
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Глава 7 Экология
Об образе жизни анкилозавров существует несколько точек зрения. С одной стороны, их рассматривают как животных сугубо наземных, связанных с водоемами лишь потребностью в питье и сборе пищи в прибрежных заводях. С другой стороны, некоторые черты их строения, в первую очередь неуклюжесть, неповоротливость, свидетельствуют о более тесной связи их с водой. Скорее всего, правильно мнение, что жили анкилозавры по берегам рек и озер, богатых растительностью, проводя большую часть времени в воде.
В строении анкилозавров проявляются морфологические признаки, которые в комплексе могут указывать на их амфибиотический образ жизни. Это бочонковидная или сильно уплощенная форма туловища, далеко заходящие на вентральную поверхность ребра, наличие у сайхании отростков ребер, разрастающихся шлейфом и перекрывающих соседние ребра; крупные ноздри, слегка поднятые над уровнем нижнего края черепа (по их внутренним краям, возможно, крепились мышцы, регулирующие закрытие вероятно имевшегося клапана, предохранявшего дыхательный проход от попадания воды); мелкие зубы, способные измельчать только мягкую водную растительность; широкие плоские копытообразные широко расставленные концевые фаланги пальцев для передвижения по мягкому грунту.
Подтверждением такого предположения служат массовые скопления остатков анкилозавров в одном местонахождении. К примеру, на площади менее 30 м² на местонахождении Алаг-Тэг в 1969 г. было найдено девять скелетов анкилозавров различной степени сохранности. Рядом, на площади, не превышающей 14 м², было обнаружено около шести неполных скелетов. Тафономический анализ местонахождения (Твердохлебов, Цыбин, 1974) показывает, что породы, вмещающие большое количество костных остатков анкилозавров, представляли собой обезвоживающуюся часть берега, ставшую болотом вследствие аридизации климата, о чем говорит своеобразная текстура пород.
Подобная массовость остатков анкилозавров характерна и для таких местонахождений, как: Хонгил-Цав, Баин-Ширэ и особенно Бага-Тариач. В этих массовых скоплениях представлены два рода: пинакозавр и в несколько меньшей степени таларурус.
Очевидно, систематичность находок массовых скоплений анкилозавров, не имеющая места ни в одной другой группе динозавров, может быть объяснена, скорее всего, тесной связью жизни животных с водой. Подобная концентрация особей в одном местонахождении дает основательный повод полагать, что анкилозавры вели полуводный образ жизни. К этому же выводу мы приходим, используя морфофункциональный анализ, являющийся, безусловно, более надежным критерием для реконструкции образа жизни животных.
Внешне анкилозавры были достаточно крупными животными: размеры их туловища достигали 4-5, а иногда и 8-9 м. Под поверхностным слоем рога, покрывавшего туловище и голову животного, лежал панцирь из остеодермальных элементов. Остеодермальные щитки и шипы, срастаясь определенным образом, составляли различные конструкции, однако панцирь анкилозавров никогда не достигал степени консолидации черепашьего панциря, а скорее напоминал защитные образования броненосцев. Часто в панцире анкилозавров наблюдается двухъярусность расположения остеодерм. Такое строение, например, имели шейное и плечевое полукольца сайхании и тархии. В двойных конструкциях наблюдается такая закономерность: нижележащий ярус состоит из сравнительно толстых и плотных щитков, верхний же ярус, представленный шипами, значительно более легкий; у поздних анкилозаврид (пинакозавр, сайхания, тархия) стенки шипов тонкие, пористые, с большим количеством отверстий для кровеносных сосудов.
Остеодермальный панцирь анкилозавров, возникнув как защитное образование от неблагоприятных факторов внешней среды (например, высыхания, а также от хищников), возможно, расширил свои функции. Так, например, хотя кость и не является наилучшим проводником теплоты, панцирь вторично мог бы служить животному своего рода аппаратом для регулирования перенагревания и переохлаждения тела.
Учитывая тот факт, что и шипы стегозавров, и шипы анкилозавров увеличивали поверхность туловища, логично предположить, что интерпретация шипов стегозавров как составной части механизма отдачи или поглощения тепла (Farlow, Thompson, Rosner, 1976) в некоторой степени может быть применена и к анкилозаврам. Вряд ли в ходе эволюции шипы анкилозавров становились более тонкостенными и ажурными для усиления роли защиты. Создается впечатление, что количество отверстий, пронизывающих их стенки, явно превышает необходимое для теплоснабжения рогового чехла. Обилие их, если мы примем аналогию со стегозаврами, возможно, объяснялось усиленным кровоснабжением кожи над остеодермами. Сильно васкуляризированная кожа, вместе с остеодермальными элементами могла представлять собой примитивную терморегулирующую систему. В периоды, когда температура окружающей среды превышала температуру тела животного, поверхность его нагревалась. Сосуды, расположенные в верхних слоях кожи, над остеодермами, расширялись, за счет чего кровь начинала стремиться к этим более легко проходимым участкам сосудов, скорость кровообращения увеличивалась, начинала осуществляться усиленная передача тепла от тела к среде, благодаря чему перегревания животного не происходило. Во время резкого падения температуры окружающей среды, когда животное оказывалось под угрозой переохлаждения, сосуды сужались, кровь от поверхностных слоев кожи "отводилась", кровообращение в этом районе замедлялось, теплообмен со средой сокращался. Эффект подобной системы увеличивался благодаря тому, что под шипами, как говорилось выше, обычно располагались остеодермальные пластины, которые, срастаясь, могли работать как временный пассивный ограничитель выделения или поглощения теплоты; другими словами, внутри остеодермального панциря поддерживалась достаточно стабильная температура за счет его пассивной изоляции, а благодаря сильно васкуляризированной коже, покрывавшей поверхность туловища и увеличивающие его площадь шипы, осуществлялась относительно активная терморегуляция.
Различия рисунков поверхностей шипов стегозавров и анкилозавров можно объяснить следующим образом. У стегозавров ветвление кровеносных сосудов происходило на поверхности шипов, в области, непосредственно покрытой при жизни только кожей. У анкилозавров шипы имели роговой покров, поэтому, видимо, для повышения эффективности сосуды ветвились в сравнительно более верхнем слое кожи, отделяющей остеодерму от рога, не оставляя при этом следов ветвления на поверхности шипа.
Поскольку относительного совершенства данная система могла достигнуть лишь у поздних анкилозавров с более пористыми шипами, возможно, первоначально панцирь играл роль лишь пассивного терморегулятора, замедляя перегрев, как, по предположению Л. Хозацкого (1965), это происходит у черепах, одновременно выполняя защитную функцию.
Форма тела анкилозавров варьирует, но преобладает бочонковидная, с различной степенью латерального сжатия. У пинакозавра, единственного из монгольских анкилозавров, туловище уплощено дорсовентрально.
Строение поясов и конечностей говорит об отсутствии каких-либо приспособлений у анкилозавров к быстрому передвижению. Передние конечности, как показала Т. Марианская (Maryanska, 1977) на примере Saichania, были сильно согнуты, плечо ориентировано в сторону почти горизонтально и слегка назад, локтевая и лучевая кости сильно наклонены медиально. Очевидно, на таких конечностях тяжелое бронированное туловище не могло передвигаться со сколько-нибудь значительной скоростью. К этому же выводу приводит и изучение мускулатуры конечностей эвоплоцефалюса (Coombs, 1978в). Сросшиеся коракоид и лопатка, наличие добавочной связи коракоида с позвоночником (Maryanska, 1977) также свидетельствуют об отсутствии приспособлений к подвижности в плечевом поясе. Вследствие сильной изогнутости передних конечностей передняя часть туловища была несколько более приближена к земле и отстояла от нее у сайхании приблизительно на 35 см (Maryanska, 1977), т.е. в целом туловище было поставлено очень низко. Задние конечности были более выпрямлены, поэтому тазовый пояс располагался выше уровня плечевого.
Анкилозавры были четвероногоходящими животными. И если, основываясь на строении ацетабулярной впадины, мы можем констатировать вторичный переход цератопсов к четвероногохождению, то непрободенная acetabulum, по мнению Марианской (Maryanska, 1977), говорит о первичности их четвероногохождения.
Голова соединялась с туловищем посредством сравнительно длинной шеи (7 позвонков). Постеровентрально ориентированный затылочный мыщелок говорит о том, что atlas контактировал с ним сзади-снизу, т.е. под некоторым углом к оси черепа, следовательно, голова сидела не на палкообразно вытянутой шее, как указывалась на некоторых реконструкциях (Lull, 1922). Об изогнутости шеи и наличии некоторого прогиба свидетельствует и развитый в разной степени (у таларуруса слабо, у пинакозавра и тархии значительнее) скос фасеток шейных позвонков. С помощью этой конструкции шея избегала нагрузок на излом, которые неизбежно возникали при наличии тяжелой бронированной головы, и прогиб служил в качестве амортизатора. Благодаря такой форме затылочного мыщелка (суставная поверхность больше повернута вентрально) движения головы в горизонтальной плоскости были более свободны, чем в вертикальной, что, возможно, было важно при почти латеральной постановке глаз на черепе.
Движения нижней челюсти также были сильно ограничены за счет конструкции квадратных и птеригоидных мыщелков. Таким образом, челюсти были в состоянии осуществлять лишь режущие движения в вертикальной плоскости с помощью ряда мелко зазубренных по краю зубов, на которых у большинства анкилозавров не обнаружено сколько-нибудь достоверных следов стирания. Животное срывало растения посредством острых, загнутых в виде тупого клюва премаксилл, скорее всего, покрытых при жизни роговым чехлом, с помощью примитивной зубной системы мелко настригало мягкую сочную водную растительность, не наносящую повреждений эмали зубов.
Анализируя строение гиоидного аппарата сайхании, Т. Марианская (Maryanska, 1977) пришла к мнению, что он более всего сходен с таковым варанов, т.е. возможно, анкилозавры были не исключительно растительноядными животными, а могли питаться, к примеру, насекомыми или их личинками (Nopcsa, 1928). В этом случае длинный язык, который мог иметься у них (по аналогии с варанами), помогал в поисках и добыче пищи.
Вероятное присутствие якобсонова органа (Maryanska, 1971, 1977) также говорит в пользу того, что рацион анкилозавров, возможно, не был ограничен только растениями, хотя последние, несомненно, составляли его значительную часть.
Глаза анкилозавров в процессе эволюции передвигались несколько назад, изменяя свою плоскость от параллельной длинной оси черепа до немного повернутой вперед и составляющей с плоскостью длинной оси острый угол. За счет подобного поворота плоскости глазниц увеличивалась площадь обзора и появлялась основа для зрения с перекрытием полей обзора (рис. 15).
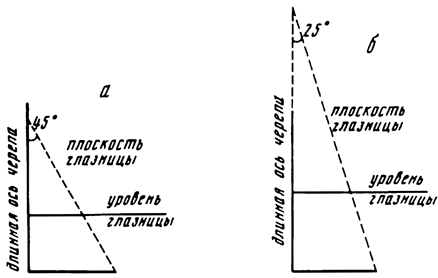
Рис. 15. Схема направленности плоскости орбит в черепе анкилозаврид а - шамозавр; б - тархия
Т. Марианская (Maryanska, 1977), анализируя выдвинутую впервые Е.А. Малеевым (1954) на примере сирмозавра (= пинакозавра) гипотезу о зарывании анкилозавров, приходит к выводу, что в пользу этого предположения могут говорить и уплощенная форма туловища пинакозавра, и наличие панциря на вентральной стороне, и короткие килеватые пластины, расположенные латерально, а также возможное наличие костного века (Coombs, 1972), которое могло быть приспособлением для защиты глаз при зарывании. Однако костей, названных у эвоплоцефалюса костным веком, ни у пинакозавра, ни у других монгольских анкилозавров не было обнаружено, хотя они есть и у многих других динозавров. Кроме того, у большинства анкилозавров туловище сжато с боков. От принятия предположения о зарывании анкилозавров нас удерживает и тот факт, что для этого анкилозавры, по-видимому, были слишком крупными животными.
Хвост анкилозаврид имел несколько необычное строение. Подвижной оказывалась лишь его проксимальная часть, тогда как дистальная была образована сильно измененными позвонками, соединенными не только широко перекрывающимися пре- и постзигапофизами и сочлененными соседними гемальными дугами, но и удлиненными окостеневающими сухожилиями (табл. II). Она представляла собой, очевидно, ударную палицу, на конце которой располагался остеодермальный диск в виде булавы. Существует предположение (Рождественский, 1974), что длинный сильный хвост мог служить анкилозаврам активным органом защиты. И хотя у анкилозаврид уже имеется пассивная защита в виде панциря, трудно пока найти другое назначение хвоста столь своеобразной конструкции.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'