
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Глава пятая. Юрские морские пресмыкающиеся Поволжья
"Кое море всем морям мати?"
(Стих о Голубиной Книге)
"Коркодил, зверь водный, хребет его аки гребень, хобот змиев, глава василискова. А егда имать человека ясти, тогда плачет и рыдает, а ясти не перестает, а егда главу от тела оторвав, зря на неё - плачет"
"Сказания Русского Народа" (Собрание И. Сахарова. 1849).
В тяжелые годы Гражданской войны, разрухи и топливного кризиса возникла необходимость усиленной разработки местного топлива, в том числе и горючих сланцев Поволжья, около Ульяновска, Сызрани и в других местах.
Многим жителям Поволжья хорошо знакомы высокие конические отвалы глины и другой пустой породы, которые высятся горами около выработок горючих сланцев. А работники сланцевых рудников и местные краеведы время от времени находят в этих отвалах или по берегам Волги остатки крупных ископаемых ящеров, рыб, моллюсков и других обитателей моря, покрывавшего в юрское время Поволжье, почти всю Восточную Европу и большие участки других материков (рис. 61). И вряд ли найдется приволжский краеведческий музей, в котором среди других окаменелостей не оказался хотя бы один позвонок ихтиозавра или другого морского ящера...
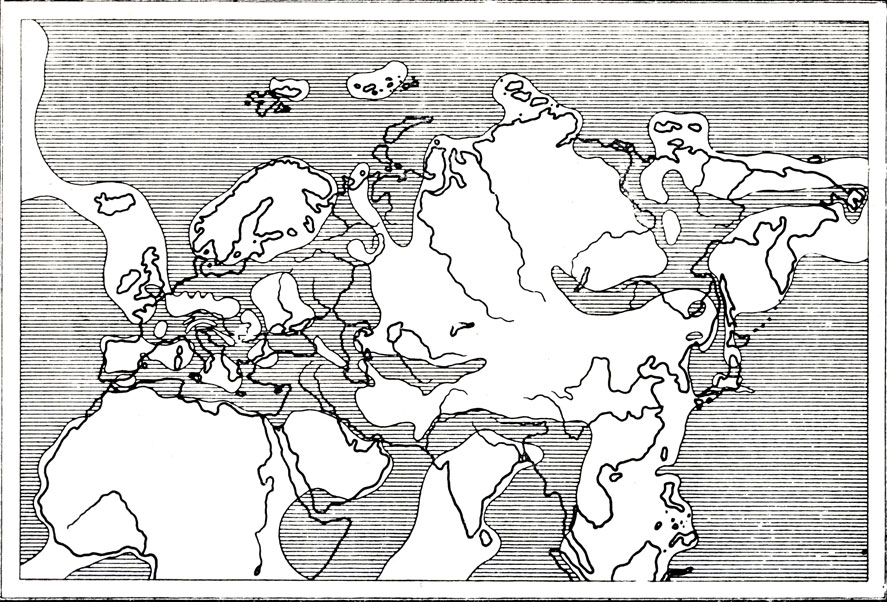
Рис. 61. Схематическая палеогеографическая карта верхнеюрского времени (по Термье, 1952)
Первые упоминания об этих пресмыкающихся в отечественной литературе относятся еще к середине прошлого столетия. Они связаны с именами выдающихся натуралистов, профессоров Московского университета К. Ф. Рулье и Г. И. Фишера, указавших на находки, сделанные в юрских отложениях на Москве-реке, у деревни Щукиной по речке Звенигородке и в других местах Подмосковья.
В Поволжье чаще всего встречаются разрозненные очень характерные позвонки ихтиозавров - короткие, напоминающие шашку, сильно двояковогнутые, часто с маленьким отверстием посредине для проходившей сквозь тела позвонков хорды (спинной струны), если только отверстие не заполнено породой (рис. 62). Сколь ни странно на первый взгляд, но находки целых скелетов и морских ящеров и появившихся много позднее морских млекопитающих (китов, тюленей) относительно редки. Причина кроется отчасти в быстром поедании трупа погибшего животного разнообразными хищными рыбами, а в прибрежной зоне птицами, наземными хищниками, крабами и т. д. К тому же у ихтиозавров и других морских ящеров, как и у китов, взаимная связь позвонков посредством суставных отростков ослаблена. В ней нет необходимости, как это требуется для сухопутных животных; у водных позвоночных потеря в весе, равная весу вытесненной телом воды, сильно уменьшает нагрузку на позвоночный столб и суставные отростки позвонков; а гибкость животного с ослаблением суставных отростков и их взаимной связи возрастает. У ихтиозавров и других водных мезозойских пресмыкающихся соединение тела позвонка с охватывающей спинной мозг нейральной дугой позвонка было, по-видимому, менее прочно, чем у наземных пресмыкающихся. Все это приводило к тому, что и позвоночный столб ихтиозавра после разрушения связок легко разваливался на отдельные позвонки, и сами позвонки распадались на "тела" позвонков и нейральные дуги. Чаще всего и находят именно тела позвонков (рис. 62, а, б) как более крупные и прочные части позвоночного столба; а нейтральные дуги оказываются захороненными отдельно и утерянными.
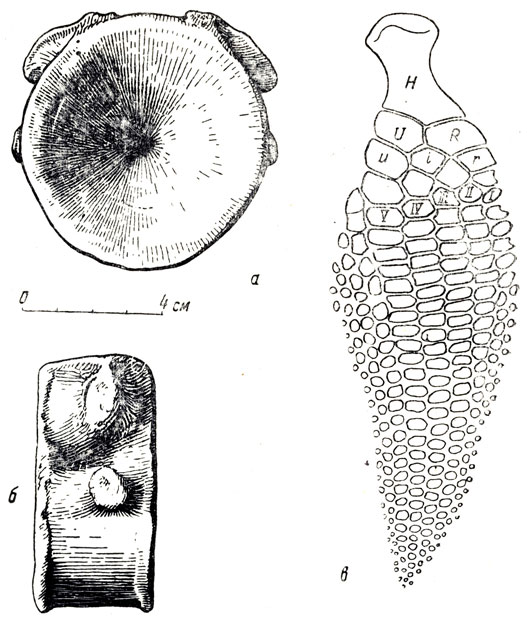
Рис. 62. Тело позвонка ихтиозавра спереди (а) и сбоку (б); видны бугорки для двойного причленения ребра (правый берег Волги около Ульяновска); скелет правой передней конечности (в) ихтиозавра эуриптеригиуса (Eurypterygius communis Conybear) H - humerus; U - ulna; R - radius; u - ulnare; i - intermedium; r - radiale. Нижняя юра Англии
Но от ихтиозавров, распространенных в морях юрского- периода по всему земному шару, известны целые скелеты, в том числе с отпечатками покрывавшей животное гладкой, лишенной крупных чешуи кожи (рис. 63) и даже с остатками самой кожи, пропитанной законсервировавшими и сохранившими ее битуминозными веществами.

Рис. 63. Стеноптеригиус (Stenopterygius quadriscissus Queiistedt). Вид со спинной стороны. Нишняя юра Англии. Британский естественно-исторический музей в Лондоне
В скелете юрского ихтиозавра, длиной обычно от двух до пяти-шести метров, бросается в глаза общая рыбообразная форма (откуда и название: греческое ихтис - рыба и саура - ящерица), вернее, быть может, дельфинообразная; далее - длинный череп с узкой вытянутой мордой и огромными глазницами € большими в виде пластинок окостенениями в стенке глазного яблока (кольцо костей белочной оболочки глаза, склеры); укороченный шейный отдел позвоночного столба, со срастанием нескольких позвонков (как у китов) в единый комплекс; остальной позвоночный столб из многочисленных очень коротких позвонков; хвостовой отдел позвоночника отогнут вниз и уходит в нижнюю лопасть сильного, вертикально поставленного хвостового плавника (у рыб хвостовые позвонки, в большей или меньшей степени видоизмененные, отогнуты в верхнюю лопасть хвоста). Парные конечности ластообразные, укороченные (рис. 64).
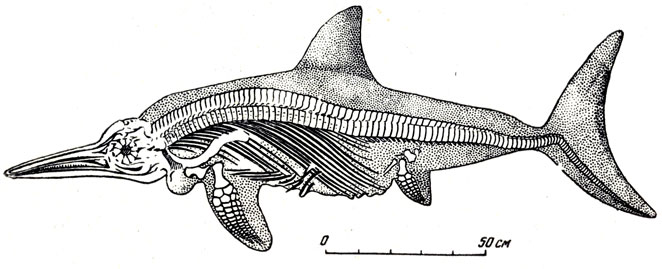
Рис. 64. Лептоптеригиус (Leptopterygius tenuirostris Conybear). Экземпляр с отпечатком кожи. Нижняя юра Холыщадена, Германия. Британский естественно-исторический музей. Лондон
Морда ихтиозавра вытянута за счет сильного удлинения межчелюстных костей; наружные носовые отверстия вследствие этого кажутся "сдвинутыми" далеко назад от конца морды и расположены рядом с глазницами, поскольку глазницы сильно расширены. Лобные кости небольшие, а лицевая часть черепа сложена главным образом межчелюстными и носовыми костями. Отверстие непарного теменного глаза расположено между передними концами теменных костей, на их границе с лобными. Многочисленные, не очень крупные зубы ихтиозавров острые, конические, приспособленные для хватания добычи.
Очень характерны конечности ихтиозавров. Непарные конечности, т. е. высокий спинной плавник и хвостовой, напоминают по своим контурам плавники рыб и дельфинов (рис. 64). Спинной был лишен костного скелета, но, вероятно, имел какую-то внутреннюю опору в виде плотной соединительной ткани, возможно даже хряща, развивавшегося независимо от позвоночного столба. Что касается хвостового плавника, то и в нем, помимо позвоночного столба, относительно размеров хвоста слабого, вероятно, были какие-то упругие соединительнотканные образования, подобно имеющимся в хвостовом плавнике китов и рыб.
Передние и задние ноги ихтиозавров (см. рис. 64) так видоизменены превращением в ласты, имевшие почти исключительно рулевое назначение, подобию грудным и брюшным плавникам рыб и ластам дельфинов, что в них нелегко "отыскать" кости кисти и стопы. Все они, за исключением плечевой, локтевой и лучевой так однородны по форме (в виде пластинок или табличек с закругленными углами), что запястные, пястные кости и фаланги пальцев совершенно сходны. Аналогично изменен скелет и задней конечности. Вдобавок очень увеличено число фаланг (типерфалангия), а у многих родов и число пальцев, доходящее до 10 (гипердактилия; рис. 62, в); в итоге общее число костей передней конечности достигает двухсот и более; ласт (вероятно был очень гибким, эластичным. Задние конечности значительно слабее передних и у большинства ихтиозавров в связи с этим и таз на пути к исчезновению. Зато хвостовой плавник был сильный и несомненно имел то же значение толкающего вперед органа, что и у рыб и китообразных. Движение вперед выполнялось, как у рыб и дельфинов, ритмически проходящими мелкими волнообразными изгибаниями тела. Эти волны изгибания, часто следуя одна за другой спереди назад, переходят на хвостовой плавник, совершающий своими лопастями гребные движения, отдаленно напоминающие работу пароходного винта; все это сообщает животному непрерывное и быстрое поступательное движение вперед. Парные плавники, ласты, имеют при этом лишь направляющее значение, в частности "рулей глубины". Спинной плавник обычно играет роль киля, напоминающего киль яхты. Положение этого "киля" на спинной стороне, а не на брюшной, удобнее при плавании и нырянии на мелководье.
Ихтиозавры перешли полностью к жизни ,в море и стали живородящими. На это указывают случаи нахождения скелетов маленьких ихтиозавров, иногда еще с не вполне окостеневшим скелетом, и поэтому часто неполностью сохранившимися, внутри скелетов крупных особей (рис. 65). Были предположения о том., что это является итогом пожирания молодых взрослыми особями. Примеры такого поедания молоди своего же вида известны для очень многих животных - от рыб до млекопитающих включительно; такой "каннибализм" не исключен, вообще говоря, и для ихтиозавров. Но в ряде случаев известны находки ихтиозавров с несколькими (до дюжины и более) скелетиками молодых особей, совершенно неповрежденными, иногда согнутыми головой к хвосту, как это свойственно молоди в утробном периоде се развития. Более того, известны находки самок ихтиозавров, погибших и быстро захороненных после рождения части молоди или в момент родов. При этом скелет молодого животного торчит из брюшной области скелета самки не головой вперед, а хвостом из задней половины брюшной части. Очевидно, у ихтиозавров рождение детенышей происходило не головой вперед, как обычно у млекопитающих, а задним концом тела, как это наблюдается у китов. У китов хвост и задняя часть тела рождающегося приходят в соприкосновение с водой, более холодной, чем материнский организм, и начинает рефлекторно oсокращаться и работать еще до полного рождения детеныша; мать и детеныш некоторое время плывут "в два хвоста"...
От ихтиозавров известны скелеты прекрасной сохранности, н особенности из Зашадной Европы (Англия, Германия и др., рис. 65). Однако обычно скелеты, находимые в сланцевых рудниках, в большей или меньшей степени сплющены давлением вышележащих слоев породы, и поэтому выставленные в музеях oскелеты ихтиозавров в больших плитах чрезмерно широкие, вернее высокие в спинно-брюшном направлении. На самом деле ихтиозавры имели веретеповидную (рис. 63), хорошо обтекаемую форму тела ящерицы или дельфина; сходство с дельфинами увеличивалось и гладкой, лишенной крупной чешуи кожей ихтиозавров. В отличие от дельфинов и других китов, у которых хвостовой плавник лежит в горизонтальной плоскости, у ихтиозавров он располагался в вертикальной. Это внешнее сходство, вызванное сходным с рыбами и дельфинами приспособлением к к одним и тем же условиям жизни ("обтекаемость" хищного животного, быстрого пловца), таково, что даже навело в свое время одного из палеонтологов на ошибочную мысль о происхождении китов непосредственно от ихтиозавров, что, разумеется, совершенно пе отвечает действительности: дельфины, вообще киты, потомки древних хищных млекопитающих, сошедших в ходе эволюции в воду, предками которых были наземные мезозойские млекопитающие; ихтиозавры же - вымершая ветвь мезозойских рептилий, возникшая в триасе и угасшая в меловом периоде, не оставившая потомков. Время расцвета ихтиозавров - нижняя юра, когда они были наиболее многочисленны. oОтличные, быстроходные пловцы, ихтиозавры охотились за рыбой, питались ГОЛОБОНОГИМИ моллюсками и в жизни моря играли примерно ту же роль, что и нынешние дельфины; но при менее энергичном обмене веществ и меньшей общей активности, чем у млекопитающих, ихтиозавры вряд ли могли бы конкурировать с дельфинами в выносливости, не говоря уже о низком уровне развития головного мозга по сравнению с млекопитающими.
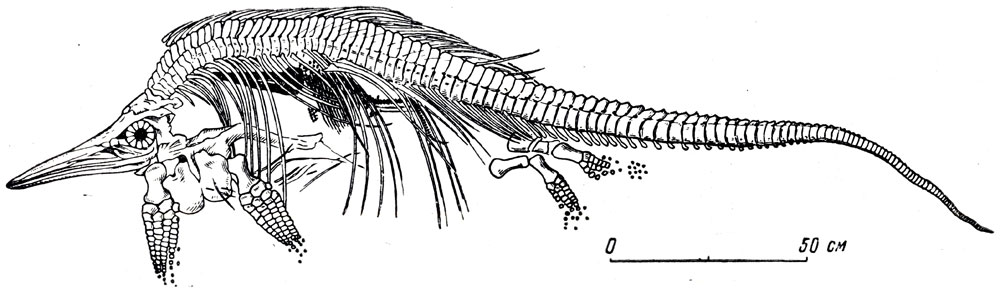
Рис. 65. Стеноптеригиус (Stenopterygius quadriscissus Quenstedt). скелет самки со скелетом детеныша внутри брюшной части. Нижняя юра. Хольцмаден, Германия. Музей Палеонтологического института АН СССР
Другие жители юрского моря и современники ихтиозавров почти на всем протяжении их истории - плезиозавры - представляли собой иной тип приспособления к жизни в море (рис. 66, 67). Небольшая, а у некоторых относительно маленькая голова с пастью, вооруженной высокими, редкими, острыми коническими зубами, длинная "лебединая" шея за счет многочисленных позвонков, у большинства короткое широкое уплощенное туловище, короткий хвост, явно не игравший роли в плавании, и мощные веслообразные передние и задние конечности почти одинаковых размеров и строения - таковы характерные черты плезиозавра.

Ихтиозавры и плезиозавры. Рис. К. К. Флерова
Ихтиозаврам быстрое поступательное движение сообщали мелкие волнообразные ритмические изгибания тела, пробегавшие от головы назад и переходившие на мощный хвост, у плезиозавров же плавание явно выполнялось передними и задними ластами; они действовали, как весла, сообщая поступательное движение и регулируя направление. Плечевой пояс и таз плезиозавров состоял из костей, сильно уплощенных в связи с гребным характером работы мускулатуры конечностей. Верхние, "спинные" части переднего пояса и таза, т. е. лопатка и подвздошная кости, имеют незначительную величину; брюшные же части плечевого пояса и таза не только широкие, но и сильно разрослись вперед и назад (рис. 66). Это обеспечивало широкую поверхность и мощное развитие "гребных" мускулов, прикреплявшихся к относительно длинным плечевым и бедренным костям, притом не только для мускулов, "отводивших" веслообразные ласты назад, но и для мускулов, "заносивших" конечности вперед. Комбинированные движения четырех ластов, вероятно, давали плезиозаврам не только быстроту плавания, но и возможность быстрых поворотов на месте, общую подвижность. Характерно слабое развитие суставных отростков на дугах позвонков. Ребра имели лишь одно соединение с позвонком (с его поперечным отростком), это давало большую подвижность, чем при обычном двойном соединении - с телом позвонка и с дугой. Довольно высокие остистые отростки указывают на сильное развитие спинной мускулатуры. Плезиозавр представлял своеобразную весельную "двойку" и несомненно был превосходным пловцом, однако он вряд ли мог конкурировать в быстроте и подвижности с ихтиозавром. Ловле добычи, быть может, отыскиванию добычи на дне, помогала длинная гибкая шея. Широкие сильные конечности, вероятно, давали возможность плезиозаврам вылезать время от времени на берег, как это делают тюлени и другие ластоногие, что вряд ли было доступно ихтиозаврам, окончательно порвавшим с сушей и ставшим полностью "моряками", подобно дельфинам. Возможно, плезиозавры были в известной степени прибрежными жителями, и морское прибрежье могло давать им убежище и от более подвижных в воде ихтиозавров, для которых вода стала родной стихией, и от самых страшных морских хищников юрского моря, огромных плиозавров (рис. 68-71). Косвенным указанием на такой выход, вернее на выползание на сушу плезиозавров, вероятно, можно считать сильное развитие брюшных ребер, иногда образующих у плезиозавров прочный решетчатый скелет брюшной стенки.
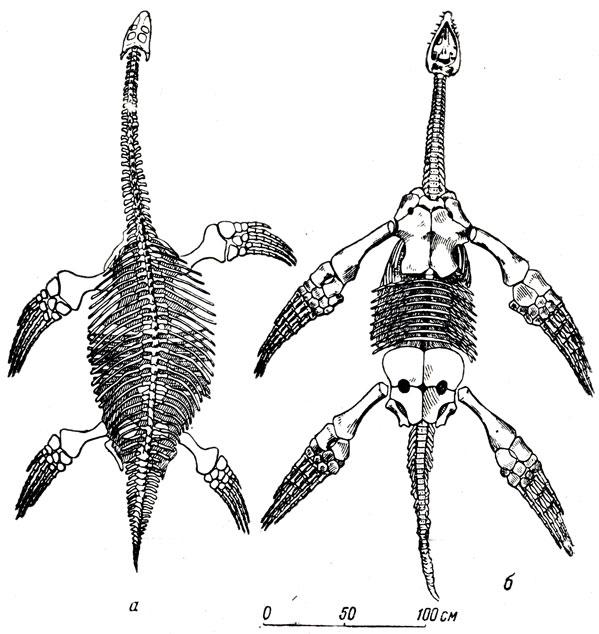
Рис. 66. Скелеты плезиозавров. а - Cryptocleidus oxoniensis Phil, из верхней юры Англии. Вид сверху. Американский естественно-исторический музей в Нью-Йорке; б - Thaumatosaurus victor E. Frass из нижней юры Хольцмадена. Германия. Вид снизу. Музей Штуттгарта
Самые крупные хищные пресмыкающиеся волжского юрского моря, да, вероятно, и вообще когда-либо существовавшие в морях, - огромные плиозавры, достигавшие восьми метров длины, с черепом двухметровой длины и более, вооруженным огромными, иногда трехгранными, зубами с зазубреной по ребрам эмалью (рис. 68, а). В позвоночном столбе передняя и задняя поверхности тел позвонков плоские. Пояса парных конечностей, как у плезиозавров, широкие, плоские, длинные набрюшной стороне; сами конечности большие, веслообразные с увеличенным числом фаланг. Всем этим плиозавры очень напоминают плезиозавров, с которыми объединяются в один общий отряд "зауроптеригий" (греческое "саура" ящер и "птеригион" - крыло, в данном случае - ласт). Но для плезиозавров характерна короткая голова и длинная гибкая шея, помогавшая настигать добычу, тогда как у плиозавров шея короткая, малоподвижная, и животное должно было настигать добычу стремительным броском вперед. Передние зубы, коронки которых в поперечнике достигают шести сантиметров и более, глубоко всажены в челюсть и торчат вершинами несколько в бок; при этом верхние входят свободно в промежутки между нижними. Огромная пасть при челюстях до полутора, двух и более метров длины (в литературе описаны черепа до трех и более метров), вооруженная мощными острыми зубами со слегка согнутыми коронками, явно рассчитана на крупную добычу, вероятно, вплоть до самых больших плезиозавров. Плиозавры не могли соперничать с плезиозаврами в ловле мелкой изворотливой добычи, но отнюдь не должны были уступать плезиозаврам в быстроте, как пловцы (рис. 69, 70).

Рис. 67. Плезиозавр (рис. К. К. Флерова). Музей Палеонтологического института АН СССР
лш
У плиозавров очень своеобразно соотношение некоторых костей лицевой части и крыши черепа (рис. 68, 71). Межчелюстные кости (Рт) надвинуты задними концами надлобные (Fr) и достигают передних концов теменных костей (Ра), а теменные, находя вперед на лобные и вклиниваясь по средней плоскости черепа между лобными, раздвигают их на некотором протяжении в стороны. В общем верхняя сторона лицевой части черепа, вследствие надвигания костей на значительном протяжении друг на друга, местами в три слоя, при большой общей поверхности швов между взаимно перекрывающими друг друга костями представляет очень совершенную по прочности конструкцию: длинная лицевая часть черепа хорошо приспособлена для сопротивления напряжениям на излом, возникающим при захлопывании пасти, или стремлении крупной добычи вырваться из пасти. Море было богато рыбой, головоногими моллюсками, остатки которых встречаются внутри брюшной области скелетов плиозавров, и другой добычей.
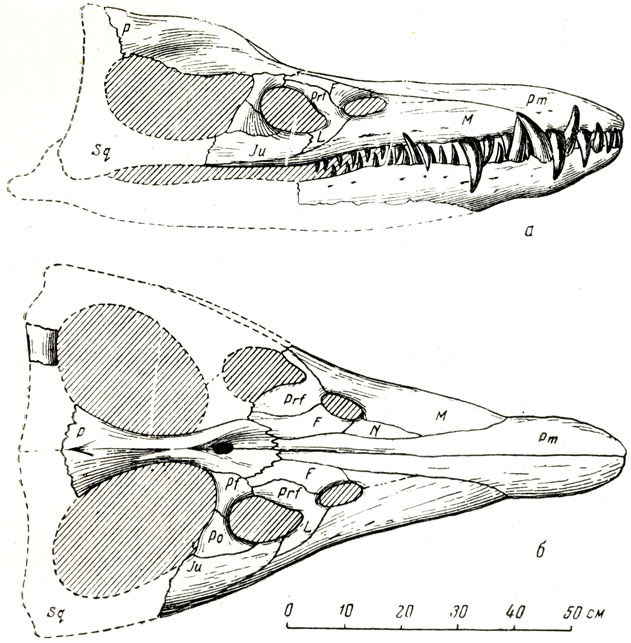
Рис. 68. Череп плиозавра (Pliosaurus rossicus Novojilov) Вид сбоку (а) и сверху (б). Нижневолжский ярус, Буинский сланцевый рудник Чувашской АССР. Музей Палеонтологического института АН СССР (Н. И. Новожилов, 1948)
Таковы в кратких чертах были хищные ящеры. Их остатки встречаются в отложениях морей юрского периода, в том числе и юрского моря Поволжья (В отложениях морей юрского периода найдены также скелеты очень интересных морских крокодилов (и в современную эпоху некоторые крокодилы, например крупный Crocodilus porosus Зондских островов, иногда встречаются в море за несколько сот километров от берега). Кроме того, в то время уже были и морские черепахи, но в юрских отложениях Поволжья пока не обнаружено остатков крокодилов и черепах, хотя в прибрежных зонах юрских морей Поволжья и крокодилы и черепахи почти наверняка были). Обычно скелетные остатки, находимые в этих осадках - в глинистых прослоях сланцевых рудников, - сильно минерализованы, именно пиритизированы. При окислении на воздухе, особенно влажном, сера пирита дает сернистые окислы, образующие с водой воздуха сернистую кислоту и т. д. Итог этого - почти неизбежная гибель пирити-зированных палеонтологических материалов, если не принять своевременно мер для их сохранения. Необходимые условия сбережения материала: хранение в сухом помещении, тщательная пропитка костного материала спиртовым раствором лака, препятствующего проникновению влаги воздуха и покрытие с этой целью лаком поверхности костей и зубов.
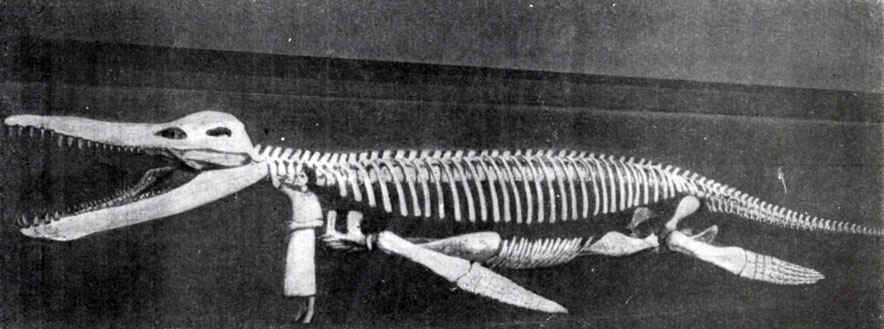
Рис. 69. Кронозавр (Kronosaurus), гигантский плиозавр из юрских отложений Квинсленда (Австралия). Музей зоологии и сравнительной анатомии им. Агассица в Харварде, Массачузетс, США
Однако неизмеримо большее, чем в музеях, количество палеонтологических материалов погибает при разработке сланцев в рудниках из-за непонимания огромной научной ценности черепов и скелетов, встречающихся во время добычи сланцев. Известны случаи, когда скелеты огромных плиозавров уничтожались, так как "мешали" выработке сланца, и разбитые на куски черепа и кости шли в отвал вместе с пустой породой...

рис. 70. Плиозавр (Macroplata swinton). Реконструкция Вильсона. (из Свистона, 1958)
Настоящий очерк имеет, между прочим, целью заинтересовать персонал сланцевых рудников теми замечательными научными ценностями, которые должны были бы становиться гордостью нашей науки, а не погибать бесследно.
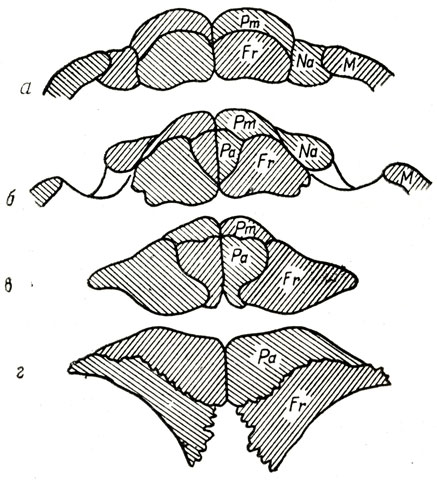
Рис. 71. Вертикальные поперечные разрезы крыши черепа плиозавра (Pliosaurus rossims Novojilov), наблюдаемые на изломах а - в области наружной боковой границы носовых (Na) костей с верхнечелюстными (М); б - у передних краев наружных носовых отверстии; в - у задних краев носовых отверстий; г - впереди переднего края отверстия теменного глаза; Рm - praemaxillare; Fr - frontale; Na - nasale; N - maxillare; Pa - parietale (H. И. Новожилов, 1948)
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'