
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
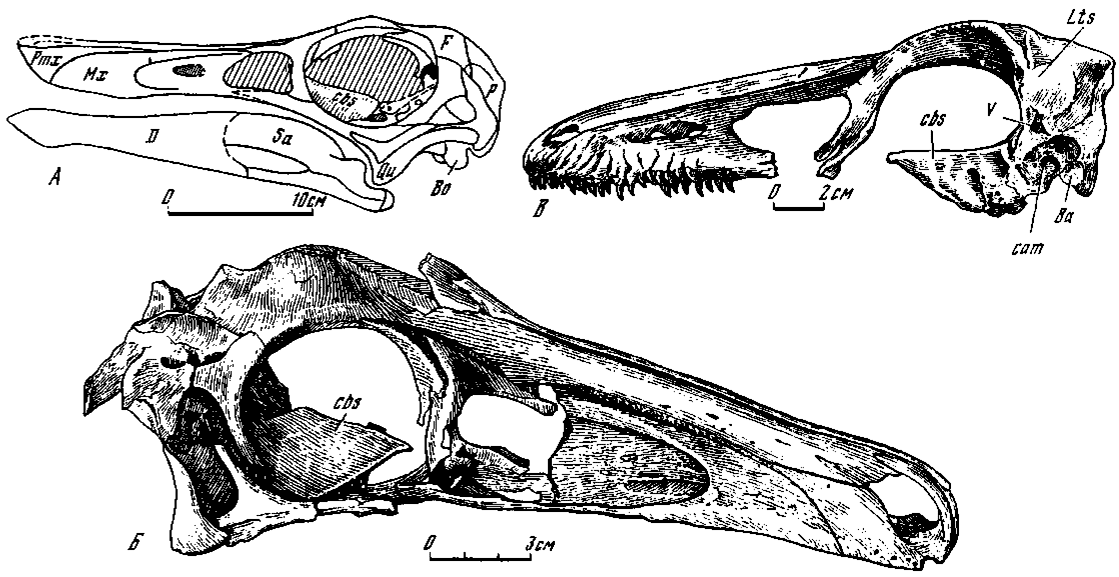
Основание черепа
Специфические черты базикрания у хищных динозавров позволяют выделять по крайней мере три его типа, главным образом, на основе степени развития базисфеноидной капсулы.
Первый тип включает нейрокраний с бескапсульным основанием. Он в общем считался свойственным хищным динозаврам и был с большими или меньшими подробностями известен у тираннозаврид и дромеозаврид, а также у более ранних аллозаврид и, возможно, целурид. Ко второму типу относится череп с капсульным основанием, строение которого характеризуется присутствием специфической базисфеноидной капсулы, впервые описанной у монгольских орнитомимид и заурорнитоидид (Osmolska, Roniewicz, Baisbold, 1972; Barsbold, 1974). Третий тип, условно называемый промежуточным (между первым и вторым), не имеет полностью оформленной капсулы, однако базикраний в этом случае отличался значительным объемным расширением, в том числе и в передней части, где у капсульного типа располагалась базисфеноидная капсула. Хотя формирования капсулы не произошло, передняя часть базикрания кажется внешне почти достигшей капсульной структуры. Внутри в базикраний развита система синусов. Переходный тип обнаружен у эрликозавра (Barsbold, Perle, 1980).
Рассмотрение специфических черт нейрокрания целесообразно начать с простейшего бескапсульного типа.
Бескапсульный тип (рис. 3). Отсутствие капсулы в немалой мере отражается на конфигурации базикрания. Прежде всего, он не получает столь заметного объемного расширения; вместе с тем, базиптеригоидные отростки и сфеноокципитальные бугры выступают значительно сильнее, чем в других типах. Наиболее ярко это проявляется у тираннозаврид и аллозаврид, основание базисфеноида у которых сильно выступает вниз и вперед, отдавая утолщенные и удлиненные базиптеригоидные отростки с хорошо развитыми суставными концами. Сзади к базисфеноиду присоединяется основная затылочная кость, формирующая окципитальную часть широких массивных сфеноокципитальных бугров. Cristae ventrolateralis et tuberalis чаще всего значительно развиты. Сходным строением обладает базикраний у монгольских дромеозаврид, а также у итемируса (Курзанов, 1976а) и аллозаврид (Madsen, 1976a). Увеличение высоты базикрания и всего черепа связано с вертикальной протяженностью базисфеноида. Относительно наибольшая высота черепа наблюдается в группах гигантских хищников с максимальной вертикальной протяженностью базисфеноида. Это прежде всего тираннозавриды и аллозавриды, после них идут "высокоголовые" дромеозавриды (типа Dromaeosaurus), наконец, самое последнее место в убывающем по высоте черепа ряду будет принадлежать "узкоголовым" дромеозавридам (типа Velociraptor).
Базикраний у овирапторид заметно выходит за рамки рассматриваемого типа. Он сформирован в виде компактной платформы, удлиненной в передне-заднем направлении и несколько сжатой с боков, что нехарактерно для бескапсульного базикрания. Наибольшее же отклонение состоит в необычно сильной редукции базиптеригоидных отростков, которые почти не обособлены от основания базисфеноида. Птеригоиды прилегают (и, возможно, срастаются) к едва обозначенным базиптеригоидным отросткам. Такой характер сочленения должен был полностью блокировать или же сильно ограничивать его подвижность. По степени редукции базиптеригоидных отростков и отсутствию значительного вентрального протяжения базикраний у овирапторид имеет сходство с промежуточным типом. Однако отсутствие сильного объема расширения базикрания, рассматриваемого в качестве решающего признака "переходности", заставляет относить овирапторидный базикраний к бескапсульному типу.
Базисфеноидный ростр в бескапсульном типе в виде тонкого горизонтального стержня протягивается вперед и выше межптеригоидной ямы. У тираннозаврид и аллозаврид ростр по вертикали значительно отстоит от основания черепа и приближен к переднему отделу нейрокрания. Напротив, дромеозавриды характеризуются более низко расположенным ростром, и монгольский велоцираптор в этом плане, пожалуй, занимает наиболее крайнее место. Сравнительно низкое положение ростра свойственно также овирапторидам. При этом у них основание ростра сильно уплощено и расширено в сагиттальной плоскости, однако без какого-либо объемного расширения.
Бескапсульный базикраний, скорее всего, развит при подвижном базиптеригоидном сочленении, необходимом для широко распространенного в черепе хищных динозавров метакинетизма. Такая специфическая особенность бескапсульного основания черепа, как высокая степень развития базиптеригоидных отростков, выраженная в их удлинении, массивности, разработанности суставных концов и т.д., иллюстрирует значительную подвижность в данном сочленении. Редукция базиптеригоидных отростков у овирапторид предположительно связана с блокировкой (или же слабой подвижностью) одноименного сочленения. Среднее ухо в черепе данного типа не имело оформленной костной плоскости и носило вполне типичный рептилийный характер.
Капсульный тип (рис. 4). Капсульное строение базикрания было впервые установлено у монгольских орнитомимид и заурорнитоидид (Osmolska, Roniewicz, Barsbold,: 1972; Barsbold, 1974). При этом возможно выделение двух типов структур: в первом - базисфеноидная капсула открыта сзади (орнитомимозавры); во втором подтипе капсула не имеет наружного отверстия (заурорнитоидиды).
В обоих случаях базисфеноидная капсула представляет собой грушевидное полое образование в основании мозговой коробки черепа, направленное вперед своим суженным концом. Задняя, наиболее расширенная, область капсулы проявляет специфические особенности в каждом из подтипов. Дорзально вдоль срединной линии капсулы протягивался узкий желоб, переходивший на базисфеноидный ростр и отмечавший основание межглазничной перегородки.
В орнитомимидном подтипе базисфеноидная капсула своей наиболее расширенной задней частью переходила в короткое и высокое преддверие, открывавшееся наружу назад крупным отверстием, сжатым сверху и расширявшимся книзу. Дно и боковые стенки преддверия образованы вентральной частью базикрания, при этом нижний край отверстия ограничивался базиптеригоидными отростками, с которыми соединялись нисходившие от сфеноокципитальных бугров парные ребра сложного поперечного сечения, обрамлявшие наружное отверстие с боков. Дорзальный конец каждого из ребер находился немного впереди базиокципитальной части сфеноокципитальных бугров, отдавая назад к ним выпуклый вентролатеральный гребень (подобный гребень известен у ящериц - Save-Saderberg, 1947), участвовавший в формировании бугра. Медиальная поверхность стенки преддверия вдоль края внутреннего отверстия, которым соединялась собственно капсула с преддверием несла аналогичный по форме гребень, соединявшийся с вентролатеральным поперечной перемычкой. Вследствие развития крупного наружного заднего отверстия базиптеригоидные отростки, ограничивавшие его снизу, располагались значительно ниже сфеноокципитальных бугров, и основание базикрания в этой части имело перерыв.
Заурорнитоидидный подтип капсулы отличался, прежде всего, отсутствием наружного заднего отверстия. Вследствие этого вентральная часть базикрания здесь сформирована без всякого перерыва. Базисфеноидная капсула предположительно имела связь с системой синусов, развитых внутри вентральной части базикрания. Кроме того, капсула у заурорнитоидид связана отверстием в задней ее стенке или же синусами с полостью среднего уха. Таким образом, оформленного преддверия с наружным отверстием (как у орнитомимид) в базисфеноидной капсуле заурорнитоидидного подтипа не было. Базиптеригоидные отростки и сфеноокципитальные бугры находились на одном уровне. Вентролатеральный гребень базикрания не получил развития.
Переходный тип (рис. 5, 6). В основании черепа у эрликозавра настоящая полная грушевидная базисфеноидная капсула не получила развития, однако данная область была объемно сильно расширенной. Это позволяет видеть в ней как бы переходную к оформленной капсуле стадию. Расширение затрагивало не только переднюю часть базикрания, но и в большей мере заднюю, структурно связанную с полостью среднего уха, что характерно и для заурорнитоидид. Передняя часть развита в виде выпуклого основания переднего отростка базисфеноида, слабо обособленного от остальной части базикрания. Вдоль отростка дорзально протягивался его желобчатый край, принимавший основание межглазничной перегородки. Вентральная поверхность без перерыва продолжалась назад от базиптеригоидных отростков к сфеноокципитальным буграм. Указанные структуры, как и у заурорнитоидид, находились на одном горизонтальном уровне.
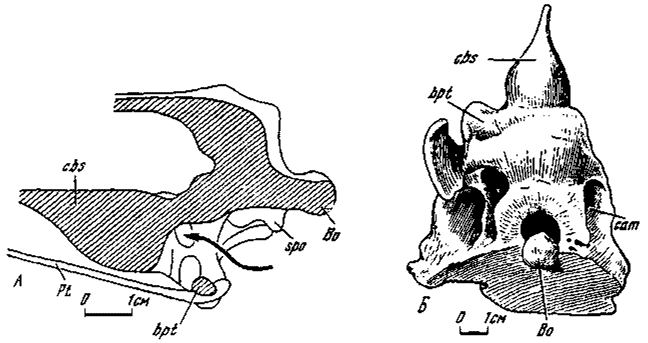
Рис. 5. Базисфеноидная капсула: А - Garudimimus brevipes (сагиттальный разрез, схематизировано, правая половина); Б - Saurornithoides junior (№ 100/1,снизу и сзади)
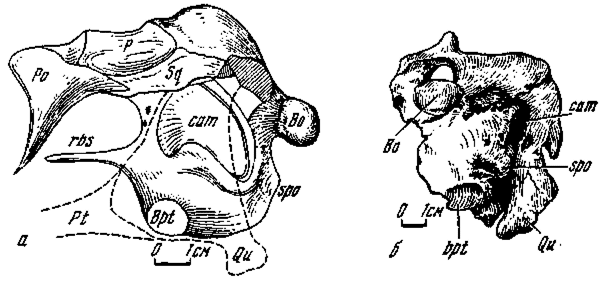
Рис. 6. Черепно-мозговая коробка Erlikosaurus andrewsi (№ 100/111): а - слева (схематизировано); б - сзади
Внутренняя полость в базикрании представлена системой синусов. Это означает, что базисфеноидная капсула связана с вентральной системой синусов в основании мозговой коробки черепа. Следует отметить, что в сравнении с расширенным базикранием у эрликозавра, закапсульная часть основания черепа у орнитомимид и заурорнитоидид сильно редуцирована; следовательно, синусная система у последних не была столь обширной, как будто капсула в немалой мере заместила ее.
Описанное выше строение объемного базикрания в определенной мере отражает становление базисфеноидной капсулы "закрытого" типа, т.е. без заднего наружного отверстия. Ряд характерных черт, кажется, дает основание такому заключению. Во-первых, топографические и структурные соотношения базисфеноидного ростра у капсулы обоих типов и объемного базикрания вполне идентичны. Во-вторых, капсула без заднего отверстия связана с появлением полностью оформленной костной полости среднего уха, отмеченной впервые у заурорнитоидид (Barsbold, 1974), а также обнаруженной у эрликозавра (Barsbold, Perle, 1980). Подобная полость отсутствовала при капсуле с наружным отверстием у орнитомимид.
Базисфеноидная капсула при первом описании предположительно связывалась с карманом Ратке (Osmolska, Roniewicz, Barsbold, 1972). Однако внутренняя полость капсулы слишком велика, чтобы соответствовать этому чисто эмбриональному образованию, каким является карман Ратке. Выше отмечалось, что объемно увеличенный, с развитой системой синусов, но бескапсульный базикраний у эрликозавра наводит на мысль о развитии базисфеноидной капсулы в связи с пневматизацией базисфеноида. Хорошо известны системы воздухоносных синусов основания черепа у крокодилов и птиц; у первых воздухоносные каналы связаны с евстахиевыми трубами. В основании черепа у протозуха также отмечены очень крупные воздухоносные синусы (Walker, 1972). Допустимо предположение, что капсульный базисфеноид у двух групп теропод - орнитомимозавров и заурорнитоидид - связан с дальнейшей пневматизацией черепа, широко распространенной в архозавровых ветвях.
Сонные каналы. Развитие сонных каналов впервые было отмечено у дромеозаврид и итемируса (Курзанов, 1976; Colbert, Russell, 1969), у которых на боковой стороне основания базиптеригоидных отростков обнаружены входные отверстия внутренних сонных артерий. У итемируса на пути к гипофизной ямке сонные артерии проходят через синусы в теле базисфеноида. Возможно, в сходном положении находились входные отверстия сонных каналов и у аллозаврид (Madsen, 1976b). У тираннозаврид и овирапторид эти отверстия наблюдать не удалось. При отмеченном положении сонных каналов небные ветви внутренних сонных артерий с одноименными ветвями лицевого нерва проходили под основанием черепа поверхностно, что показано у итемируса (Курзанов, 1976а) и предполагается у дромеозаврид. В этом случае небные ветви сонной артерии и лицевого нерва проходили под основанием базиптеригоидных отростков и протягивались далее вперед.
Парабазальные каналы. Парабазальные каналы удалось обнаружить у орнитомимозавров, сегнозавров и заурорнитоидид (рис. 7). Наиболее полные представления об их структуре были получены при изучении примитивной ветви орнитомимозавров - гарудимимид, восполнявших в этом плаке существовавший до сих пор пробел.
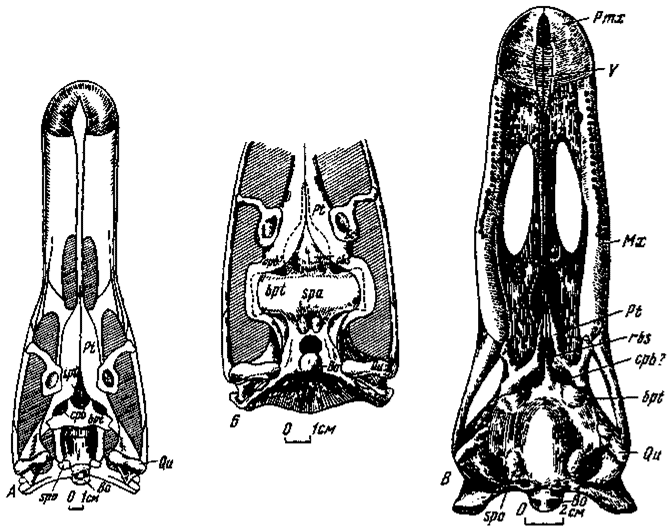
Рис. 7. Вентральная часть базикрания и небный комплекс: А - Garudimimus brevipes (№ 100/13); Б - Saurornithoides junior (№ 100/1; птеригоиды эктоптеригоиды и базиптеригоидное сочленение реконструированы, передняя часть небного комплекса не показана); В - Erlikosaurus andrewsi (№ 100/111)
Вход в парабазальный канал у гарудимимуса представлен отверстием сразу впереди укороченного, поперечно направленного базиптеригоидного отростка вблизи задне-нижнего края капсулы, примерно в области перехода к вентральной поверхности базикрания. Ранее отмечалось, что в бескапсульном типе базикрания этот переход не имеет перерыва.
Внешне в рассматриваемой области не наблюдается швов между базисфеноидом и парасфеноидом, прикрывающим его снизу. Птеригоиды, надвинутые к передне-боковым краям входных отверстий, по-видимому, лишь подстилают базикраний и не принимают участие в формировании каналов, проходящих между базисфеноидом и парасфеноидом. Входные отверстия каналов довольно крупные, овальной формы, длинная ось отверстий направлена под углом к сагиттальной плоскости. Отверстия вели в короткие вертикальные каналы, протягивавшиеся в области переходов задних вентролатеральных краев базисфеноидной капсулы к закапсульной части базикрания. Снаружи к стенке прилегает передний участок квадратного фланга птеригоида, при отхождении нижнего края которой от капсулы образуется щелевидный проход, подстилавшийся основным телом птеригоида и, вероятно, пропускавший вперед небную артерию и ветвь лицевого нерва.
В рассматриваемом типе парабазальных каналов немалое значение имеет блокировка базиптеригоидного соединения, сопряженная с редукцией базиптеригоидных отростков, дистальные концы которых утрачивают суставной характер. Кроме того, развитие птеригоидов, подстилающих вентральную поверхность базикрания в области входных отверстий, возможно, также служит указанием ограничения подвижности базиптеригоидного сустава. Вполне отчетлива редукция межптеригоидной ямы.
Тождественность в строении базикраниальной области у орнитомимозавров позволяет принимать сходный тип в структуре парабазальных каналов и у прогрессивных орнитомимид, соответствующая часть черепа которых в коллекции сохранилась лишь частично. У галлимимуса имеются поперечно направленные укороченные базиптеригоидные отростки, с которыми сливается вентральная часть базисфеноидной капсулы, и в области перехода капсульной и закапсульной частей базикрания находятся сходно построенные парабазальные каналы. Лишь разрушенность рассматриваемых участков лишала до сих пор возможности правильной трактовки данных структур у галлимимуса (Osmolska, Roniewicz, Barsbold, 1972). И в этом случае наблюдается блокировка базиптеригоидного соединения.
У заурорнитоидид также имеются поперечные базиптеригоидные отростки, отличающиеся лишь относительной удлиненностью и шириной, с которыми спереди смыкается вентральная часть базисфеноидной капсулы. Область перехода капсулы к закапсульной части базикрания в общем подобна орнитомимидной, и это дает основание предполагать сходство и в строении парабазальных каналов. Вероятно, сходным было также положение птеригоидов вблизи нижне-боковой части капсулы, что позволяет допускать аналогичное прохождение вперед небной артерии и ветви лицевого нерва.
Во многом близкое сходство в строении парабазальных каналов отмечено и у сегнозаврид. У эрликозавра базисфеноидная капсула отсутствует, базиптеригоидные отростки сильно редуцированы и как бы вытесняются значительным объемным расширением базикрания. Вход в парабазальный канал в отличие от орнитомимидного представлен щелевидным отверстием, ограниченным сзади редуцированным базиптеригоидным отростком. Птеригоиды, также надвинутые к переднебоковым краям входных отверстий, по-видимому, подстилают базикраний и не принимают прямого участия в образовании каналов. Как и при капсульном типе, входные отверстия вели в короткие вертикальные каналы, протягивавшиеся в передне-боковой области базикрания, к которой снаружи прилегают начальные участки квадратных флангов птеригоидов. Отходя нижне-передней частью от базикрания, они формируют щелевидное отверстие, пропускавшее вперед небную артерию и ветвь лицевого нерва. Блокировка базиптеригоидного соединения, возможно, достигает наибольшей степени не только в силу максимальной редукции базиптеригоидных отростков, но также появления дополнительного соединения гипертрофированных сошников, рассматриваемых ниже. Межптеригоидная яма значительно редуцирована.
Примеры строения парабазальных каналов у разных групп хищных динозавров позволяют рассмотреть общие условия их развития. Во-первых, парабазальные каналы данной структуры в трех случаях из четырех связаны с развитием базисфеноидной капсулы (у гарудимимид, орнитомимид, заурорнитоидид). У сегнозаврид (эрликозавра) полностью оформленной базисфеноидной капсулы не было, однако базикраний объемно сильно увеличен. Парабазальные каналы находились в области перехода между капсульной и бескапсульной частями базикрания и при отсутствии внешне наблюдаемых швов протягивались между базисфеноидом и прикрывающим его снизу парасфеноидом. У эрликозавра при отсутствии капсулы каналы приурочивались к передне-боковым частям объемно расширенного базикрания. Межптеригоидная яма сильно редуцирована. Во-вторых, базиптеригоидные отростки были всегда направлены поперечно и в большинстве случаев редуцированы в разной степени - наиболее сильно у сегнозаврид, в меньшей мере - у заурорнитоидид. В-третьих, имеет место блокировка базиптеригоидного соединения, вызванная чаще всего редукцией базиптеригоидных отростков, дистальные окончания которых утрачивают суставной характер, а также появлением дополнительного соединения птеригоидов (и сошников у эрликозавра) с базисфеноидом, способствовавшего блокировке. Таким образом, парабазальные каналы у хищных - динозавров, аналогично видиевым каналам у териодонтов и черепах (Татаринов, 1965, 1966, 1976; Camp, Welles, 1956), формируются в условиях блокированного базиптеригоидного соединения.
Характер строения парабазальных каналов у теропод, по-видимому, снимает вопрос о видиевых каналах, признававшихся до сих пор, по крайней мере, у некоторых групп динозавров (Рождественский, 1968; Ostrom, 1961). Недавно было высказано сомнение в правильности их отождествления у гадрозавров и показано развитие сонных каналов у итемируса (Курзанов, 1976а). Сейчас становится ясным, что предположения о путях прохождения небной артерии и одноименной ветви лицевого нерва, допускавшие присутствие видиевых каналов у орнитомимид и заурорнитоидид (Osmolska, Roniewicz, Barsbold, 1972; Barsbold, 1974), не имели под собой основы и были вызваны к жизни главным образом сохранностью существовавшего в то время черепного материала.
Ушная область эндокрания. Строение эндокрания вблизи ушной капсулы впервые было подробно показано на примере хорошо сохранившейся мозговой коробки у итемируса (Курзанов, 1976а). При этом ряд черт был признан специфичным только лишь для узкой группы, в которую входит эта форма. Однако при рассмотрении околоушной области у других хищных динозавров сразу же выясняется общий характер этих особенностей. В этом плане изучались экземпляры мозговой коробки у овирапторид, эрликозавра, а также неопределенного карнозавра из Аргалант (отдельно найденная мозговая коробка) (рис. 8).
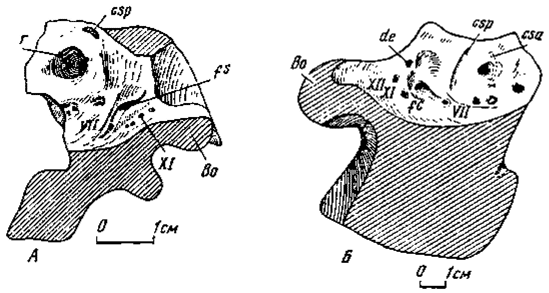
Рис.8. Ушная область эндокрания: А - Ingenia yanshini Barsbold (№100/30, сагиттальный разрез, правая половина); Б - неопределенный карнозавр из Джаргалант (№100/200, сагиттальный разрез,левая половина)
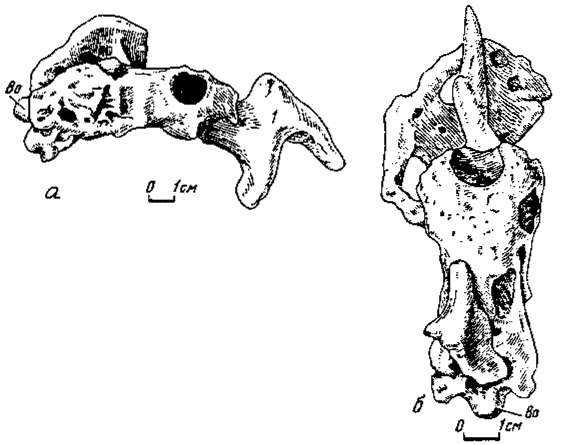
Рис.9. Форма мозга Ingenia yanshini (№100/30): а - справа: б - сверху
Мозговая полость в ушной области у перечисленных хищных динозавров становилась особенно высокой из-за значительной вогнутости дна эндокрания, увеличивавшей ее объем, возможно, в связи с сильным развитием мозжечка, что первоначально отмечалось на примере итемируса (Курзанов, 1976а). Боковая стенка полости во всех рассматриваемых случаях несла глубокий и обширный карман, вытянутый примерно в направлении парокципитального отростка. Карман имел гладкие стенки, и мере углубления становился более узким, но слегка расширялся на дне. Он вдавался в ушную капсулу между полукружными каналами, охватывавшими его, но прямо связи с ней не обнаруживал. Крупное устье кармана несколько сжато с боков. Карман в подобном положении впервые отмечался в эндокрании у итемируса и был назван внутрислуховым (Курзанов, 1976а).
Немного позади устья кармана и ближе к его нижней части находится довольно крупный выступ, связанный с объемным расширением в данной области лабиринт внутреннего уха. Развитие противонаправленных выступов на боковых стенках мозговой полости значительно сужало ее верхнюю часть сразу выше слуховых капсул Передний полукружный канал огибал по дуге карман спереди и сверху, сзади же бы; ограничен нисходившим до уровня вершины выступа crus communis. Полукружный канал не наблюдается. Горизонтальный полукружный канал должен был протягиваться сразу под карманом. Положение полукружных каналов дает основание предполагать, что объемное расширение ушной капсулы, фиксируемое упоминавшимся выше выступом эндокрания, соответствовало полости преддверия костного лабиринта. Эндолимфатический проток, по-видимому, оттеснен карманом к краю большого затылочного отверстия, как предполагалось у итемируса. Прохождение перилимфатического протока в рассматриваемом случае не выявлено.
Ниже кармана расположено несколько отверстий. У овирапторид в передней группе из трех отверстий самое нижнее и более крупное сквозное представляло выход для лицевого нерва. Два более верхних и мелких, возможно, проводили сосуды. Более задняя группа, расположенная ближе к нижней части преддверия, фиксируемого выступом, также состоит из трех отверстий, предположительно, впускавших в ушную капсулу ветви слухового нерва.
Ниже и сзади выступа, маркировавшего полость преддверия, косо расположено желобообразное углубление, в верхнем заднем конце которого находится довольно крупное яремное отверстие, пропускавшее IX-XI нервы и заднюю мозговую вену. Немного выше дна эндокраниальной полости расположен продольный ряд из четырех отверстий, каудально увеличивавшихся и пропускавших ветви XII нерва, снаружи выходивших через отверстие латеральнее затылочного мыщелка.
Расположение отверстий для выхода черепно-мозговых нервов в рассматриваемой области у неопределенного карнозавра из Аргалант и эрликозавра во многом сходно отмеченному выше. У карнозавра ниже и впереди кармана находилось крупное отверстие для выхода тройничного нерва. У сегнозавра же отверстие для прохождения тройничного нерва в толще кости ведет в три канала, раздельно открывавшихся снаружи. Сзади и выше него расположены два меньших отверстия, из них переднее сквозное пропускало лицевой нерв, заднее - слуховой. Крупное яремное отверстие находилось в углублении сзади эндокраниального выступа, фиксировавшего область преддверия. Выход XII нерва вполне аналогичен отмеченному на примере овирапторид.
Ушная область эндокрания у представителей четырех групп хищных динозавров - итемируса (Курзанов, 1976а) и, как показано, у овирапторид, эрликозавра и карнозавра из Аргалант, обладала принципиально единым планом строения. При первоначальном описании итемируса обширный карман в костном пространстве между полукружными каналами связывался с венозной системой, с существованием в нем венозного синуса. Вместе с тем, допускалось и флоккулярное происхождение кармана. Однако флоккули получают развитие только у млекопитающих, возможное исключение составляют териодонты. У рептилий же намечены только боковые доли мозжечка. Нам кажется более вероятным допущение мозжечкового происхождения карманов (за счет боковых долей, а не флоккулей мозжечка), чем присутствие в нем венозного резервуара. Следует отметить, что стенки кармана вполне гладкие в отличие от стенок кармана у итемируса, что привлекалось в качестве одного из доводов в пользу существования какого-то (скорее всего, венозного) жидкостного мешка в кармане. У одного из териодонтов (Татаринов, 1977) флоккулярная яма находится в близком рассмотренному положении. Еще раньше, до публикации итемирусового материала, в эндокрании у американского дромеозаврида (Dromaeosaurus) была отмечена ямка, вполне тождественная по положению среди полукружных каналов рассмотренной выше и связывавшаяся или с обособлением выступов мозжечка, или с sacculus внутреннего уха (Colbert, Russell. 1969). Последнее, как стало ясно сейчас, отпадает полностью, и тем самым предполагаемый мозжечковый карман отмечается еще у одного представителя хищных динозавров. Положение отверстий дромеозавра, пропускавших в данной области ряд черепно-мозговых нервов, также вполне тождественно рассмотренному.
Строение ушной области эндокрания у ряда различных хищных динозавров совершенно ясно свидетельствует в пользу постоянства ее главных черт. Оно показывает, что мозговая коробка действительно наиболее консервативная часть черепа, в остальном существенно изменявшегося. Следовательно, особенности этой области, считавшиеся уникальными для итемируса (Курзанов, 1976а), возведенного на этом основании в ранг семейства, носят гораздо более общий характер*.
* (Сходный карман отмечается еще у одной группы теропод - авимимид и более не связывается с развитием жидкостного мешка (Курзанов, 1981).)
Форма головного мозга. На единственном примере у овирапторид удалось наблюдать редкий случай сохранения внутренней пластинки твердой оболочки головного мозга (рис. 9). Это показало в общих чертах величину эпидюрального пространства и позволило судить об объеме, занятом мозгом. Поверхность внутренней пластинки ясно фиксирует положение и форму больших полушарий, сильно расширенных и выпуклых по отношению к позади расположенным частям мозга. Сами полушарии занимают несколько меньше трети всей длины пространства, занятого мозгом. Дорзально между полушариями намечается широкое пологое углубление, отвечавшее переднему участку верхней сагиттальной пазухи. Внутренняя пластинка прободена крупными округлыми отверстиями, соответствовавшими зрительным долям и основанию обонятельного тракта. При этом отмечается несимметричное расположение отверстий, отвечающих зрительным долям: левое обращено вбок и вниз, тогда как правое - вбок и назад. Дорзальная поверхность внутренней пластинки с шероховатостью, видимо, отражающей развитие синусов эпидюрального пространства. Судя по отдельным костям крыши черепа, само пространство над мозгом было значительным, лишь немногим уступая объему, занятому мозгом.
Костная полость среднего уха. Полость среднего уха с костными стенками впервые была обнаружена у заурорнитоидеса и названа описательно "латеральной депрессией" (Barsbold, 1974). Позднее подобная полость была найдена в черепе у эрликозавра (Barsbold, Perle, 1980). Полость среднего уха у заурорнитоидеса в виде объемистой депрессии, открытой снаружи, занимает значительное пространство в вентро-латеральной части мозгового черепа сзади базисфеноидной капсулы между базиптеригоидными и основанием парокципитальных отростков (рис. 10). В строении полости участвовали различные составляющие черепа, соотношения которых выявляются лишь в общих чертах. Передняя часть полости составлена преимущественно базисфеноидом, ее верхний край - переднеушной костью, задний край - основанием парокципитального отростка, нижний - боковой и основной затылочными костями. Медиальная стенка полости несет пересекавшиеся гребни, по-видимому, связанные с развитием многочисленных воздухоносных синусов; они особенно хорошо видны у эрликозавра. Через синусы шло соединение передней части полости с базисфеноидной капсулой. Намечается такой же проход через синусы медиальной стенки полости, направленный вверх и внутрь, к гипофизной ямке. Предполагается, что через синусы передней части базисфеноида проходили каналы для внутренних сонных артерий. Сразу под верхним краем полости расположено удлиненное отверстие, отождествляемое с выходом для VII нерва. У птиц в сопоставимом положении может находиться также отверстие, соединявшее полость среднего уха с воздушными полостями внутри черепа. Снаружи и выше края полости расположено отверстие для выхода тройничного нерва. Самое заднее окончание полости занято крупным отверстием, соответствовавшим овальному окну, снаружи, возможно, объединенного с круглым, от которого по передней стенке парокципитального отростка протягивается неглубокий стапедиальный желоб. Ниже и впереди овального окна, ближе к нижнему краю полости и в значительной мере ограничиваясь им, расположены крупные, хорошо оформленные отверстия, которыми синусы открывались в полость среднего уха.
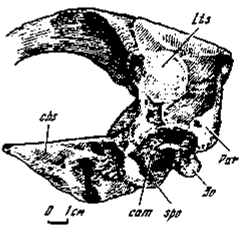
Рис.10. Полость среднего уха Saurornithoides junior (№100/1 слева)
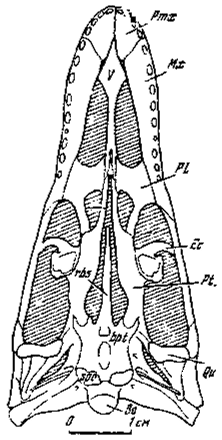
Рис.11. Череп Dromaeosaurus albertensis Matthew et Brown cнизу (Colbert, Russel, 1969)
Полость среднего уха у эрликозавра (рис. 6) во многом идентична рассмотренной у заурорнитоидеса. Вместе с тем сохранность некоторых ее существенных элементов позволяет более полно представить модификации в строении оформленной костной полости среднего уха у хищных динозавров. Наибольшее внешнее различие состоит в значительном объемном расширении ушной области и основания мозгового черепа, что не выражено, во всяком случае, в такой степени, у заурорнитоидеса. При этом боковые стенки мозгового черепа особенно в нижней части сильно раздаются вбок и становятся выпуклыми. Столь же сильно раздается и область сфеноокципитальных бугров, которые значительно смещаются латерально. Таким образом, рассматриваемая часть базикрания у эрликозавра как бы заполняет все пространство между базиптеригоидными и парокципитальными отростками. Медиальная стенка полости пронизана синусами, что отмечено и у заурорнитоидеса. Синусы затрудняют опознание отверстий, через которые в базисфеноиде проходили каналы для внутренних сонных артерий.
По объему полость эрликозавра не уступает заурорнитоидесной, но края полости в большей степени сближены друг с другом, что делает ее более закрытой, чем у заурорнитоидеса. Далее, края полости у эрликозавра сложены преимущественно базисфеноидом. Снаружи отверстие полости плотно прикрывалось участком квадрато-птеригоидного соединения (оно не сохранилось у заурорнитоидеса). Таким образом, наружная крыша полости почти на всем протяжении имела двойную стенку из-за соединения здесь медиального отростка квадратной кости и квадратного фланга птеригоида. Открывалась же полость лишь сзади ствола квадратной кости удлиненно-овальной ушной вырезкой, приблизительно отмечавшей положение наружного уха. То же самое предполагается и у заурорнитоидеса. В этом плане проступают определенно рептилийные черты в строении полости, хотя высокая степень костного оформления в обоих рассмотренных случаях делает ее более сходной с птичьим средним ухом.
Таким образом, строение полости среднего уха у заурорнитоидеса и эрликозавра вполне единообразно. Кроме отмеченных отличий следует выделить лишь преимущественное участие базисфеноида в строении полости у последнего. В этом отношении полость отличается от птичьей, сформированной главным образом периотикой и боковыми затылочными костями. Следует отметить также закрытость большей части отверстия полости среднего уха квадрато-птеригоидным соединением, причем обе названные кости срастались с его краями. Развитие ушной вырезки сзади ствола квадратной кости свидетельствует о присутствии барабанной полости рептилийного типа.
Среднее ухо без костных стенок. Среднее ухо без костных стенок встречается у разных групп хищных динозавров, судя по всему, оно не имеет прямой связи с присутствием капсульного базикрания, как казалось ранее при первом описании такой ушной полости (Barsbold, 1974). Это хорошо иллюстрируется наличием полости среднего уха без костных стенок у орнитомимид и гарудимимид, обладающих базисфеноидной капсулой. Среднее ухо без оформленной костями полости известно также у тираннозаврид, дромеозаврид и овирапторид, характеризовавшихся бескапсульным базикранием. У всех них полость среднего уха была построена по типу современных рептилий (Janensch, 1935; Wyeth, 1924). На ископаемом материале положение среднего уха подобного типа фиксируется овальным окном, от которого по передне-нижнему краю парокципитальных отростков протягивается ясно обозначенный стапедиальный желоб, вмещавший слуховую косточку. По направлению желоба можно допустить, что наружное ухо находилось сразу за задним краем квадратной кости немного ниже ее верхнего суставного конца, как и в случае развития костных стенок полости у некоторых групп хищных динозавров.
Заключение. Теперь становятся более ясными некоторые корреляции в строении основания мозгового черепа у хищных динозавров. По всей вероятности, базисфеноидная капсула действительно имеет "сфеноидное" происхождение и представляет собой полое вздутие, связанное с развитием синусной системы базикрания. Однако функциональное значение капсулы во многом неясно.
Развитие полностью оформленной базисфеноидной капсулы, а также объемное расширение передней части базисфеноида, еще не превратившейся в настоящую капсулу, возможно, обусловливает присутствие парабазальных каналов; как отмечалось, их развитие связано с блокировкой базиптеригоидного сочленения, и в этом плане можно провести параллель с черепахами и териодонтами. Отсутствие капсулы (или объемного расширения передней части базикрания), подвижность базиптеригоидного сочленения, т.е. черты, наиболее ярко выраженные у дромеозаврид и тираннозаврид, вполне коррелируются с отсутствием парабазальных каналов. Оформленная костная полость среднего уха птичьего типа имелась у различных групп хищных динозавров. У всех была развита базисфеноидная капсула или же присутствовало объемное расширение передней части базикрания. У орнитомимозавров при хорошей капсуле костная полость не развита. Во внутреннем ухе так называемый "внутрислуховой карман" имел всеобщее распространение и, видимо, был связан с боковыми долями мозжечка.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'