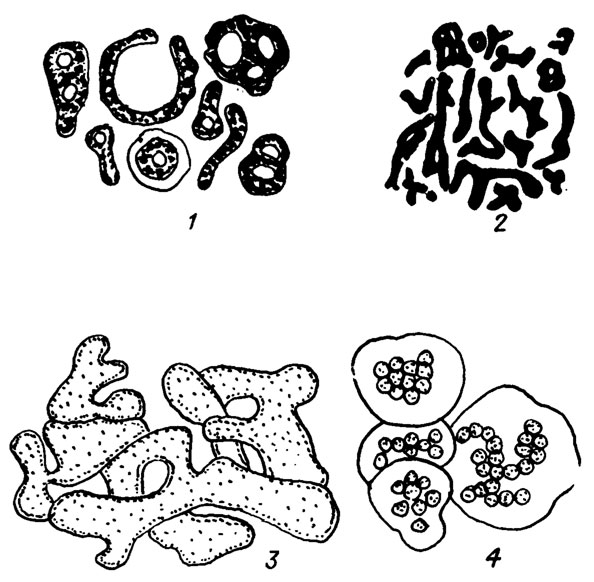
Рис. II. 1. Колонии хроококковых водорослей на примере Microcystis aeruginosa (Kutz) Elenk. Сильно увеличено (по М. М. Голлербаху и др. /1953/). 1, 3 - типичная шаровидная форма колоний, которая нередко модифицируется в эллипсоидную, реже - в цилиндрическую; 2 - силуэтное изображение колоний, имеющих причудливые очертания - от шаровидных форм до нитевидных и ветвистых, причем в слизи образуются иногда отверстия, так что колонии являются продырявленными или даже сетчатыми; 4 - обычный тип разбросанного положения клеточек по всей толще слизи
2. Chamaesiphonophyceae - одноклеточные и колониальные формы; всегда прикреплены к субстрату, размножаются они посредством эндо- и экзоспор; плазмодесмы, гетероцисты, гормогонии и влагалища отсутствуют. Клеточные оболочки большей частью толстые, упругие, ослизняющиеся. В слоевищах нити часто срастаются боковыми сторонами, образуя ложную паренхиму, в которой первоначальное строение нитей просматривается с трудом. У одноклеточных форм отчетливо проявляется дифференцирование на основание и вершину.
3. Hormogonophyceae объединяют наиболее широко распространенных представителей, клетки у которых соединены плазмодесмами и делятся только в поперечном направлении. Совокупность клеток в пределах нити носит на звание трихома. Размножение осуществляется с помощью гормогониев, т. е. обособившихся участков трихомов; у ряда форм наблюдается образование спор. Нити бывают как одинакового строения на всем своем протяжении, так и с различным строением базальных и верхушечных клеток (рис. II. 2), которые в нитях разных форм имеют цилиндрическую или бочонковидную форму и делятся только в поперечном направлении. Трихомы бывают однорядные, реже многорядные и часто окружены слизистым цилиндром, носящим название влагалища. Это трубчатое образование, открытое большей частью на обоих концах и имеющее разную консистенцию: плотную или слизистую, однородного состава или слоистую (рис. II. 3). Слои могут располагаться параллельно продольной оси или под некоторым углом, причем в последнем случае нередко наблюдается образование "воротничков" или "воронок". Трихом вместе с влагалищем носит название нити. У некоторых представителей (роды Microcoleus, Hydrocoleus, Schizothrix) нити образованы несколькими трихомами, лежащими параллельно в одном влагалище. В случае наличия внутри влагалища нескольких трихомов нить уже рассматривается не как отдельный организм, а как колония. Термин слоевище (таллом) применяется при описании некоторых колоний класса Hormogonophyceae (например, у семейства Pseudocapsosiraceae Elenk.) и является синонимом термина "колония" у гормогониевых водорослей. У многих нитчатых форм встречаются особые клетки, называемые гетероцистами, или пограничными клетками, которые характеризуются хорошо развитыми двухслойными оболочками и постепенным отмиранием содержимого. Роль гетероцист до сих пор окончательно не выяснена.
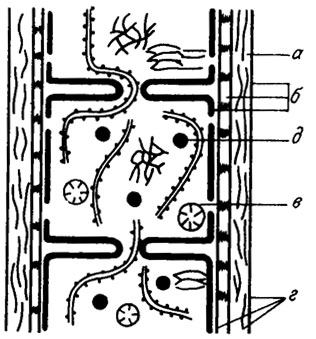
Рис. II. 2. Схема строения нити современных синезеленых водорослей /Курс..., 1981/. а - чехол; б - слои клеточной стенки; в - трихом; г - влагалище; д - гранулы с фосфатом
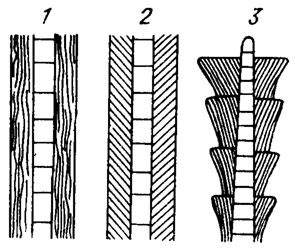
Рис. II. 3. Схематическое изображение строения слоистых влагалищ: 1 - параллельно-слоистое; 2 - косослоистое; 3 - воронкообразное, сильно увеличено (по А. А. Еленкину /1936/)
Нити Cyanophyta бывают разветвленными и не ветвящимися. Настоящее ветвление встречается почти исключительно у представителей семейства Stigonemataceae и происходит таким образом, что интеркалярная вегетативная клетка трихома делится вдоль и одна из дочерних вырастает вбок как зачаток боковой ветви. Более распространено ложное ответвление, при котором участки трихома между гетероцистами, плотно прилегающими к влагалищу, не могут расти в длину и пробивают одним концом боковую систему влагалища, образуя ложную ветвь.
Нитчатые синезеленые водоросли живут в виде изолированных нитей и колоний (слоевищ), и их скопления образуют дерновинки самого различного облика.
Синезеленые водоросли встречаются во всевозможных местообитаниях: в планктоне стоячих и медленнотекущих пресных вод, в прибрежном морском бентосе, как эпифиты и корки обрастания на различных твердых субстратах, на поверхности снега, вне воды - на влажных скалах и почве, внутри почвенного слоя. Довольно обильны и разнообразны синезеленые в горячих источниках, где они могут развиваться при температуре 65-70°.
Большинство синезеленых водорослей ведет прикрепленный, или эпифитный, образ жизни, селясь на прибрежных скалах, камнях, различных подводных предметах, на морских животных и водорослях. Среди морских синезеленых водорослей имеется значительное число космополитов, есть виды, приуроченные к теплым морям, но, по-видимому, встречается много и эндемичных форм. Основная масса синезеленых обитает в пресных водоемах.
Имеющиеся сведения об экологии синезеленых водорослей свидетельствуют о том, что температурный режим выступает фактором, определяющим последовательность смены видов и их доминирование. Подавляющее большинство синезеленых предпочитает местообитания с более высокой температурой (25-35°). Особую чувствительность к температуре водоросли проявляют на ранних этапах роста. Расцвет одного вида происходит при температурном режиме, оптимальном для него и неоптимальном для других /Сиренко, 1969/. Один из главных факторов периодического развития водорослей - дожди и связанная с ними эутрофия (принос питательных веществ). Синезеленые водоросли, как наименее чувствительные к повышенной эутрофии, вытесняют другие виды, обитающие в бассейне, и продолжают развиваться еще лучше /Кукк, 1965/. Оптимум развития большинства видов синезеленых (при колебании рН от 7,9 до 9,5) с четко выдержанным специфическим видовым составом /Сиренко, 1969/. Синезеленые водоросли являются фото- и хемоавтотрофами, известны среди них и редкие гетеротрофы, они добывают пищу либо путем фотосинтеза, либо прямым окислением восстановленных неорганических соединений, причем этот процесс осуществляется всей поверхностью клетки /Гусев, Никитина, 1979/. Толщина слизистого чехла Cyanophyta часто во много раз превышает толщину клеток, которые он окружает, и есть предположение, что он аккумулирует важные для клеток химические элементы. Это улучшает рост водоросли. Предполагают, что слизь защищает клетки от высушивания, Синезеленые водоросли приспосабливаются к температурным колебаниям, изменениям солености, влажности, интенсивности света и газовому режиму.
Велика роль синезеленых водорослей как постоянных компонентов во многих симбиотических связях с растениями и животными. Слизистая оболочка служит прекрасной средой для поселения многих бактерий, грибов и водорослей других видов /Еленкин, 1936; Горюнова и др., 1969; Штина, Панкратова, 1974; Громов, 1976/. В последнее время специалисты, изучающие синезеленые водоросли, уделяют особое внимание их цитологии, физиологии и биохимии. Синезеленые стали объектом и бактериологии /Горюнова и др., 1969; Громов, 1976; Гусев, Никитина, 1979/, не меньший интерес они представляют и для геологии как наиболее древние поселенцы планеты, хорошо сохранившиеся в ископаемом состоянии.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'