
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Оценка систематического значения проявлений конвергентного и параллельного развития
Конвергенция. Под конвергенцией понимается наличие у отдельных родственных групп организмов сходных морфологических или функциональных признаков, появление которых обусловлено не общностью происхождения, а приспособлением к более или менее одинаковым условиям существования.
Конвергентное развитие приводит к появлению у той или иной группы организмов комплекса объединяющих особи этой группы признаков, отсутствующих у их предков, но имеющихся у отдаленных в филогенетическом отношении группах, в которых ранее, одновременно или позднее появился этот комплекс признаков, будучи вызванным у всех этих групп приспособлением к одинаковым условиям жизни. Конвергенция может наблюдаться у различных типов, классов, отрядов. В качестве примера можно привести червеобразную форму внешнего скелета у простейших - некоторые Ammodiscidae, Schwagerinidae (Nipponitella), Cornuspiridae (Rectocornuspira). Ophthalmidiidae и другие, червей (Polychaeta-Serpula, Spirobis), моллюсков (у брюхоногих Vermetidae, у головоногих Nipponites, Anisoceras, Ancyloceras); бокалообразную - у фораминифер (Umbella), губок (Ventriculites), археоциат, кораллов, моллюсков (Hippurites), брахиопод (Richthofenia), хордовых (Ascidia); веретенообразную - у рыб, рептилий (ихтиозавры) и млекопитающих (китообразные); башенкообразную - у раковины аммонита Тurrilites и брюхоногого моллюска Turritella. Крайне напоминают раковины двустворчатых моллюсков раковины ракообразных Ostracoda и Phyllopoda.
В перечисленных случаях конвергентной является общая форма тела или внешнего скелета. Конвергентно могут развиваться отдельные органы (это так называемые аналогичные органы, общие по функции, но разные по происхождению). В качестве примера можно привести крышечки у кораллов (Goniophyllum, Calceola), брахиопод (Richthofenia), двустворчатых моллюсков (Requienia, Hippurites), брюхоногих моллюсков (Neritidae, Trochidae, Naticidae), ракообразных (некоторые Cirripedia). Аналогичны раковины у кораблика (Nautilus) и аргонавта (Argonauta argo). У наутилид (и аммонитов) раковины выделяются мантией и служат органом защиты и плавательным аппаратом, а у аргонавта образуются дорсальными руками и только самками. Они являются выводковыми сумками. В качестве особого вида конвергенции можно рассматривать отдельные случаи мимикрии (и, в частности, миметизма).
На основании признаков конвергентного характера обладающие ими организмы не могут объединяться в систематические группы. Эти признаки иногда называют несистематическими. Однако в отдельных семействах, родах и видах эти признаки и детали могут иметь большое систематическое значение. Как указывалось выше, крышечка является аналогичным органом у ряда организмов. Но по крышечкам, точнее, по деталям строения их можно различать семейства, роды и даже виды брюхоногих моллюсков. Веретенообразная форма тела у рыб, рептилий и китообразных конвергентна. Однако известно, что особенности формы тела рыб широко используются в их систематике. Червеобразная форма наружного скелета беспозвоночных в целом конвергентна, но детали этой формы, как видно из приводимого примера (см. схему), имеют важное систематическое значение.
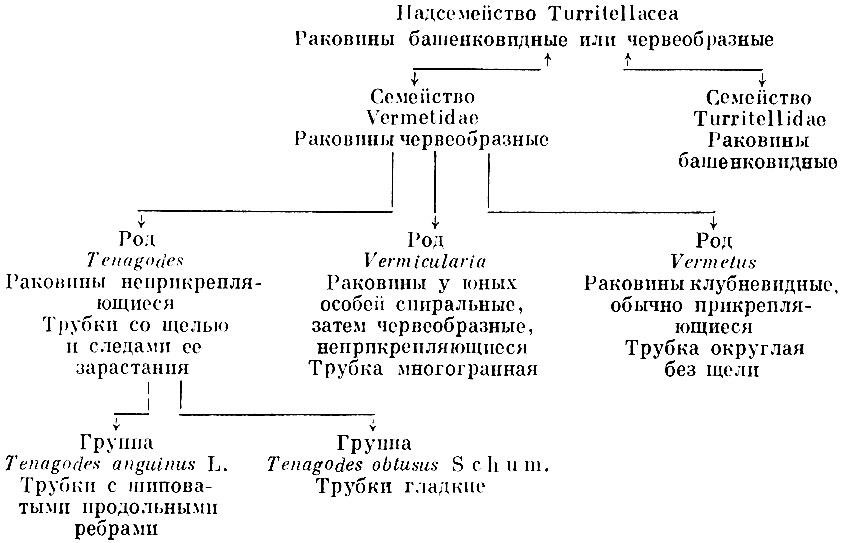
Червеобразная форма наружного скелета беспозвоночных
Как видно, к оценке систематического значения конвергентно возникших признаков требуется особый подход. В приведенном примере к надсемейству Turritellacea и семейству Vermetidae лишь на основании признака червеобразности раковин могли быть отнесены брюхоногие: Avardaria (Rissoacea), Pseudomalaxis (Extractrix) extractrix Воetger (Epitoniacea), а равно и аммонит Nipponites. Следует особо отметить, что до сих пор окончательно не решен вопрос, относится ли ископаемый род Burtinella (= Tubulostium) к брюхоногим моллюскам или к червям. Высказывается предположение, что этот род является сборным, включающим червей и моллюсков семейства Vermetidae с резко выраженными конвергентными признаками.
Гомеоморфия. Под гомеоморфией понимается сходство организмов близких филогенетических ветвей, возникшее в результате параллельного развития в более или менее общих условиях обитания. И. И. Шмальгаузен [1946а] указывает, что проведение грани между конвергенцией и параллелизмом в развитии бывает затруднительно. Поэтому каждый раз необходимо выяснить природу подмеченного сходства. "Если приспособительное сходство развилось совершенно независимо на базе первоначальных различий (аналогия органов), то мы будем говорить о конвергенции. Если же такое же приспособительное сходство развилось, хотя и независимо, но на базе сходств, унаследованных от общих предков (в гомологичных органах), то следует говорить о параллелизме [Шмальгаузен, 1964а, стр. 434]. Гомеоморфия будет пленарной, или полной, когда распространяется на целые таксоны, например, на популяции видов одного рода, на виды родов одного семейства, на роды семейств одного отряда, на семейства отрядов одного класса. В зависимости от ранга таксонов, на уровне которых проявляется гомеоморфия, можно различить следующие виды.
1. Видовая гомеоморфия - сходство видов одного рода при параллельном развитии в близких условиях изолированных биотопов. Прекрасным примером могут служить широко известные факты поразительного сходства скульптированных вивипарид Дунайского плиоценового бассейна с ныне живущими в водоемах Восточного Китая, а также плиоценовых унионид Румынии с современными из южных штатов Северной Америки.
2. Родовая гомеоморфия - сходство подродов или родов одного семейства, но параллельных ветвей развития. Например: внешнее сходство сарматского вида Cardium (Cerastoderma) fittoni Orb. и киммерийского вида Caladacna esheri Mayer, а также близость миоценового рода Eoprosodacna и плиоценового Prosodacna; неоднократное параллельное возникновение грифоидных устриц, относимых к искусственному роду Gryphaea и не менее искусственному подсемейству Gryphaeinae.
3. Семейственная гомеоморфия - сходство семейств одного отряда в параллельных ветвях развития. В качестве примера можно привести семейства Patellidae и Acmaeidae. Колпачкообразные и блюдцеобразные раковины этих семейств внешне крайне трудно различимы, но существенно разнятся по анатомическим особенностям и наличию разных протоконхов - спирального у Patellidae и конического у Acmaeidae. Классическим примером параллельного развития издавна служат семейства Equidae и Palaeotheriidae непарнопалых животных.
4. Отрядовая гомеоморфия - сходство отрядов одного класса в параллельных ветвях развития. Австралийские сумчатые (Marsupialia) имеют много родов, крайне напоминающих различные роды других отрядов млекопитающих (сумчатый тушканчик, сумчатый крот, сумчатая соня, сумчатый волк и др.).
Гомеоморфия будет партикулярной, или частичной, если лишь отдельные виды или отдельные роды крупных ветвей развития обнаруживают признаки сходства. В. Н. Шиманский [1956, стр. 30-32] приводит следующие яркие примеры гомеоморфии: 1) поразительное сходство раковин юрского аммонита Ringsteadia cuneata (Тrautsсh.) и триасового наутилоида Gonionautilus securis (Dittmar); 2) сходство раковин некоторых наутилоидей (например, Trocholitidae) и климений; 3) сходство количества и расположения элементов перегородочной линии у аммонита Manticoceras neverovi Воgоsl. из девона Алтая и наутилоида Permoceras bitauniense (Haniel) из нижней перми Тимора; 4) поразительное сходство раковин упомянутого выше Permoceras bitauniense (Haniel) и Рseudonautilus geinitzi (Орреl) из верхней юры Западной Европы ("сходство между этими двумя видами, принадлежащими к двум разным надсемействам и даже подотрядам, не больше, чем между двумя видами одного рода"). Известен ряд случаев сходства конфигурации перегородочных линий у некоторых родов аммоноидей разных ветвей развития.
Как гомеоморфное сходство рассматривается внешнее сходство раковин триасового рода Tetractinella (семейство Athyridae, у которых спиральный ручной аппарат) и раковин верхнеюрского рода Theirothyris (семейство Zeilleriidae, у которых петлевидный ручной аппарат), а также сходство раковин некоторых видов семейства Enteletidae с раковинами ряда видов семейства Rhynchonellidae, резко различных по внутреннему строению.
В этом отношении заслуживает внимания параллельное возникновение колпачкообразной раковины у представителей разных отрядов брюхоногих моллюсков (см. схему).
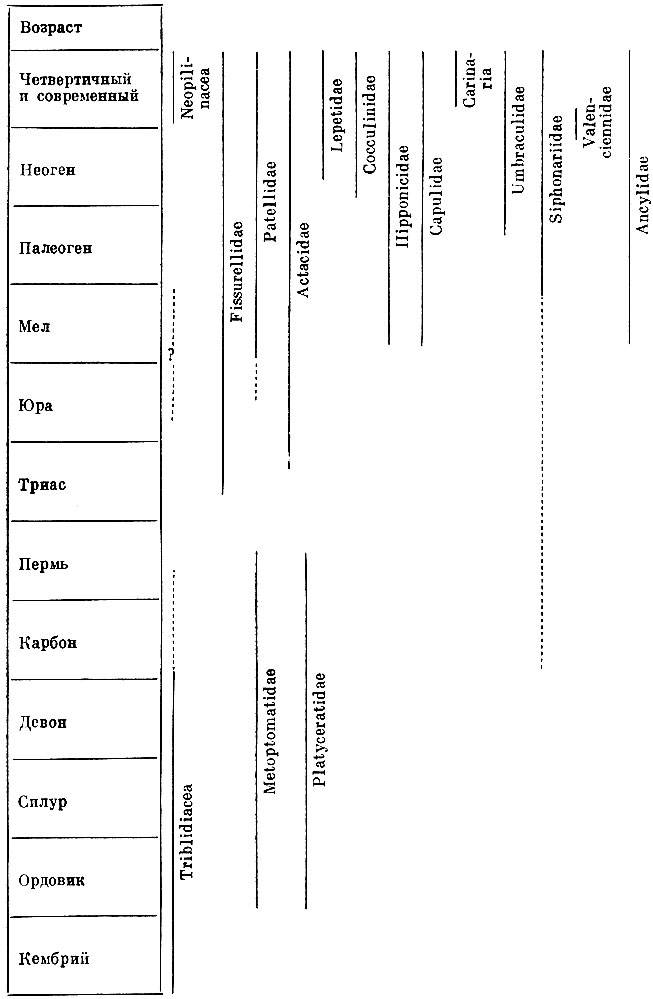
Параллельное возникновение колпачкообразной раковины у представителей разных отрядов брюхоногих моллюсков
Следует особо упомянуть двустворчатых брюхоногих моллюсков, раковины которых настолько имитируют раковины двустворок, что их действительное систематическое положение может быть определено лишь после специальных исследований [Сох, Rees, 1960; Keen, Smith, 1961]. В данном случае речь может идти скорее о конвергентном сходстве, нежели о гомеоморфном, так как створки Двустворчатых брюхоногих не гомологичны своркам Bivalvia.
Изучение параллелизма в развитии организмов позволяет обнаружить неточности в систематике отдельных групп, так как показывает их гетерогенность и вскрывает ее причину. Ярким примером сказанного может служить семейство Pectinidae, в котором в качестве самостоятельных родов выделялись экологические формы. Эти так называемые роды объединяли виды, принадлежащие к различным ветвям развития, но приобретшие общие признаки из-за сходства условий обитания. Таковы Pseudamussium, Lissochlamys, Hinnites, Lissopecten и др.
По литературным данным уже давно [Dacque, 1921; Trueman, 1922; Zeuner, 1933] известно, что устрицы при переходе от прикрепленного образа жизни на твердых грунтах к свободному обитанию на мягких грунтах приобретают грифоидный характер. Было показано несколько разновозрастных рядов (юрских и меловых) перехода устриц в грифеи. Однако эти факты не были учтены О. С. Вяловым при разработке классификации устриц, в которой [Вялов, 1936] грифеи" не только объединялись в особый род, но выделялись в подсемейство Gryphaeinae.
Ошибочность такого представления была доказана Р. Ф. Геккером на основании углубленного морфологического и экологического изучения устриц палеогена Ферганы [Геккер, Осипова, Вельская, 1962]. Р. Ф. Геккер показал, что О. С. Вялов под родовым названием Gryphaea объединил группы морфологически различных устриц, принадлежащих к самостоятельным ветвям развития: "Мы полагаем, что аналогичная судьба, т. е. изменение принадлежности к более высоким систематическим категориям постигнет также другие роды и подроды в системе устриц О. С. Вялова ... по той причине, что эта система построена в значительной степени на формальных признаках"* [Геккер, Осипова, Вельская, 1962, стр. 127]. Подтверждения этому мы находим и в статье К. Бейрлена [Beurlen, 1958] об экзогирах, где он пишет, что данная О. С. Вяловым классификация экзогир представляет формальную, искусственную, "пульверизированную", как он выражается, классификационную схему.
* (Подобная мысль была высказана ранее И. А. Коробковым [1954, стр. 197].)
Такие названия, как Hinnites, Lissochlamys, Lissopecten, Pseudamussium, Gryphaea, Exogyra, и многие другие настолько широко вошли в практику, что несмотря на искусственность объединяющихся под этим названием групп их следует сохранить как наименования направленной специализации. Эти искусственные группы, объединяющие на подродовом или родовом уровне формы, появившиеся в результате параллельного развития, И. А. Коробковым [1957а, стр. 32-33] предложено называть приспособительными формами (Forma accommodata). В отличие от геноморф Ланга и Трумена они объединяют гомеоморфные, а не конвергентные формы и принадлежат к одному роду. При описании названий видов, принадлежащих к приспособительным формам, целесообразно писать в квадратных скобках за родовым названием наименование приспособительной формы. Например: Chlamys [Forma accommodata Hinnites] corstii Defr. [Коробков, 1957а, стр. 15 и 16].
Выявление параллелизма в развитии ископаемых организмов, так необходимое для проверки правильности их классификации, часто бывает затруднено отсутствием достаточного количества материала, его неполной сохранностью и формальным подходом к его изучению. Важно отметить, что признаки параллелизма всегда подчинены признакам непосредственного родства. Иными словами, признаки сходства в данном аспекте менее существенны, чем признаки различия. Cardium (Cerastoderma) fittoni Оrb. и Caladacna esheri Mayer сходны по форме створок и характеру скульптуры, что обусловлено обитанием раковины на песчаном грунте солоно-ватоводных бассейнов. Но эти двустворки существенно разнятся не только по видовым, но главное и по родовым признакам - у Caladacna нет латеральных зубов, а у Cerastoderma их четыре; у Caladacna два кардинальных зуба в правых створках и по одному в левых, а у Cerastoderma по два кардинальных зуба в каждой створке.
Далеко не всегда наличие признаков сходства у видов разных родов одного семейства или у родов разных семейств одного отряда, т. е. признаков сходства между организмами в обособленных ветвях развития, может рассматриваться как проявление параллелизма в развитии. Наличие указанных признаков не только возможно, но даже обязательно как при дивергентном развитии, так и при адаптивной радиации. Часто такие признаки ошибочно рассматриваются как проявления гомеоморфии, под которой понимается поверхностное сходство между таксонами при более или менее глубоком различии в существенных чертах строения [George, 1962]. Такое определение гомеоморфии неполное. Пропущено основное в характеристике этого явления - отражение параллелизма в развитии. Например, совершенно неправильно рассматривать как проявление гомеоморфии сходство морфологии начальных стадий при различии поздних [Миклухо-Маклай, 1961, стр. 35]. Примеров такого сходства бесчисленное множество. Именно оно кладется в основу доказательства дивергентности развития.
Не отвечает понятию параллелизма в развитии так называемая трансверсальная гомеоморфия, т. е. такое явление, при котором организмы разных филогенетических рядов обнаруживают в известный момент (в точке схождения или в точке скрещивания) сходство по признакам, формировавшимся в этих рядах при противоположных тенденциях. Если, например, виды ряда А характеризуются последовательным усилением признака, а виды близкородственного ряда В, наоборот, в процессе развития постепенно утрачивают этот признак, то в первом ряду будет такой вид, который по степени развития данного признака будет соответствовать виду второго ряда. Как видно, такое сходство объясняется не параллелизмом в развитии.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'