
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Лицевой отдел
Лицевой отдел (рис.1) в большинстве случаев значительно удлинен и довольно низок, в особенности, у ряда "мелких" хищных динозавров - "узкоголовых" дромеозаврид (велоцирапторин), заурорнитоидид, орнитомимид и т.д. Вместе с тем при укрупнении черепа лицевой отдел нередко становится довольно высоким, что свойственно прежде всего карнозаврам и "высокоголовым" дромеозавридам (дромеозавринам). Овирапториды при небольших размерах характеризуются, пожалуй, наиболее укороченным лицевым отделом.
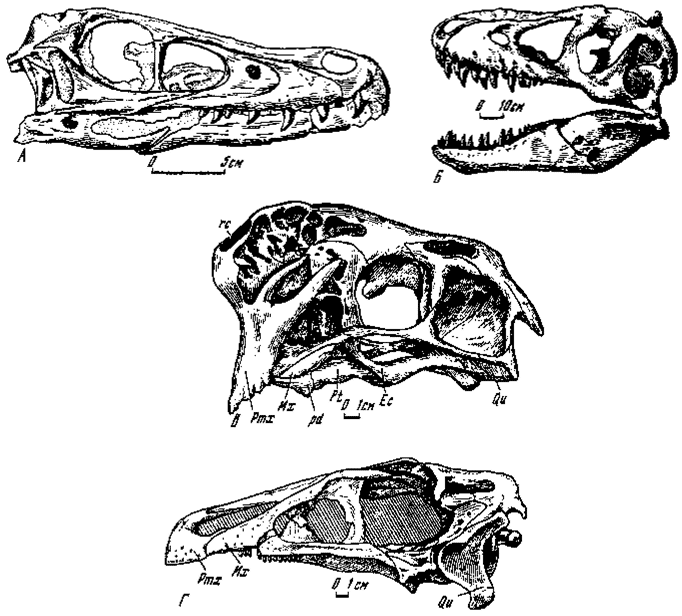
Рис. 1. Череп хищных динозавров: А - Velociraptor mongoliensis Osborn (№ 100/25, справа); Б - Tarbosaurus bataar Maleev (№.100/59, слева); В - Oviraptor philoceratops Osborn (№ 100/42, слева); Г - Erlicosaurus andrewsi Perle (№ 100/111, слева)
Верхняя челюсть была зубатой, как у большинства групп или же беззубой. Орнитомимиды (к ним близки гарудимимиды) представляют первую, ставшую известной, группу беззубых хищных динозавров (Marsh, 1881) снабженных роговым клювом. Позднее к ним прибавились овирапториды (Барсболд, 1976 а, б; Osborn, 1924), а в последнее время - ценагнатиды (Osmolska, 1976), у которых пока известна лишь нижняя челюсть (Sternberg, 1940; Cracraft, 1971). Своеобразное место занимает эрликозавр, передняя часть челюстей которых была беззубой. Верхняя челюсть у орнитомимид, слагавшая костную основу рогового надклювья, была удлиненной и слегка расширенной, т.е. в целом сохраняла пропорции, известные у зубчатых групп. Полной противоположностью служит верхняя челюсть у овирапторид - она была чрезвычайно высокой и в ее составе основное место принадлежит межчелюстным костям с широкой латеральной поверхностью, служившим основой для рогового надклювья. Челюстные кости были сильно сокращенными, что нехарактерно для хищных динозавров. У эрликозавра общие пропорции верхней челюсти примерно такие же, как у зубатых групп, однако межчелюстные и самые передние концевые участки челюстных костей были беззубыми, тогда как большая часть челюстных костей имела мелкие зубы "хищного" характера. Размеры и форма беззубой части верхних челюстей свидетельствуют об укороченности и небольшой расширенности рогового надклювья.
Наружные носовые отверстия находились обычно на переднем конце лицевого отдела. Лишь у овирапторид они смещены назад и сильно вверх, выше предглазничных отверстий, что представляет собой существенное отклонение от нормы у хищных динозавров. Не совсем обычны размеры носовых отверстий у эрликозавра, у которого они достигают более чем половины длины лицевого отдела.
Челюстная кость впереди предглазничного отверстия прободена одним - двумя маленькими отверстиями, которые обычно называют дополнительными предглазничными (Osborn, 1912). Однако строение образующей их части челюстной кости в медиальном плане оказывается довольно сложным. Изнутри в носовых отростках челюстных костей обособляются сообщающиеся друг с другом полости, разделенные перегородкой - максиллярной септой (Madsen, 1976a). Тонкость медиальной стенки не способствует их сохранению в ископаемом состоянии, и это препятствовало до сих пор более детальному ознакомлению с их строением. Подобные структуры отмечались у цератозавра под названием челюстных полостей, или максиллярных синусов (Hay, 1908), впоследствии - у аллозавра и маршозавра (Madsen, 1976a), вероятно, у дейнонихуса (Ostrom, 8 1969a, b), а также у монгольского анкилозавра сайхании (Maryanska, 1977). По-видимому, эти полости связаны с носовой и тогда их следует называть максиллярными синусами. Они начинаются сразу сзади наружных носовых отверстий, латерально открываясь максиллярными отверстиями. У овирапторид из-за задне-верхнего смещения наружных носовых отверстий максиллярные синусы располагаются над ними. У аллозаврид (Madsen, 1976a) синусы, состоявшие из двух камер, разделенных максиллярной септой, были перфорированы спереди и сзади. Каждая из камер имела отдельное медиальное и дорзальное отверстия. Синусы протягиваются вдоль участка внутренней поверхности челюстных костей, по длине занимаемого пространства, совпадая с контактирующими друг с другом их небными отростками, что хорошо наблюдается у тираннозаврид.
Верхний край лицевого отдела у некоторых групп рельефно скульптирован. У цератозавров это крупный выступ в виде расширенного рога на носовых костях, у других - парные скульптированные гребни, протягивающиеся вдоль краев носовых и переходящие на слезные кости. Наибольшего выражения они достигают у дилофозавра, в меньшей степени - у аллозаврид (Marsh, 1884; Hay, 1908; Gilmore, 1920; Madsen, 1976a, b). Однако это все более ранние, юрские формы. В поздних же ветвях известен лишь единственный случай развития крайне своеобразного костного выроста. Это непарный, сжатый с боков, высокий продольный гребень у овирапторид (Барсболд, 1981), протягивавшийся от верхней части межчелюстных до переднего края лобных костей включительно (рис. 2). Межчелюстная кость помимо обычных носового и челюстного отростков несет еще один, самый передний, восходящий, который и формирует нижне-передний край костного гребня. Носовые и челюстные отростки ограничивают крупные удлиненные наружные носовые отверстия. Большая задняя часть гребня, по-видимому, образована носовыми костями, крайне узкие удлиненные передние отростки которых слагали верхний край наружных носовых отверстий, над которыми проходит наиболее высокая часть гребня. В его заднем окончании участвуют своими передне-медиальными массивными отростками лобные кости, к которым переходит полого спускавшийся дорзальный край гребня. Боковая поверхность его не сплошная, она состояла из сплетения тонких костных перекладин. Латеральные ажурные стенки гребня были близко сдвинуты, возможно, почти до соприкосновения друг с другом. Передне-верхний более или менее сплошной костный край гребня нес узкие и удлиненные парные отверстия или карманы неясного назначения, разделенные продольной перегородкой. Глубина отверстий могла ограничиваться шириной сплошного костного края или же они свободно сообщались с узким пространством между стенками гребня. Ажурность его стенок свидетельствует, что гребень мог быть покрыт роговым шлемом (как у казуаров), что требовало обильного кровоснабжения. У казуаров костная основа рогового шлема занимает крышу черепа вплоть до затылочных костей и также отличается значительной рыхлостью костной ткани. Следует отметить, что костные гребни отмечены в черепе у крупных особей собственно овирапторов и отсутствуют у других овирапторид - ингений. У единственного черепа меньшего размера, с некоторыми сомнениями относимого к овираптору, гребень отсутствует, что позволяет видеть в нем возрастной (половой) признак.
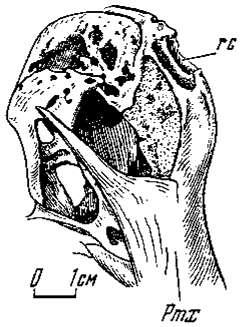
Рис.2. Костный лицевой гребень Oviraptor philocerators (№100/42, спереди и справа)
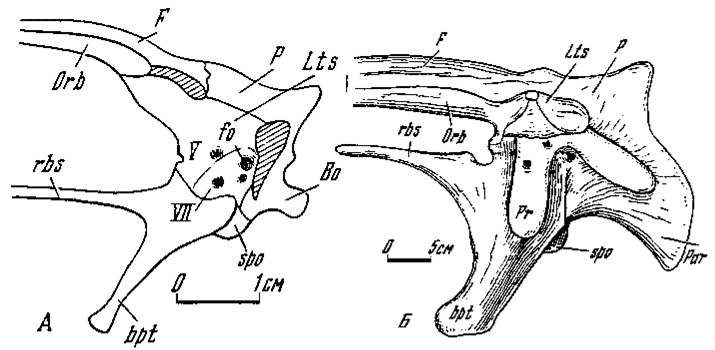
Рис.3. Черепно-мозговая коробка (слева, схематизировано): А - Velociraptor mongoliensis; Б - Tarbosaurus bataar
Передняя часть нейрокрания. Рассмотрение этой части достаточно ограничить преимущественно латеросфеноидами и орбитосфеноидами, по сравнению с другими составляющими, более подверженными модификациям у разных групп хищных динозавров. Латеросфеноиды присутствуют у всех изученных групп. Зачастую они занимают немалую долю в верхней части боковой стенки нейрокрания (рис. 3), отдают орбитальные крылья в вентральной части переднего отдела черепа, а также входят в окружение области выхода зрительного нерва. В боковой стенке латеросфеноиды на значительном участке соединяются с теменными костями. Эта граница носит чаще подвижный характер и принадлежит к числу критических в явлении метакинетизма у хищных динозавров. Соединение же орбитальных крыльев латеросфеноидов с заднеглазничными и лобными костями обычно жесткое. У ряда групп латеросфеноиды налегают на проотическую часть спинки гипофизной ямки. Латеросфеноиды нередко принимали непосредственное участие в формировании отверстия для выхода зрительных нервов. Иногда они в значительной мере окружали этот выход, что имеет место у заурорнитоидид и дромеозаврид. Латеросфеноиды у тираннозаврид близко подступают к зрительному отверстию, но непосредственного участия в его формировании, возможно, и не принимают, так как оттесняются здесь орбитосфеноидами. Аналогичное наблюдается и у аллозаврид (Madsen, 1976a). Латеросфеноиды могут протягиваться значительно вперед от уровня отхождения их орбитальных крыльев и участвовать в формировании вентральной поверхности переднего отдела нейрокрания. В этом случае они граничат с орбитальными частями лобных костей, что наблюдается, к примеру, у овирапторид.
Положение орбитосфеноидов довольно изменчиво. По-видимому, они могут испытывать редукцию, иногда до полного исчезновения. Вероятно, так обстоит дело у заурорнитоидид и дромеозаврид, тогда как редуцированные орбитосфеноиды у орнитомимид занимают ограниченный участок лишь в нижнем краю зрительного отверстия. У тираннозаврид, как отмечалось, орбитосфеноиды полностью окружают это отверстие и протягиваются вперед, формируя короткий, узкий срединный участок вентральной поверхности переднего отдела нейрокрания между орбитальными краями лобных костей. У овирапторид орбитосфеноиды находятся в сходном положении. Изнутри они образуют трубчатый зрительный канал, обособленный от выше расположенного прохода обонятельных нервов.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'