
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Палеонтологический метод и основы стратиграфической классификации
Основным методом определения относительного возраста пород является палеонтологический метод. Его применение базируется на дивергентном поступательном развитии органического мира и принципе необратимости эволюции, установленных Дарвином (1859).
Стратиграфическое расчленение осадочных пород, развитых в данном регионе, начинается со всестороннего изучения естественных обнажений и кернов скважин, вскрывающих эти отложения в закрытых районах. Изучается литологический состав отдельных слоев, их взаимоотношения друг с другом, причем принимается, что при ненарушенном залегании подстилающий слой является более древним, а покрывающий более молодым (принцип Стенона). Если между слоями нет следов перерыва, то их формирование шло последовательно и непрерывно; если же между ними наблюдается так называемое стратиграфическое несогласие, то предполагается наличие перерыва в осадконакоплении, а также возможность размыва нижележащих слоев. В таких случаях важно установить размеры перерыва. О следах перерыва свидетельствует неровная, эродированная поверхность нижнего слоя, наличие в подошве вышележащего слоя грубообломочного материала - гравия, гальки, желваков фосфоритов, остатков переотложенных скелетов различных организмов. В кровле нижнего слоя наблюдаются следы зарывающихся и сверлящих организмов: норы ракообразных, сверлящих и зарывающихся двустворок, заполненные осадком вышележащего слоя. Если между слоями наблюдается угловое несогласие, то это объясняется тем, что подстилающие породы до образования вышележащих подвергались воздействию тектонических движений, в результате которых их первоначальное горизонтальное залегание было изменено.
Из каждого слоя или группы слоев изучается систематический состав ориктоценозов. Для этого извлекаются органические остатки - фоссилии, которые после технической обработки определяются. При извлечении из слоев ископаемых отмечаются особенности их захоронения, распределение в слое, количественные взаимоотношения между отдельными видами, проводятся палеоэкологические наблюдения, выясняется их прижизненное положение, принадлежность к различным экологическим группам. Для выяснения наличия спор, пыльцы и микрофоссилий отбираются образцы для дальнейшей технической обработки. Предварительный анализ фоссилий в полевых условиях позволяет отличать отложения, образованные в морских или континентальных условиях, в лагунах или пресноводных бассейнах. Однако для "немых" толщ, не заключающих органических остатков, решить этот вопрос иногда бывает очень трудно.
На основании изучения естественных обнажений и прослеживания по простиранию слоев или пачек слоев составляется сводный стратиграфический разрез изучаемого региона. Для обоснования возраста слоев, выделенных в разрезе, - определяются органические остатки: выясняются их систематическая принадлежность, их ареалы и время существования. Если сохранность материала не позволяет определить органические остатки до вида, то тогда, в зависимости от сохранности материала и опытности палеонтолога, определение дается до рода, до семейства и т. д. (в открытой номенклатуре). В тех случаях, когда встречены совершенно новые формы, они описываются как новые виды, роды. Крайне важным для установления возраста пород, из которых извлечены органические остатки, является их анализ с точки зрения биостратиграфического значения каждой из изученных форм. Отдельно анализируются остатки растений (отпечатки листьев, остатки плодов, семян, споры и пыльца), позвоночных и беспозвоночных. Среди морских форм выделяют организмы бентоса, нектона и планктона, выявляют формы, широко распространенные (космополиты), и формы, развитые только в данном регионе (эндемичные); формы, жившие на месте находок (автохтонные - кораллы, губки и другие прикрепленные формы) или попавшие в ориктоценоз из других биоценозов (аллохтонные).
На основании определения состава организмов, живших в изучаемый отрезок времени, и выяснения их стратиграфического значения выделяются отдельные стратиграфические единицы, или стратоны, и обосновывается их возраст. Дробность стратиграфических подразделений зависит от масштаба геологосъемочных работ и масштаба карт, которые необходимо составить, от задач, стоящих перед тематическими исследованиями, а также от обилия и сохранности палеонтологического материала.
Прежде чем остановиться на значении некоторых групп беспозвоночных для целей стратиграфии, следует напомнить, что палеонтологический метод впервые применил У. Смит при составлении шкалы стратиграфических образований Англии (1799) и при составлении им первой геологической карты Англии, Уэльса и юга Шотландии (1815). Эту возможность использования ископаемых органических остатков - для выделения отдельных формаций вслед за Смитом применили Броньяр и Кювье при изучении разрезов Парижского бассейна.
Важное значение для разработки основ стратиграфической и геохронологической шкал имело предложение первого профессора Оксфордского университета Англии Бакленда (1818) о выделении соподчиненных стратиграфических единиц. Бакленд, подобно Линнею, разделившему весь животный и растительный мир на соподчиненные таксоны, предложил следующую иерархию стратиграфических единиц: класс, порядок, формация, слой. Класс рассматривался как высшая стратиграфическая единица, слой - как низшая. В частности, три слоя: серый мел, нижний мел и верхний мел объединялись в формацию "мел", которая относилась к порядку "горизонтальному", т. е. пород, залегающих горизонтально, и к классу "вторичных пород", содержащих органические остатки и фрагменты более древних пород. Для каждого слоя кроме названия указывались литологический состав, мощность и наиболее типичные места распространения. В дальнейшем эти названия стратиграфических единиц изменились, но принцип выделения остался. В это же время (1820) в Англии было предложено объединять формации в системы. За сравнительно короткий срок (1818-1841 гг.) в Западной Европе были выделены все системы, принятые в настоящее время.
В 1841 г. Д. Филлипс предложил объединить системы в такие единицы, как палеозой, мезозой и кайнозой. Возникла новая отрасль геологии - стратиграфия, развитие которой сыграло важную роль для становления палеонтологии. Перед стратиграфией были поставлены две взаимно связанные задачи: 1) расчленение разреза и выделение соподчиненных стратиграфических единиц, 2) внутрирегиональная и межрегиональная корреляция выделенных стратонов. В соответствии с господствующим в первой половине XIX в. мнением о постоянстве и неизменяемости видов предполагалось, что для каждой формации и системы характерны свои фауна и флора, которые в конце каждого периода уничтожались, а в начале каждого последующего создавались заново.
Известный французский палеонтолог и стратиграф д'Орбиньи (1802-1857) в 1842 г. в своей крупной работе "Палеонтология Франции" предложил разделять систему на ярусы - стратиграфические подразделения, объединяющие группы слоев, часто разного литологического состава, но заключающие характерную для каждого яруса фауну. Каждый ярус д'Орбиньи назвал, в соответствии с идеей Смита, по местонахождению типичного разреза: например, оксфордский ярус юрской системы был назван по месту его развития вблизи г. Оксфорда, Англия; барремский и аптский ярусы меловой системы названы по местам их развития в одноименных деревнях во Франции - Баррем и Апт. Вначале ярусное деление было предложено для меловой и юрской систем. В дальнейшем ярусы были выделены во всех системах, однако для некоторых систем до сих пор нет общепринятого ярусного деления (например, для кембрия, палеогена, неогена).
Значительные трудности возникают при применении схемы ярусного деления, разработанной для морских отложений при изучении континентальных пород, в которых встречены наземные организмы. Так, для морских девонских отложений выделение ярусов основано на этапах развития аммоноидей и брахиопод, а для водных-континентальных бассейнов - на этапах развития бесчелюстных - остракодерм и панцирных рыб (плакодерм). Ярусная схема для морских отложений резко отличается от ярусной схемы континентальных пород и их сопоставление представляет обычно большие трудности.
В 1856-1858 гг. немецкий палеонтолог и стратиграф Оппель (1831-1865) предложил ярусы разделять на более мелкие стратиграфические единицы - зоны, отмечая, что каждая зона содержит характерные виды, отличные от видов, встречаемых в выше- и нижележащих слоях. Оппель предложил первое зональное деление для юрских отложений Западной Европы и рекомендовал каждую зону называть по характерному виду, или "виду-индексу".
Палеонтологический метод казался в то время простым и практически удобным, так как расчленение и увязка разрезов проводилась на сравнительно небольшой территории Западной Европы. Для геологов первой половины XIX в. ископаемые организмы были только свидетелями относительного возраста определяемых слоев и еще не стали звеньями непрерывной цепи жизни,, которыми их сделала эволюционная теория Дарвина. Благодаря усилиям Дарвина и его последователей палеонтология превратилась в эволюционную науку; органические остатки стали звеньями цепи жизни на Земле. Однако Дарвин отметил, что геологическая летопись несовершенна и отличается неполнотой; значительная часть памятников истории жизни на Земле была разрушена и исчезла бесследно. Тем не менее имеющиеся палеонтологические документы представляют большую ценность для расчленения и корреляции разрезов. Фауна из отложений любого стратона по уровню организации всегда будет промежуточной между фаунами предшествующего и последующего стратонов. Это достаточно четко выявляется по различным группам беспозвоночных, но особенно выразительно при сравнении фаун позвоночных. Особое внимание Дарвин обратил на кажущееся внезапным появление в кембрии разнообразных представителей животного мира, объясняя это не внезапностью, а метаморфизмом докембрийских пород либо недоступностью их для наблюдений. Не менее важной проблемой стала после открытия и освоения новых материков и земель проблема геологической синхронизации отложений, развитых на удаленных территориях, но относимых к одним и тем же стратиграфическим подразделениям.
Во второй половине XIX в. был накоплен большой фактический материал по стратиграфии и фауне, и на первой сессии Международного геологического конгресса (МГК), состоявшейся в Париже в 1878 г., была создана комиссия по унификации стратиграфической номенклатуры. Результаты работы комиссии были рассмотрены второй сессией МГК в Болонье (1881) и была принята схема стратиграфических подразделений и их хронологических эквивалентов. На восьмой сессии МГК в Париже в 1900 г. в основу стратиграфической классификации были положены подразделения геологического времени, а подразделения осадочных отложений рассматривались как эквиваленты этих временных подразделений. Принятая сессией схема двойной классификации получила в дальнейшем всеобщее признание:

В этой схеме не было эквивалента эре. В нашей стране им стала группа, а для эпохи - отдел. Для зоны не было принято геохронологического эквивалента, была рекомендация употреблять момент, фаза, но эти термины не получили распространения. Из классификации видно, что для указания времени существования той или иной группы организмов должны быть использованы временные категории (например, трилобиты появились в раннем кембрии и вымерли в середине пермского периода), но при указании о находках в соответствующих слоях используются стратиграфические единицы ("остатки трилобитов встречены в отложениях нижнего кембрия"). Таким образом, эти две шкалы взаимно связаны: одна указывает на время, другая - на отложения, накопившиеся за соответствующий отрезок времени.
В областях и районах, где не могут быть выделены единицы международной стратиграфической шкалы, устанавливают местные стратиграфические подразделения: серию, свиту, пачку. Свита объединяет комплекс отложений либо однородных в литологическом отношении, либо представляющих более или менее однообразное чередование определенных типов пород, накопившихся в течение определенного этапа развития данного региона. При характеристике свиты на первое место выдвигаются литологические признаки, а затем, если имеются органические остатки, дается палеонтологическая характеристика. Свиты объединяются в серии и в свою очередь разделяются на пачки. Серия рассматривается как крупное литолого-стратиграфическое подразделение, объединяющее несколько свит. Все местные литостратиграфические единицы выделяются как определенные комплексы осадочных, магматических и метаморфических пород, ясно отграниченные от смежных комплексов, литологически легко опознаваемые в поле, имеющие достаточно широкое площадное распространение.
Осложнения, возникающие при применении названий вспомогательных единиц, связаны с тем, что в странах, говорящих на английском языке, для литостратиграфических единиц приняты: группа, формация, член (member) и слой. Термин "серия" используется в качестве эквивалента русского понятия "отдел", а "группа" в качестве эквивалента "серии".
В последние годы в связи с накоплением нового фактического материала вновь значительно повысился интерес к вопросам стратиграфической классификации и номенклатуры. Американские стратиграфы предложили выделять три основные категории стратиграфических подразделений: литостратиграфические, биостратиграфические и хроностратиграфические.
Литостратиграфические единицы выделяются по литологическим признакам и закономерностям накопления осадков.
К биостратиграфическим единицам в настоящее время относится только зона, остальные подразделения рассматриваются как просто стратиграфические. Фактически к биостратиграфическим единицам должны быть отнесены все стратиграфические единицы.
К хроностратиграфическим подразделениям относятся такие, которые объединяют отложения, образованные в течение определенного отрезка времени, и границы между ними представляют изохронные (одновозрастные) поверхности, не зависящие от литологического состава отложений и их палеонтологической характеристики, т. е. представляют лишь отвлеченное понятие. Поэтому выделение хроностратиграфических подразделений как таковых излишне.
В соответствии с рекомендациями геологического конгресса желательно выделять следующие подразделения:

При написании названия яруса в разных странах приняты следующие окончания:

В настоящее время выделены пять эр: архейская, протерозойская, палеозойская, мезозойская и кайнозойская. Первые две образуют эон криптозой, последующие три - эон фанерозой.
Архей и протерозой выделяются по данным абсолютного возраста и по крупным тектоно-магматическим циклам. Для расчленения протерозоя используются строматолиты и онколиты - известковые продукты жизнедеятельности сине-зеленых водорослей. Продолжительность криптозоя составляет около 4 млрд. лет, в то время как длительность фанерозоя не превышает 570 млн. лет. Радиологические методы (калий-аргоновый, рубидий-стронциевый, уран-торий-свинцовый) дают цифры абсолютного возраста пород. Однако к этим цифрам следует подходить с большой осторожностью, так как допускаемые ошибки бывают большими (табл. 1).

Таблица 1. Деление докембрия в СССР
(М. А. Семихатов (1972) предлагает рассматривать в качестве эр рифей и афебий, последний объединяет ранний и средний протерозой, а протерозой поднять до ранга зона, противопоставляя его фанерозою и архею.)
Существенная перестройка структуры земной коры, которая произошла на рубеже 2600±100 млн. лет тому назад, служит границей между археем и протерозоем. Отложения с возрастом от 2600±100 до 1700±50 млн. лет отвечают второму крупному этапу развития земной коры, который относится к раннему и среднему протерозою; наконец, последний этап в докембрии выделяется как поздний протерозой, или рифей, от 1700±50 до 570-600 млн. лет. Терминальная часть рифея известна под названием венда, в отложениях которого были обнаружены первые бесскелетные отпечатки многоклеточных организмов. Наиболее богатым местонахождением вендского комплекса фауны является в настоящее время местонахождение Эдиакара в Южной Австралии, открытое в 40-х годах нашего столетия. В Эдиакаре обнаружены остатки губок, кишечнополостных, червей, членистоногих, иглокожих.
Фанерозойский эон разделен на три эры: палеозойскую, мезозойскую и кайнозойскую (см. табл. 2). В настоящее время предлагается палеозойскую эру разделить на две самостоятельные: собственно палеозойскую и метазойскую (Друщиц, Шиманский, 1962). Эры были выделены на основании изменения таких крупных таксонов, как отряды, подклассы и даже классы; периоды - на основании изменений состава менее крупных таксонов - отрядов, надсемейств. Для расчленения отложений каждого периода используются наиболее быстро эволюционирующие группы организмов. Для разделения кембрия важную роль играют трилобиты, для ордовика и силура, - граптолиты, для девона и для мезозоя - аммониты, для палеогена и неогена - фораминиферы, двустворки и брюхоногие моллюски.
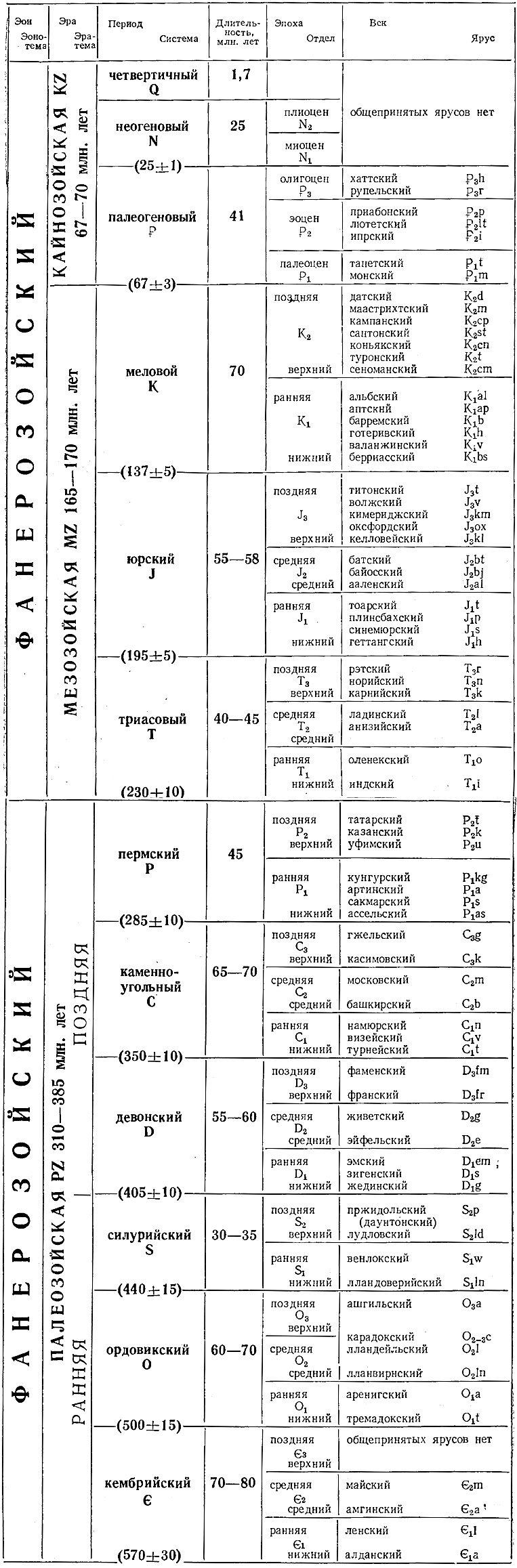
Таблица 2. Основные геохронологические и стратиграфические подразделения
На основании развития этих же групп животных выделяются зоны - наиболее дробные стратиграфические единицы. Объем и границы зоны устанавливаются по распространению одного или нескольких характерных видов, определяющих название зоны. В отложениях любой зоны могут встречаться остатки разных категорий организмов (рис. 23): 1) жившие только в течение хронозоны, 2) появившиеся в предыдущей хронозоне и испытавшие расцвет в изучаемый отрезок времени, 3) появившиеся в данной хронозоне, достигшие расцвета, но продолжающие свое существование и дальше, 4) появившиеся в данной хронозоне, но испытавшие расцвет позднее, 5) появившиеся ранее, характерные для данного времени, но вымирающие позднее, 6) появляющиеся на рубеже двух зон, 7) доживающие свой век в данной хронозоне, 8) транзитные, существующие в течение относительно длительного времени, 9) вымершие в предыдущее время, 10) появляющиеся в следующей зоне. Для выделения зон наиболее важное значение имеют виды, относимые к 1-3-й, 5-й группам (виды и роды).
Но кроме короткого времени существования для широких межрегиональных корреляций важно, чтобы эти организмы обладали и крупными ареалами; при малых ареалах выделяются только местные зоны, или лоны (т. е. локальные зоны, название, предложенное недавно Г. Я. Крымгольцем). К числу организмов, имеющих большие ареалы, относятся планктонные и нектонные организмы (например, аммониты, белемниты, граптолиты, планктонные фораминиферы). Ареалы многих видов обычно ограничены палеобиогеографическими областями или провинциями. Поэтому зоны, выделенные в одной палеозоогеографической области, обычно не могут быть распространены на другие области, и при корреляции одновозрастных зон, выделенных в разных биогеографических областях, возникают значительные трудности. В некоторых случаях помогают виды-космополиты. Еще большие трудности возникают при корреляции морских и континентальных отложений. Здесь на помощь приходит изучение спор и пыльцы и наземных растений, часто заносимых в водные бассейны, изучение разрезов в промежуточных зонах, в которых наблюдается чередование морских и континентальных отложений.
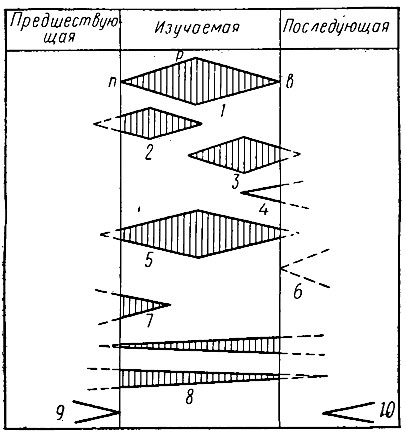
Рис. 23. Возможные варианты распространения видов (родов) в соответствующих зонах (Жемчужников, 1934, с изменениями): п - появление, р - расцвет, в - вымирание (объяснение см. в тексте)
Для одного и того же региона часто предлагают зональное деление по разным группам. Естественно, что зоны, выделенные по одним группам, не будут совпадать с зонами, выделенными по другим, поскольку скорости эволюции и закономерности развития у разных групп различны и обычно не совпадают. Все это служит основанием для дискуссии практически по любой границе между стратонами. Чтобы достичь однозначного решения о границах, правильнее будет проводить зональное деление по одной ведущей группе и рассматривать это деление как эталонное, а развитие других групп животных контролировать и сравнивать с этапами развития эталонной группы. Для каждой зоны, яруса избирается стратотипический разрез, или стратотип, который должен быть наиболее полным, без перерывов, с верхней и нижней границами и, самое главное, с остатками ведущей группы животных.
Наряду с зонами как стратиграфическими подразделениями выделяются так называемые биозоны. Биозоны не представляют собой единиц ни стратиграфических, ни геохронологических подразделений. Биозоной называются отложения, отвечающие полному распространению какой-либо одной определенной систематической группы организмов (вида, рода, семейства, класса и т. д.) (Степанов, 1958). Интервал времени, соответствующий биозоне и отвечающий полному распространению любой группы, называют биохроном. Время максимального развития группы носит название гемера (гр. hemeros - день, время); отложения, накопленные в течение гемеры, - эпиболь. Например, биозона аммонитов охватывает девон - мел, а их гемера относится к юре и мелу. Предложено также выделять тейльхрону - время распространения вида в данном районе - и тейльзону - слои, накопленные за это же время. Тейльзона и тейльхрона в разных районах, как правило, не совпадают, так как существование каждого вида в каждом регионе обусловлено определенным комплексом условий и их изменение может приводить либо к полному исчезновению вида, либо к его миграции в соседние районы. Виды бентоса особенно чувствительны к изменениям фаций, в то время как нектонные и планктонные формы от них не зависят. Представление о биозоне любого таксона складывается на основании анализа данных о тейльзонах, а представления о биохроне - на основании местных тейльхрон. Для палеонтолога крайне важно знать как биохрону, так и тейльхрону каждого таксона на всем ареале и на отдельных его участках.
Наиболее короткий период - четвертичный; его длительность оценивается по последним данным в 1,7 млн. лет, т. е. не превышает продолжительности одной зоны остальных периодов; поэтому он не разделяется на отделы и ярусы.
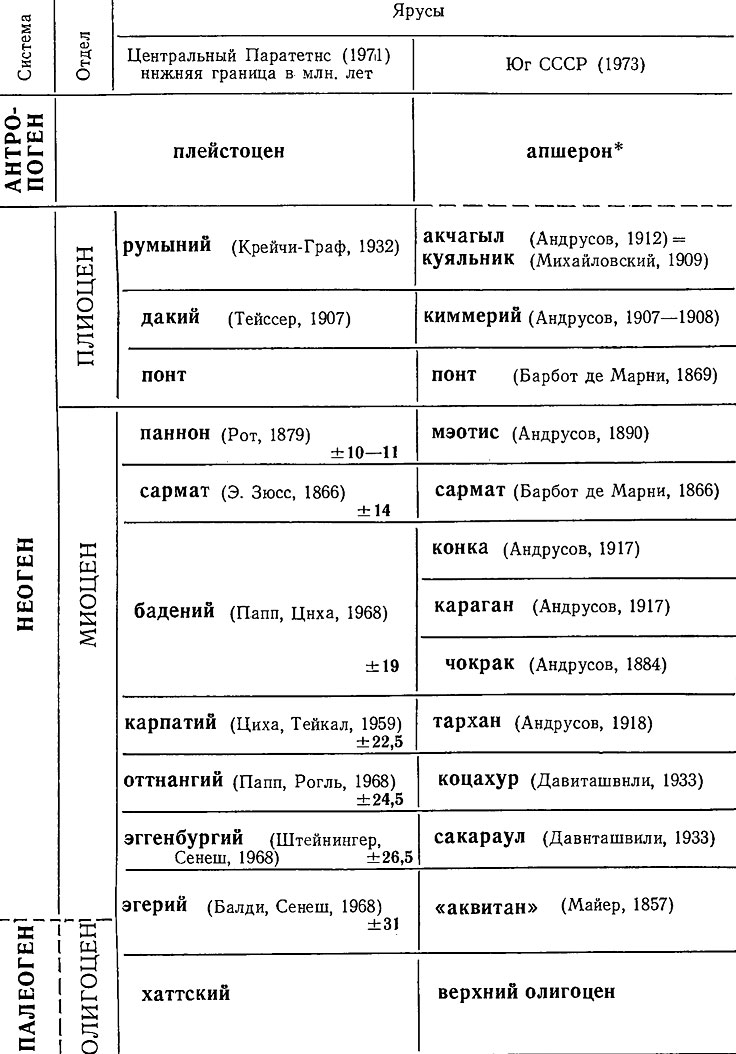
Таблица 3. Схема ярусного деления неогена
(В СССР апшерон пока отнесен к плиоцену.)
Применение палеонтологического метода в стратиграфии часта осложняется по различным причинам: переотложение органических остатков, эндемизм фаун, реликтовые и реккурентные формы.
Переотложение органических остатков и нахождение их во вторичном залегании очень широко распространены в природе. Как правило, переотложение возникает в результате размыва подстилающих пород, и переотложенные органические остатки чаще всего приурочены к базальному горизонту. В нем может встречаться галька, обломки и целые глыбы палеонтологически охарактеризованных пород, из более древних отложений. Так, в базальном горизонте альба (нижний мел) центральной части Крыма встречены глыбы известняков с фауной баррема и отдельные переотложенные раковины аммонитов, нижнего и верхнего апта. Таких примеров можно привести довольно много.
Эндемичные формы обладают ограниченными ареалами и могут быть использованы только для ограниченных районов; к ним также относятся только что установленные новые виды, известные вначале локально. Примером эндемичных фаун служит фауна Байкала, состоящая на 75% из эндемичных форм, фауна ряда островов (например, Галапагосских островов, блестяще исследованных Ч. Дарвином), долин рек, горных областей. Эндемизм чаще всего связан с изоляцией. Следует помнить, что эндемизм может быть обусловлен не только изоляцией и древностью форм, но и молодостью форм, только что возникших недавно в пределах области и не успевших распространиться.
К реликтовым, или суперститовым, формам относятся древние формы, живущие среди более молодых; например, современные: неопилина галатея - последний представитель класса моноплакофор, кистеперая рыба - латимерия, последний представитель мезозойских целекантид. Находки подобных форм в ископаемом состояния могут привести к ошибкам при определении возраста.
К реккурентным формам относятся одни и те же формы, которые появляются в различных стратиграфических горизонтах в связи с повторением в разрезе благоприятных для них фаций. Реккуренция связана с миграцией фауны при возникновении неблагоприятных условий для ее обитания и возвращением ее при восстановлении условий близких к прежним. Реккурентные фауны чаще всего наблюдаются в условиях ритмичного чередования слоев, например в Донбассе морские фауны повторяются в прослоях известняков, чередующихся с континентальными угленосными отложениями. Фаунистические комплексы доманиковой фации (битуминозные известняки) повторяются в живетском, франском и фаменском ярусах девона на востоке Русской платформы и в Приуралье, что вначале затрудняло в каждом отдельном случае определение их возраста.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'