
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Глава 5 Филогения анкилозавров
Предков анкилозавров долгое время искали среди стегозавров. Это наиболее близкая к анкилозаврам группа по сравнению с другими динозаврами. Их объединяет наличие остеодермальных элементов в коже, узкий треугольный череп стегозавров сходен с таковым нодозаврид, некоторое сходство обнаруживается и в строении нёба, а также района базисфеноида.
Наиболее вероятным в качестве исходного на основании строения тазового пояса считался род Scelidosaurus (Romer, 1968; Thulborn, 1971). Однако, по мнению У. Кумбса (Coombs, 1978), такие черты в строении посткраниального скелета сцелидозавров, как длинный задний отросток лобковой кости, прободенная ацетабулярная впадина, короткая преацетабулярная часть подвздошной кости и широко вентрально поставленная ее постацетабулярная часть, не могут свидетельствовать в пользу такого предположения. Кроме того, принадлежность к Scelidosaurus материала, на основании которого была высказана возможность сходства короткого переднего отростка лобковой кости с таковым анкилозавров, оспаривается (Rixon, 1968; Charig, 1972). В некоторой степени подтверждением У. Кумбсу может служить находка в Монголии нового нижнемелового анкилозавра Shamosaurus, который имеет типично анкилозавровое строение тазового пояса, с разросшейся горизонтально очень длинной преацетабулярной частью подвздошной кости. И вызывает сомнение, могла ли совершиться сравнительно резкая метаморфоза в этой области таза у анкилозавров при их относительно медленных темпах эволюции и незначительном морфологическом разнообразии (Coombs, 1978) за период с юры (времени существования сцелидозавров) до раннего мела. Другими словами, представляется возможным существование анкилозавров или, по крайней мере, их истинных предков в юре вместе со сцелидозаврами. К этому можно добавить и черты в строении черепа, отличающие анкилозавров от всех стегозавров в целом. К ним относятся: сплошное перекрытие поверхности крыши черепа остеодермальными щитками и срастание их с подстилающими костями, отсутствие верхнего височного окна, которое у стегозавров может быть маленьких размеров, но присутствует неизменно, своеобразный контакт квадратных костей с парокципитальными отростками.
Т. Марианская предполагает, что предки анкилозавров были, по всей вероятности, четвероногоходящими в отличие от орнитопод и цератопсов, исходная бипедальность которых очевидна (Maryanska, 1977). Не исключено, что считающиеся предками всех птицетазовых динозавров триасовые орнитоподы типа Fabrosaurus (Galton, 1974) могли дать начало только первично бипедальным орнитисхиям. Однако в любом случае следует согласиться с мнением У. Кумбса (Coombs, 1978) о достаточно большом морфологическом различии анкилозавров и стегозавров.
Гипотетических предков анкилозавров вероятнее всего предполагать четвероногоходящими, с примитивной мозговой коробкой, значительно менее изогнутой по сравнению не только с типичными динозаврами мезозоя (игуанодонты, гадрозавры, цератопсы), но и, по-видимому, с триасовыми орнитоподами (Fabrosaurus) и даже триасовыми псевдозухиями (Erythrosuchus). В противном случае придется допускать, что в процессе эволюции в ветви анкилозавров наряду с другими крупными морфологическими преобразованиями произошло выпрямление мозговой коробки, что нетипично ни для одной другой группы динозавров.
Вопрос о взаимоотношениях двух семейств анкилозавров У. Кумбс (Coombs, 1978) считал открытым, ссылаясь на то, что выведению их друг из друга мешают такие специфические признаки, как слияние верхних концов квадратных костей с парокципитальными отростками и приближение лопаточного гребня к гленоидной впадине у нодозаврид и рогообразные остеодермальные щитки в постеролатеральных углах крыши черепа; закрытые остеодермами боковые височные окна; сложная система придаточных синусов носовой полости и концевая хвостовая булава у анкилозаврид.
Теперь, после находки черепа Shamosaurus - первого достоверного анкилозаврида из нижнего мела Монголии, появилась возможность построения предполагаемой схемы взаимоотношений данных семейств (рис. 14). Наличие у шамозавра, являющегося по строению нёба уже типичным анкилозавридом, морды с суженной премаксиллярной частью говорит о расширении этой части у более поздних анкилозаврид и об узкомордости как исходном признаке всех анкилозавров. Узкомордость не является препятствием для выведения анкилозавров из стегозавров, как считал А. Ромер (Romer, 1968); против связи обеих групп говорят те различия, о которых шла речь выше. Наличие у ранних нодозаврид (Silvisaurus) премаксиллярных зубов, от которых у более поздних представителей семейства остаются лишь гребни, соединяющие предчелюстную кость с началом каждого зубного ряда, а у анкилозаврид исчезающие вовсе, может свидетельствовать об их большей примитивности. Такие сходные с нодозавридами признаки в морфологии черепа шамозавра, как срастание квадратных костей с парокципитальными отростками; неподвижный контакт птеригоидов с базисфеноидом; круглый, вентрально ориентированный затылочный мыщелок, а также неполное закрытие латерально спускающейся остеодермой квадратного мыщелка могут, по-видимому, говорить о некоторой степени переходности, отраженной в морфологии древних, раннемеловых анкилозаврид, шамозаврин. Общие признаки, имеющиеся и у нодозаврид, и у шамозаврин, являются, очевидно, характерными чертами их предков, но у нодозавров они оказались более стабильными, закрепленными. У анкилозавров морфология черепа в сравнении с исходной изменилась существенно: рано исчезли премаксиллярные зубы, форма черепа модифицирована уже у шамозавра, полное срастание квадратных костей с парокципитальными отростками и птеригоидов с мозговой коробкой сохранилось лишь у двух представителей.
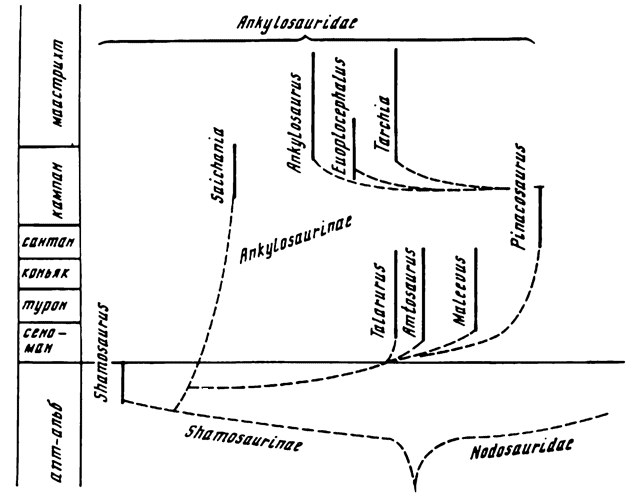
Рис. 14. Схема родовой филогении анкилозаврид
Поскольку у нодозаврид наблюдаются черты, которые, на наш взгляд, с большим основанием можно считать исходными, например премаксиллярные зубы у ранних представителей, открытое боковое височное окно (у анкилозаврид оно закрывается остеодермальным щитком, разрастающимся спереди и сверху с чешуйчатой кости и закрывающим в латеральном аспекте квадратный мыщелок, тогда как у шамозавра эта остеодерма спустилась не так низко и квадратный мыщелок виден сбоку), то, по-видимому, общего предка анкилозавров можно назвать нодозавроподобным. Существенные различия в морфологии анкилозаврид вызваны, возможно, их ранним ответвлением от предкового ствола и приспособлением к несколько иным условиям обитания и образу жизни. Что касается несплошного срастания квадратных костей с парокципитальными отростками, а также птеригоидов и мозговой коробки у большинства анкилозаврид, то это, видимо, можно объяснить развитием ничтожно слабого кинетизма, совершенно отсутствующего у нодозаврид.
Среди представителей семейства нодозаврид У. Кумбс (Coombs, 1978) наметил несколько ветвей: самых примитивных, типа Hylaeosaurus и Struthiosaurus, характеризующихся мелкими размерами, наличием зубов на предчелюстных костях и раздельностью коракоида и лопатки; другая ветвь представлена североамериканскими нодозавридами, для которых характерно срастание лопатки с коракоидом. Ранние представители второй ветви - Sauropelta и Silvisaurus - сохраняют предчелюстные зубы, но у более позднего - Panoplosaurus - они отсутствуют. Ни один из представителей анкилозаврид предчелюстных зубов не имеет. Имеется ряд других морфологических признаков, по которым можно поделить анкилозаврид на две группы. К первой будут отнесены два рода анкилозаврид - шамозавр, как самый древний представитель семейства, и сайхания, обладающая более примитивными чертами строения: относительно узкой премаксиллярной частью морды, срастанием квадратных костей с парокципитальными отростками, срастанием птеригоидов с мозговой коробкой, маленькой межптеригоидной ямой. Вторая группа включает всех остальных монгольских анкилозавров, древние представители которых, возможно, были исходными для североамериканских эвоплоцефалюса и анкилозавра.
Очевидно, первая группа имеет большое морфологическое сходство с нодозавридами, которые по облику, видимо, были максимально приближены к предкам. При переходе к шамозавринам произошли такие изменения в морфологии черепа, как некоторое расширение его в заглазничной области, закрытие разросшимися остеодермальными щитками боковых височных окон, превращение остеодерм в постеролатеральных углах черепа в подобие рогов. Нёбо типичных анкилозаврид, начиная с шамозаврин, отличается ориентированными поперечно птеригоидами и эктоптеригоидами. Затылочный мыщелок у шамозавра, так же как и у нодозаврид, обращен вниз, но он лишен "шейки", на которой сидит у последних, т.е. несколько вдавлен в основание эндокрания. Прямым потомком шамозаврин, по-видимому, можно считать сайханию.
По линии шамозавр - сайхания происходили следующие преобразования: расширение предчелюстной области морды, разрастание остеодермальных щитков с чешуйчатых костей дальше вниз, до закрытия квадратного мыщелка, преобразование остеодерм в постеролатеральных углах черепа в шипы, принятие орбитами более заднего положения и направление их плоскости несколько вперед, разрастание вторичного нёба, укорочение челюстей. Однако без изменений остался характер контакта квадратных костей с парокципитальными отростками и птеригоидов с мозговой коробкой. Вероятно, все остальные анкилозавриды могли быть дериватами этой линии.
При переходе к новой ветви, в основании которой стоит таларурус (наиболее древний из известных позднемеловых анкилозаврид), наряду с преобразованиями в вышеуказанной ветви происходило изменение характера контакта квадратных костей с парокципитальными отростками и птеригоидов с мозговой коробкой, а также увеличение размеров межптеригоидной впадины. Верхние отростки квадратных костей у этих анкилозавров не срастаются с нижней частью парокципитальных отростков, а контактируют с их передней частью, в различной степени прилегая к ней: у таларуруса плотно, у наиболее молодого монгольского анкилозавра - тархии, по-видимому, контакт осуществлялся на связках. На базиптеригоидных отростках у всех анкилозаврид имеются сочленовные площадки, посредством которых осуществлялось базиптеригоидное сочленение.
Тенденция к усилению бронированности, дальнейшей консолидации панциря (Maryanska, 1977; Coombs, 1978) предполагает наличие у предка панциря, состоявшего из отдельных не сраставшихся друг с другом остеодермальных элементов. Вместе с тем у шамозавра мы наблюдаем достаточно высокую степень консолидации элементов наружного скелета в нем имеются те же два кольца (шейное и плечевое), что и у более поздних сайхании и тархии, а также надкрестцовый тонкий остеодермальный щит; кроме того, встречены другие конструкции из срастающихся элементов панциря, не отмечавшиеся у остальных монгольских анкилозавров. Исходя из этого, ясно, что шамозавр не мог стоять непосредственно в основании ветви анкилозаврид. По этим же причинам, а также из-за отсутствия уже на предчелюстных костях зубов и закрытых боковых височных окон мы предполагаем вид предка нодозавро-, а не шамозавроподобным, хотя в обеих группах имеются одинаково древние находки и шамозаврины обладают некоторым набором морфологических черт, характерных, как мы выяснили, для предков.
Расхождение нодозаврид и анкилозаврид произошло предположительно в юре, поскольку раннемеловые представители этих ветвей имеют уже типичное для своего семейства строение.
В результате изучения морфологии и возможных взаимоотношений имеющихся видов была составлена следующая схема (см. рис. 14).
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'