
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Нижняя челюсть
Построенная по рептилийному образцу нижняя челюсть у хищных динозавров разделяется на две основные модификации - зубатую и беззубую, из которых последняя свойственна лишь трем группам - орнитомимозаврам, овирапторидам и ценагнатидам (рис. 13, 14). У сегнозавров нижняя челюсть лишена зубов в самом переднем окончании, в остальном же она полностью соответствует нормальному зубатому типу. Несмотря на немалые различия, беззубая нижняя челюсть у орнитомимид и гарудимимид принципиально также близка к зубатой, тогда как у овирапторид и ценагнатид она представляет собой гораздо более существенное отклонение. Поэтому целесообразно объединять орнитомимозавровую нижнюю челюсть с зубатым (нормальным) типом и выделять особо овирапторидный, включающий и нижнюю челюсть ценагнатид, рассматриваемую отдельно. При описании удобно придерживаться двух структурных подразделений нижней челюсти - переднего и заднего.
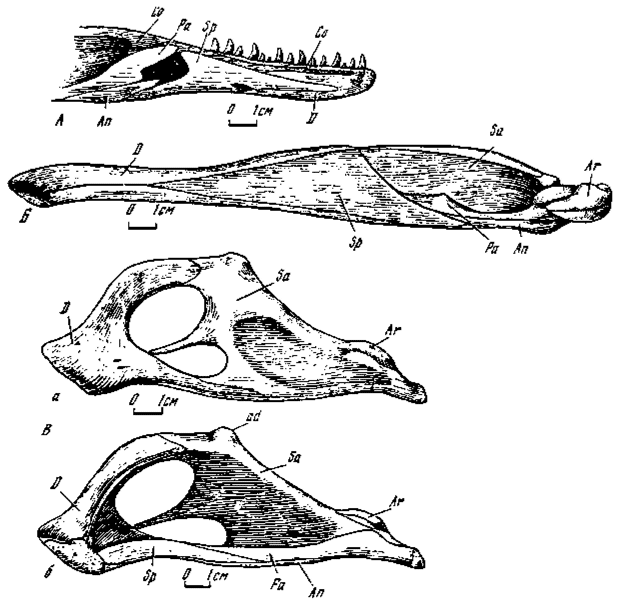
Рис.13. Нижняя челюсть: А - Velociraptor mongoliensis (№100/24, передний отдел левой ветви изнутри); Б - Garudimimus brevipes (№100/13, правая ветвь изнутри); В - Oviraptor philoceratops (№100/42); а - снаружи; о - изнутри
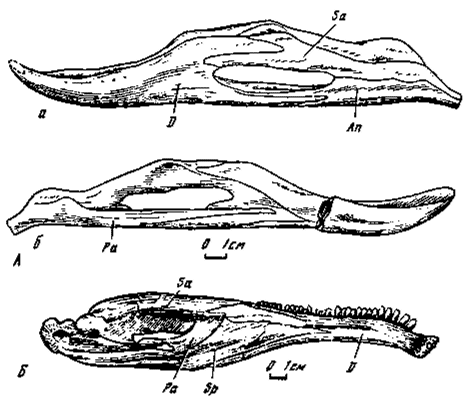
Рис.14. А - Caenagnathus collinsi (Sternberg, 1940); a - левая ветвь снаружи; б - левая ветвь изнутри; Б - Erlikosaurus andrewsi (№100/111, левая ветвь изнутри)
Нормальный тип нижней челюсти
Передний отдел. Передний отдел нижней челюсти занимает немногим большую половину всей ее длины. На значительном протяжении, соответствующем практически длине альвеолярного края, он заметно сужен, наиболее существенно у сегнозаврид. Расширение переднего отдела происходит сразу позади альвеолярного края, где в разной степени, но чаще сравнительно в слабой форме намечается аддукторный выступ к которому прикреплялась ложновисочная мышца - одна из приводивших нижнюю челюсть.
В составе переднего подразделения два главных элемента - зубная и пластинчатая кости. Кроме того, присутствует еще венечная кость, которая служит соединяющим два отдела звеном, топографически входящим в состав обеих частей. Основа переднего отдела - зубная кость - на лингвальной стороне несет продольный меккелев канал, протягивающийся до симфизальной поверхности. Выше отмечалось, что маленький участок переднего окончания нижней челюсти у сегнозавров был беззубым. У орнитомимид и гарудимимид плоскость ростральной части нижнечелюстных ветвей повернута вокруг продольной оси кнаружи так, что образует заметно уплощенную совковидную структуру на переднем конце, чего нет у зубатых групп. Передний заостренный конец пластинчатой кости частично закрывает меккелев канал, несет его вентрально смещенное отверстие и находится в разной степени приближения к симфизу. Угловой отросток пластинчатой кости значительно развит. Венечная кость, как связующий элемент, представлена разными частями в обоих отделах. Ее удлиненный и суженный передний отросток проходит до симфиза вдоль альвеолярного края, закрывая основание (межзубные пластины) зубного ряда. В районе аддукторного выступа на границе двух отделов нижней челюсти участок венечной кости лингвально покрыт пластинчатой и открывается сзади нее в виде треугольного сегмента уже в пределах заднего отдела.
Венечная кость - единственный элемент, пересекающий подвижную границу обоих отделов и более или менее устойчиво связывающий их между собой. В отличие от орнитисхий и ящериц венечная кость у хищных динозавров не формирует восходящего венечного отростка, сильно выдающегося за уровень дорзального края нижнечелюстной ветви. Наряду с задне-дорзальными участками зубной и пластинчатой костей она входит в состав аддукторного выступа. Связующая функция венечной кости лучше всего выражена у монгольских тираннозаврид (тарбозавров). Ранее ее передний отросток обычно принимался за производные зубной кости, и лишь ее самый задний сегмент отождествлялся правильно (Малеев, 1974: Osborn, 1906; Romer, 1956). Вследствие этого связующая роль венечной кости не учитывалась. У дромеозаврид, орнитомимид и гарудимимид передний отросток кости не был столь протяженным. У сегнозаврид венечная кость отсутствовала.
Задний отдел. Преобладающую часть отдела слагает надугловая кость, образующая его дорзальный край до челюстного сустава и латеральную стенку аддукторной ямы. Последняя, как правило, обширная, свидетельствующая о значительной массе прикреплявшихся здесь мышц - аддукторов. Впереди челюстного сустава кость прободена надугловым отверстием, отсутствующим у орнитомимозавров и овирапторозавров. Соединение угловой, надугловой и зубной костей обычно несет оформленную в разной степени (чаще небольшую) наружную нижнечелюстную яму. Передний отросток предсочленовной кости значительно удлинен, его расширенное окончание достигает дорзального края нижней челюсти в области аддукторного выступа, фиксируя на лингвальной поверхности границу между передним и задним отделами. Исключение составляют орнитомимозавры, у которых передний отросток не выходит за пределы вентрального края аддукторной ямы. Челюстной сустав расположен значительно ниже дорзального края заднего отдела. Суставные площадки для мыщелков поперечно удлинены. Ретроартикулярный отросток у дромеозаврид испытывает значительное расширение, при этом у велоцирапторин он заметно уплощен и слегка вогнут. Заднемедиальный край расширения несет выступ или отросток, иногда столбикообразной формы. По аналогии со сходными структурами у некоторых птиц (Воск, 1960) допускается, что этот выступ упирался в квадратную кость при чрезмерном отведении нижней челюсти, препятствуя таким способом нарушению челюстного сустава (Colbert. Russell, 1969). У орнитомимид расширение ретроартикулярного отростка имело значительную высоту и глубокую впадину, задне-медиальный край которого нес выступ, контролировавший отведение нижней челюсти. Однако у тираннозаврид подобный механизм, возможно, отсутствовал, так как ретроартикулярный отросток довольно незначителен.
Соединение двух отделов нижней челюсти отвечает определенной подвижности между ними. Это обеспечивается пазовым соединением составляющих по дорзальному краю и "скользящими" контактами более вентрально расположенных элементов. В дорзальной части нижнечелюстной ветви соединение элементов имеет следующую последовательность: лабиально-зубная-надуповая, далее венечная; лингвально-пластинчатая-предсочленовная. В случае отсутствия венечной кости, как у сегнозаврид, характер соединения остальных элементов сохраняется. В нижнем краю соединение составляющих элементов следующее: зубная + пластинчатая - угловая (лабиально), пластинчатая-предсочленовная (лингвально). Соединение лингвальных элементов (пластинчатой и переднего отростка предсочленовной костей) происходит по дуге, соответствующей заднему краю пластинчатой кости и представлявшей как бы суставное сочленение. Характер соединения лабиальных и лингвальных элементов дорзального и вентрального краев нижнечелюстной ветви обеспечивает легкое вращательное (и поступательное для отдельных составляющих) движение переднего отдела челюсти относительно заднего.
Овирапторидный тип нижней челюсти
Передний отдел. Передний отдел занимает меньшую половину длины нижней челюсти и состоит почти целиком из зубной кости, слагавшей основу рогового подклювья (рис. 13В). Ростральная часть зубных костей сильно утолщена, особенно в симфизе, и представляет собой ковшевидно вогнутое массивное образование. От него назад отходят два отростка: восходящий - расширенный и вогнутый и нижний - сужающийся и прямой, посредством которых кость соединялась с остальными элементами. Эти два отростка ограничивают спереди крупное нижнечелюстное отверстие. Нижний лингвальный край восходящего отростка на всем протяжении с продольным гребнем. Позади ростральной части зубных костей высота переднего отдела резко возрастает. Пластинчатая кость сильно сужена, без углового отростка, участвует в образовании лингвальной поверхности вентрального края нижнечелюстной ветви. Венечная кость отсутствует.
Задний отдел. Надугловая кость формирует основную часть латеральной поверхности отдела. Она ограничивает задний край нижнечелюстного отверстия, отдавая вперед узкий, удлиненный отросток, разделяющий его на две неравные части. Положение отверстия между элементами двух отделов челюсти, вероятно, отражает его первичную связь с наружной нижнечелюстной ямой, получившей в данном случае крайне сильное развитие. Надугловая кость лингвально образует обширную внутреннюю, лабиально же - наружную аддукторные ямы. В дорзальном краю кости имеется крупный сжатый с боков аддукторный выступ для ложновисочной мышцы. Передний отросток предсочленовной кости прямой, без терминального расширения. Угловая кость слагает вентральный край отдела. Челюстной сустав отличается своей расширенностью и рельефностью частей, срединный выступ сагиттально удлинен. Ретроартикулярный отросток удлиненный, суженный, дорзально слегка вогнутый, с небольшим задне-медиальным выступом.
Соединение двух отделов нижнечелюстной ветви в дорзальном краю пазовое. Окончание восходящего отростка зубной кости расщеплено в парасагиттальной плоскости и вмещает передне-дорзальный отросток надугловой кости. По вентральному краю челюсти линейно вытянутые нижний отросток зубной и пластинчатая кости вмещают угловую. Характер соединений допускает вращательное движение переднего отдела относительно заднего, при одновременном более или менее поступательном движении составляющих его вентрального края.
Сравнение двух типов и особенности нижней челюсти ценагнатид. При общерептилийном характере строения оба типа нижней челюсти хищных динозавров существенно различаются. Несмотря на некоторые расхождения в ряде черт, нормальный тип, включающий и беззубую челюсть орнитомимозавров, характеризуется сравнительно удлиненным и узким на большом протяжении передним отделом, слабым развитием аддукторного выступа, присутствием венечной (иногда может вторично отсутствовать) и углового отростка пластинчатой костей, незначительной по величине наружной нижнечелюстной ямой и нередко расширенным ретроартикулярным отростком, обеспечивавшим контроль за максимальным отведением челюсти.
Беззубая нижняя челюсть у овирапторид отличается своей высотой, ее передний отдел укорочен, аддукторный выступ крупный, полностью отсутствуют венечная и угловой отросток пластинчатой костей, наружная нижнечелюстная яма сильно увеличена и смещена вперед, ретроартикулярный отросток узкий, лишенный способности контролировать отведение челюсти.
Различия в строении двух типов в принципе мало сказались на соединении переднего и заднего отделов нижнечелюстной ветви. В обоих случаях характер соединения обеспечивал вращательное движение переднего отдела относительно заднего.
Особое место занимает своеобразная нижняя челюсть у ценагнатид (рис. 14А), которую удобнее рассматривать отдельно. Она отличается удлиненностью и малой высотой, аддукторный выступ умеренно развит. Основа переднего отдела - зубная кость с удлиненной и расширенной ростральной частью, крайне удлиненным симфизом и двумя задними, параллельно направленными верхним и нижним отростками. Окклюзивный край полого вогнут, нижнечелюстная яма узкая, удлиненная, смещена назад. В заднем отделе нижней челюсти у ценагнатид надугловая кость полностью формирует верхний край нижнечелюстной ямы. Передний отросток предсочленовной кости продолжается впереди уровня аддукторного выступа овирапторид.
При пазовом соединении двух отделов нижней челюсти в дорзальной части окончание верхнего отростка зубной кости глубоко расщеплено и вмещает передне-верхний отросток предсочленовной. По нижнему краю ветви удлиненные составляющие также формируют пазовое соединение. Вместе с тем глубина пазов дорзального и вентрального соединений, вероятно, должна была уменьшать подвижность в нижнечелюстной ветви.
Тип строения нижней челюсти у овирапторид и ценагнатид одинаковый. Однако в деталях нижняя челюсть у ценагнатид резко отличается от овирапторидной. Соотношение длины переднего и заднего отделов, высота, степень развития аддукторного выступа, форма ростральной части, ее глубина и линейные размеры, длина симфиза, размеры нижнечелюстной ямы и ее положение - практически все основные признаки преобладающей части нижнечелюстной ветви у овирапторов и ценагната, исключая область челюстного сустава, имеют резко различный характер.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'