
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Подкласс Ammonoidea. Аммониты
Общая характеристика. Раковина преимущественно планоспиральная, но может быть иного типа: прямой, согнутой, крючкообразной, спирально-конической. Жилая камера от 1/2 до 1,5-2 оборотов; протоконх шаровидной или валиковидной формы, диаметром от 0,3 до 0,8 мм. Сифон начинается шаровидным вздутием - цекумом и прикрепляется изнутри к протоконху просифоном; на первых оборотах положение сифона может варьировать от центрального до краевого; начиная со второго - третьего оборота занимает вентральнокраевое положение; исключение составляет позднедевонский отряд климений с дорзальным положением сифона. Сифон тонкий; септальные трубки от коротких до длинных, у палеозойских форм направлены назад (как у наутилоидей) - ретрохоанитовые, у мезозойских направлены вперед - прохоанитовые.
Периферические края септ изогнуты, волнисты и соответственно перегородочная, или лопастная, линия состоит из значительного числа лопастей и седел; по особенностям развития различают четыре основных типа лопастных линий: агониатитовый, гониатитовый, цератитовый и аммонитовый. Устье закрывалось известковыми крышечками-аптихами. Челюсти и радула по своему типу строения напоминают таковые колеоидей; имелся чернильный мешок. Раковина гладкая или скульптированная, до 2 м в диаметре.
Развитие, по-видимому, прямое, без личинок. Из яйца выходила аммонителла, имевшая протоконх и первый оборот, состоявший из жилой и одной или двух гидростатических камер. Яйца были маленьких размеров и довольно многочисленные. Раковина аммонителлы и первые две септы построены из призматического слоя; все последующие обороты состоят из трех слоев: двух призматических и лежащего между ними пластинчатого; все септы, кроме двух первых, построены только из пластинчатого слоя. Аммониты произошли от бактритоидей в начале девона и вымерли в конце мелового периода.
Строение мягкого тела. О строении мягкого тела аммонитов за последние годы получены новые интересные данные. О форме и размерах мягкого тела можно судить по форме и размерам жилой камеры, а также по изменению формы поперечного сечения в онтогенезе. На ранних стадиях тело было сжато в дорзовентральном направлении, затем после третьего - четвертого оборота принимало округленную форму и далее в зависимости от типа раковины - сжатое с боков, узкое и высокое или низкое и широкое, с малообъемлющими краями или, наоборот, с сильнообъемлющими. В зависимости от длины жилой камеры тело было толстым и коротким или, наоборот, длинным и червеобразным и занимало до 1,5-2 оборотов.
Тело было окружено мантией, которая прикреплялась к последней перегородке и к устьевому краю; по-видимому, двумя мускулами-ретракторами оно прикреплялось к бокам раковины; их отпечатки обнаружены в последнее время на ядрах. По неясным отпечаткам, обнаруженным с помощью рентгеновских лучей, установлены пиритизированные остатки мягкого тела - головной комплекс с небольшим числом (8-10) коротких рук, воронка и остатки пищеварительного тракта. Сравнительно недавно обнаружены остатки радулы, челюстного аппарата и чернильного мешка. Радула состоит из 7 рядов хитиновых зубчиков, как у современных колеоидей. Чернильный мешок, обнаруженный у юрских форм, в задней части жилой камеры имеет вид мешка грушевидной формы, обращенный суженной частью к устью. Его размеры составляют 1/4 длины жилой камеры, что соответствует относительным размерам чернильного мешка современных кальмаров. По-видимому, у аммонитов имелся капюшон, выполнявший ту же функцию, которую он выполняет у наутилуса, т. е. закрывал устье при втягивании тела внутрь раковины.
У многих аммонитов капюшон выделял известковую крышку, состоящую из двух пластинок - аптихов. Аптихи кальцитовые, плотно закрывали устье раковины. В ископаемом состоянии они встречаются обычно отдельно от раковины, так как после гибели тело аммонита вываливалось из жилой камеры и пустые раковины, вероятно, разносились на большие расстояния. Аммониты подобно всем головоногим были раздельнополы. Удается установить по раковинам половой диморфизм - раковины самок отличаются от раковин самцов формой и размерами, а также числом оборотов. В жилой камере самки были обнаружены оболочки яиц. В нижнемеловых отложениях (апт) около Ульяновска была найдена кладка яиц с эмбрионами аммонитов, у которых была раковина в 3/4 или полный оборот, имевшая 1-2 перегородки либо без перегородок.
Строение раковины. Форма раковины крайне разнообразна (рис. 200). Раковина состоит из протоконха, фрагмокона и жилой камеры. Любая раковина аммоноидей начинается с протоконха, или начальной камеры, шаровидной или бочонковидной формы диаметром от 0,3 до 0,8 мм. Вокруг протоконха происходит навивание всех остальных оборотов у планоспиральных раковин. Форма фрагмокона определяет основные черты строения раковины. Среди планоспиральных различают эволютные и инволютные формы, У первых умбилик широкий, у вторых узкий.

Рис. 200. Подкласс Ammonoidea. Форма раковин: 1-4 - планоспиральные, 5-15 - гетероморфные: 1 - кадикон (Cadoceras); 2 - сферокон (Phyllopachyceras); 3 - оксикон (Timanites); 4 - платикон (Haploceras); 5 - криокон (Crioceras); 6-7 - oтурикон (6 - Hyphantoceras, 7 - Ostlingoceras); 8, 14 - гетероконы (8 - Anaclinoceras, 14 - Heteroceras); 9-10 - анцилокон (9 - Ancyloceras, 10 - Worthoceras); 11 - бакуликон (Baculites); 12, 13 - птихоцераконы (12 - Ptychoceras, 13 - Hamulina); 15 - скафитикон (Scaphites)
В зависимости от формы поперечного сечения эти раковины могут иметь различное очертание: кадиконы - эволютные, сжатые в дорзовентральном направлении, сфероконы - инволютные, шаровидные, оксиконы - инволютные, с заостренным вентральным краем, платиконы - с прямоугольным поперечным сечением и плоскими боками, серпентиконы - эволютные, с медленно возрастающими оборотами. Все иные типы раковин выделяются в группу гетероморф: криоконы - планоспиральные раковины, состоящие из несоприкасающихся оборотов, анцилоконы - обладающие спирально-плоскостной частью и длинной выпрямленной, заканчивающейся крючкообразным изгибом, гетероконы - имеющие спирально-коническую и длинную выпрямленную часть, как у анцилодератид, турриконы - спирально-конические, иногда с разобщенным последним оборотом, бакуликоны - напоминают литуитиконы, состоящие из одного-двух спирально свернутых оборотов и прямой или почти прямой трубки, тортиконы - раковины, свернутые в запутанный клубок.
Жилая камера отличается от фрагмокона отсутствием перегородок; у большинства ее длина составляет от 1/2 до одного оборота; однако у эволютных многооборотных раковин встречаются жилые камеры, достигающие 1,5-2 оборотов; короткие жилые камеры наблюдаются у форм с инволютной раковиной. Устье может быть с простым и сложным краем. Во втором случае оно может иметь боковые ушки различной формы и длины или вентральный выступ, широкий или узкий или, наоборот, может быть развит гипономический синус. Наличие последнего Позволяет предполагать присутствие у данного аммонита воронки; наличие выступа ставит под сомнение присутствие у этих аммонитов воронки. У некоторых форм наблюдается сильно суженное устье.
Раковина может быть гладкой или скульптированной. На гладких раковинах наблюдаются струйки роста, отражающие форму устьевого края. Они состоят из выступов и синусов. Как у всех моллюсков, скульптура бывает радиальной и продольной или спиральной. Радиальная скульптура представлена ребрами различных размеров и формы, главных и промежуточных, несущих бугорки и без них; продольная - выражена преимущественно продольными ребрами. Сетчатая скульптура возникает при сочетании двух первых типов. Вдоль вентральной стороны бывают развиты борозды и кили (1-3). На раковинах часто различают пережимы; они связаны с утолщением раковинного слоя и чаще выражены на внутренних ядрах. Значительный интерес представляют воротники - своеобразные приустьевые отвороты, возникавшие периодически и наблюдаемое только у мезозойских литоцератид. Параболические линии и бугорки - остатки промежуточных устьев, имевших фигурное очертание, в процессе роста заполнялись новым раковинным веществом.
Окраска раковины наблюдается очень редко и описана у немногих экземпляров; она представлена темными или светлыми полосами, параллельными устьевому краю.
Септы и лопастные линии. Протоконх отделяется от фрагмокона первой септой, или просептой. Как и все последующие септы, просепта прикреплялась к раковине изнутри; линия прикрепления ее образует первую лопастную линию, или просутуру. У первых раннедевонских аммонитов просутура была простой линией, подобной таковой наутилоидей. В. Бранко (1879-1880) назвал ее аселлатной, т. е. бесседельной, в ней выделяется маленькая умбиликальная лопасть, расположенная на шве (рис. 201). Из аселлатной линии у всех палеозойских и части триасовых аммонитов развивается латиселлатная, или широкоседельная, просутура, состоящая из широкого седла, развитого на вентральной стороне, и соответствующего ему дорзального седла; между двумя седлами с каждой стороны расположены одна или две лопасти: одна, лежащая на шве, получила название умбиликальной, а вторая, лежащая рядом с вентральным седлом, - латеральной. Из латиселлатной просутуры в триасе возникла ангустиселлатная, или узкоседельная, с узким вентральным седлом, дорзальным седлом и двумя или тремя лопастями между ними; третья лопасть получила название внутренней боковой. В процессе филогенеза аммоноидей наблюдается развитие просутуры от аселлатной к ангустиселлатной. В соответствии с формой просутуры различают три типа протоконхов: аселлатный, латиселлатный и ангустиселлатный (рис. 202). Вторая перегородка отличается от просепты возникновением на месте вентрального и дорзального седел двух новых лопастей - вентральной (V) и дорзальной (D); соответственно вторая лопастная линия, или примасутура, имеет вентральную и дорзальную лопасти и расположенные между ними одну-три лопасти; число этих лопастей зависит от их числа в просутуре. Дальнейшее развитие перегородок и лопастных линий у разных аммонитов протекало по-разному. Различают три основных типа лопастных линий (рис. 201, 4): гониатитовый, цератитовый и аммонитовый.
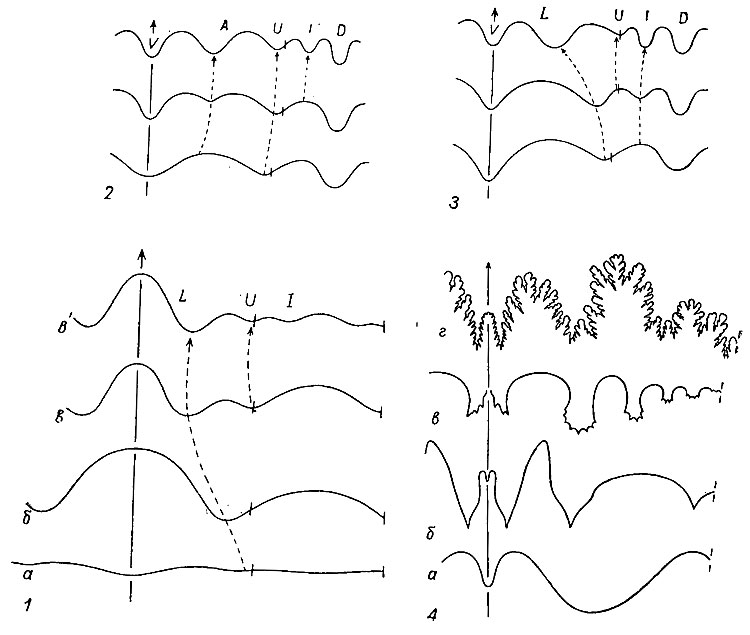
Рис. 201. Строение лопастной линии: 1 - филогения просутуры: а - аселлатная (ранний девон), б - латиселлатная (девон - триас), в, в' - ангустиселлатная (триас - мел); 2 - возникновение адвентивной лопасти (A) и 3 - умбиликальной (U); 4 - типы лопастных линий: а - агониатитовая, б - гониатитовая, в - цератитовая, г - аммонитовая; лопасти: A - адвентивная, D - дорзальная, I - внутренняя, L - латеральная, U - умбиликальная, V - вентральная
Гониатитовый тип - лопастная линия состоит из простых нерасчлененных лопастей и седел, причем, как правило, лопасти заостренные, а седла округленные.
Цератитовый тип - лопастная линия состоит из округленных седел и мелкозазубренных лопастей.
Аммонитовый тип - лопастная линия состоит из расчлененных лопастей и седел.
Иногда у девонских аммонитов выделяют агониатитовый тип - лопастная линия отличается от гониатитовой наличием очень широкой боковой (омнилатеральной) лопасти и небольшим числом элементов.
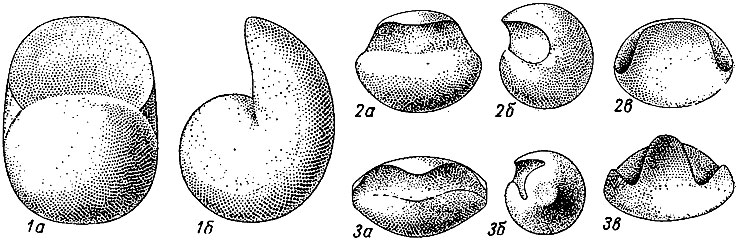
Рис. 202. Основные типы протоконха: 1 - аселлатный, 2 - латиселлатный, 3 - ангустиселлатный; а - вид со стороны первой перегородки, б - вид сбоку, в - вид с вентральной стороны
Каждая лопасть получает буквенное обозначение и индекс; с помощью обозначений и индексов можно составить формулу лопастной линии каждого аммонита. У мезозойских литодератид наблюдается особый тип строения дорзальной лопасти; от нее отходили назад две ветви задней части мантии, которые прикреплялись к предыдущей перегородке, оставляя на ее дорзальной части отпечатки, названные септальными крыльями (см. рис. 209). Изучение развития лопастной линии в онтогенезе позволяет выявлять родственные связи между аммонитами.
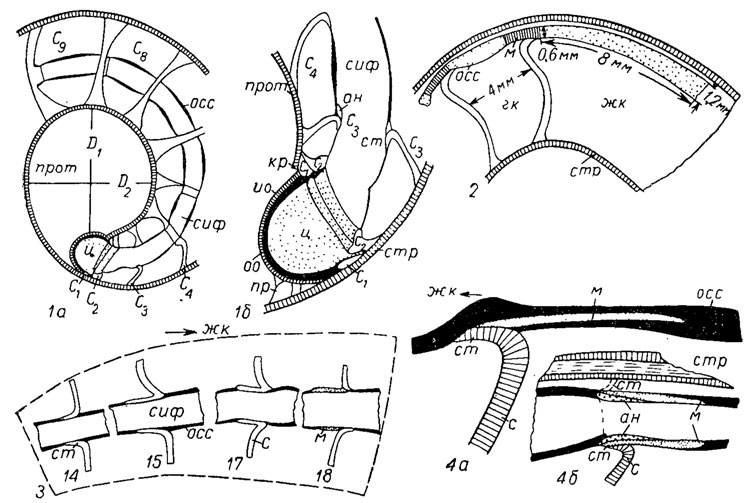
Рис. 203. Внутреннее строение филлоцератид: 1-2 - Phyllopachyceras: 1a - протоконх и начало первого оборота, 1б - увеличенная часть 1а; 2 - остатки сифона в жилой камере; 3-4 - Euphylloceras: 3 - переход от ретрахоанитовой (14) к прохоакитовой (18) септальной трубке, 4 - взаимоотношение органической оболочки сифона, манжеты и септальной трубки: 4а - стенка сифона, 4б - сифональная трубка; ан - аннулярные образования, гк - гидростатическая камера, D1, D2 - большой и малый диаметры, жк - жилая камера, ио - известковая оболочка декума, кр - кромка, м - манжета, оо - органическая оболочка цекума, осс - органический сегмент сифона, пр - просифон, прот - протоконх, C1, C2...Cn - первая, вторая и последующие септы, сиф - сифон, ст - септальная трубка, стр - стенка раковины, ц - цекум
Сифон начинается в протококхе небольшим шаровидным вздутием, названным цекумом (рис. 203). После того как тело аммонита подтянулось и освободило полость протоконха, задняя часть тела аммонита оказалась прикрепленной к стенке протоконха с помощью связки, образованной за счет эпителия мантии. Эта связка состоит из бокала, охватывающего цекум, и длинного или короткого тяжа, или ленты, или нескольких тяжей. Эта связка получила название просифона, так как исследователь, предложивший этот термин, предполагал, что просифон выполнял в протоконхе функцию сифона; в действительности он служил только для прикрепления задней части тела. После того как полость протоконха была освобождена, задняя часть мантии строила просепту, которая прикреплялась с дорзальной стороны к дистальному концу протоконха, оставляя небольшой его конец, названный кромкой; вокруг цекума выделялась тонкая известковая оболочка. Наличие в просепте двух седел - вентрального и дорзального - обусловлено существованием цекума, который служил препятствием для изгиба септы назад и образования лопасти. Но когда тело подтянулось дальше и мантия начала строить вторую перегородку, или примасепту, то между просептой и примасептой образовался тяж сифона. В примасепте для сифона возникло отверстие, что, по-видимому, обусловило появление на внутренней и наружной сторонах септы соответствующих изгибов-дорзальной и вентральной лопастей. Они сохраняются у всех последующих септ. В септах, вокруг отверстия сифона, образовались септальные трубки; у палеозойских аммонитов они направлены назад и называются ретрохоанитовыми, а у мезозойских они на взрослых оборотах направлены вперед. Септальные трубки, направленные вперед, называют прохоанитовыми. Сифон обычно снабжен соединительными кольцами, которые, по-видимому, выделялись его эпителием и служили для прочного соединения сифона с септами. Соединение было необходимо для создания герметичности камер. Сифон у аммонитов на первых оборотах может занимать любое положение - от центрального до краевого, но на всех взрослых оборотах - вентральнокраевое и только у одного отряда - позднедевонских климений - он смещен к дорзальной стенке. Относительные размеры сифона уменьшаются с возрастом: если на первых оборотах его диаметр составляет 1/3-1/4 высоты оборота, то на последних может составлять только 1/16.
Стенка раковины. Стенки протоконха и первого оборота, а также двух первых септ построены из призматического слоя. Приблизительно в конце первого оборота у всех аммонитов наблюдается резкое увеличение раковинного слоя, получившего название первичного валика; он расположен от устья протоконха на расстоянии от 270 до 360° и даже 380° (т. е. больше одного оборота). Валик обычно сопровождается первичным пережимом. Его возникновение связано с появлением перламутрового слоя. По-видимому, после того как молодой аммонит - аммонителла - вышел из яйцевых оболочек и попал в новую для него среду, ему потребовалось некоторое время для адаптации к новым условиям. Раковина аммонителлы состояла из протоконха и первого оборота. Аммонителла имела одну-две гидростатических камеры, цекум с просифоном и первый сегмент сифона. За время адаптации мантия аммонителлы приобретала способность выделять перламутровый слой, за счет которого строился первичный валик. Затем наступал новый этап в жизни аммонита, он продолжал расти, строить новые части раковины, новые перегородки. Все септы, начиная с третьей, строились за счет перламутрового слоя. После первичного валика стенка раковины всех последующих, оборотов построена из трех слоев - двух призматических и заключенного между ними перламутрового. Пережимы, или валики, которые наблюдаются на раковинах, всегда образуются за счет утолщения перламутрового слоя. Дорзальная стенка строится обычно очень экономно: только за счет внутреннего призматического слоя, остальные: слои выклиниваются у боковой стенки предшествующего оборота.
Устье раковины аммонитов после втягивания тела внутрь раковины закрывалось известковой или роговой крышкой, гомологичной: таковой гастропод. Известковые крышечки, известные под названием аптихов, образовывались эпителием капюшона, гомолога подобного образования наутилуса. Если раковина строилась из арагонита, то аптихи всегда из кальцита. Обычно аптихи встречаются отдельно от раковины и довольно часто бывает трудно установить, каким группам аммонитов они принадлежат. Поэтому их систематика, так же как систематика ринхолитов, является искусственной. Аптихи состоят из двух створок; крышечка, состоящая из одной створки, называется анаптихом. Анаптихи встречаются в ископаемом состояние в течение всего времени существования аммонитов, т. е. с девонского по меловой периоды, в то время как аптихи известны преимущественно из юрских и меловых отложений.
Основы систематики и классификация. На основании изучения изменений в онтогенезе лопастной линии, формы поперечного сечения оборотов, сифона и септальных трубок подкласс аммоноидей разделен на семь отрядов: Agoniatitida, Goniatitida, Clymeniida, Ceratitida, Phylloceratida, Lytoceratida, Ammonitida.
Отряд Agoniatitida. Агониатиты (гр. gonia - угол, a - отрицание). Раковина преимущественно спирально-плоскостная, дисковидная, или линзовидная, у ранних форм криоконная с несоприкасающимися: оборотами, позднее с соприкасающимися, но с умбиликальным отверстием и, наконец, у большинства свернутая в плотную спираль. Сифон на всех стадиях вентральнокраевой, протоконх с аселлатной и латиселлатной просутурой. Лопастная линия агониатитовая, гониатитовая и у поздних форм - цератитовая. Примасутура у ранних форм: с широкой омнилатеральной лопастью, позднее замещающейся латеральной; новые лопасти - умбиликальная и внутренняя - возникали за счет умбиликального седла. Омнилатеральная лопасть была унаследована от бактритов, от которых возникли агониатиты в середине раннего девона. Развитие отряда шло по пути увеличения размеров раковины, возникновения разнообразных морфологических типов от полностью инволютных до эволютных, усложнения лопастной линии и: возникновения новых лопастей и седел, число которых у некоторых, форм достигало 50, превращения простой вентральной лопасти в трех раздельную, простой дорзальной - в двузубчатую, сложного расчленения наружного седла и лопастей у пермских и триасовых форм. Девон-триас. Около 100 родов (рис. 204).
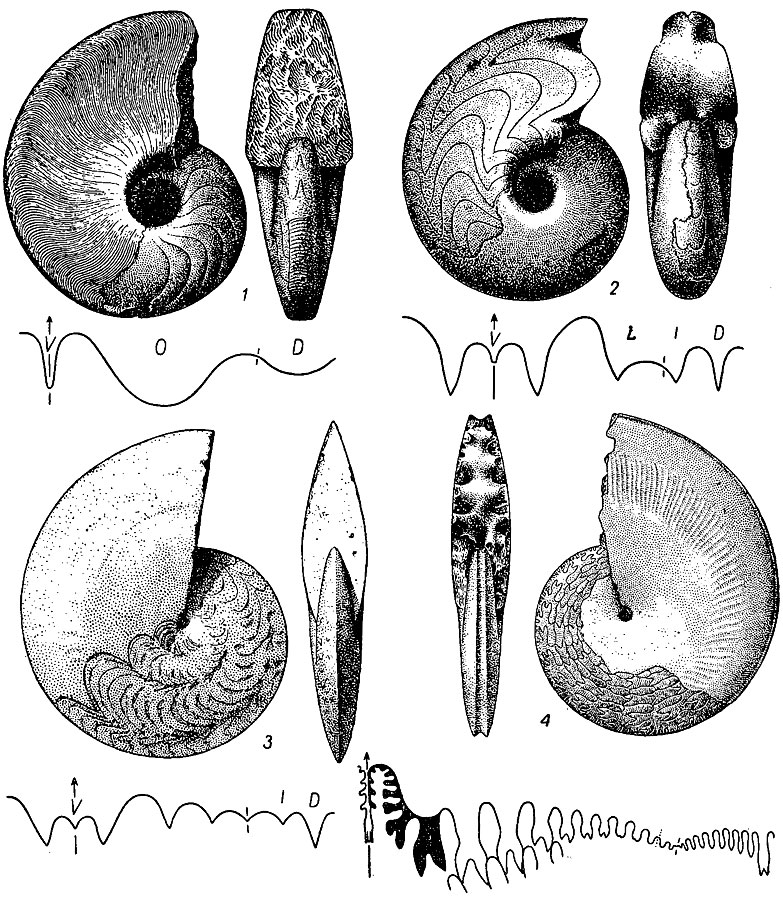
Рис. 204. Отряд Agoniatitida: 1 - Agoniatites (средний девон); 2 - Manticoceras (поздний девон); 3 - Timanites (поздний девон); 4 - Medlicottia (пермь)
Отряд Goniatitida. Гониатиты (гр. gonia - угол). Раковина разнообразной формы от зволютной до инволютной, от субсферической до дисковидной, гладкая или скульптированная, с продольной поперечной и сетчатой скульптурой. Протоконх с латиселлатной просутурой. Лопастная линия преимущественно гониатитовая, у поздних форм - дератитовая. Примасутура у ранних форм состояла из вентральной, латеральной и дорзальной лопастей. Сифон на ранних стадиях от центрального до дорзального, на всех взрослых - вентральнокраевой. Септальные трубки ретрохоанитовые, от коротких до длинных. Развитие шло по пути превращения простой вентральной лопасти в двураздельную, возникновения новой адвентивной лопасти, расположенной рядом с вентральной и внутренней боковой. Дорзальная лопасть оставалась простой или становилась двух-, реже трехзубчатой. Новые лопасти и седла возникали в результате многократного деления адвентивной, умбиликальной и внутренней боковой лопастей. Гониатиты возникли от агониатитов в среднем девоне, получили широкое развитие в карбоне и перми и в конце перми вымерли, не оставив потомков. Около 200 родов (рис. 205, 211, 4).
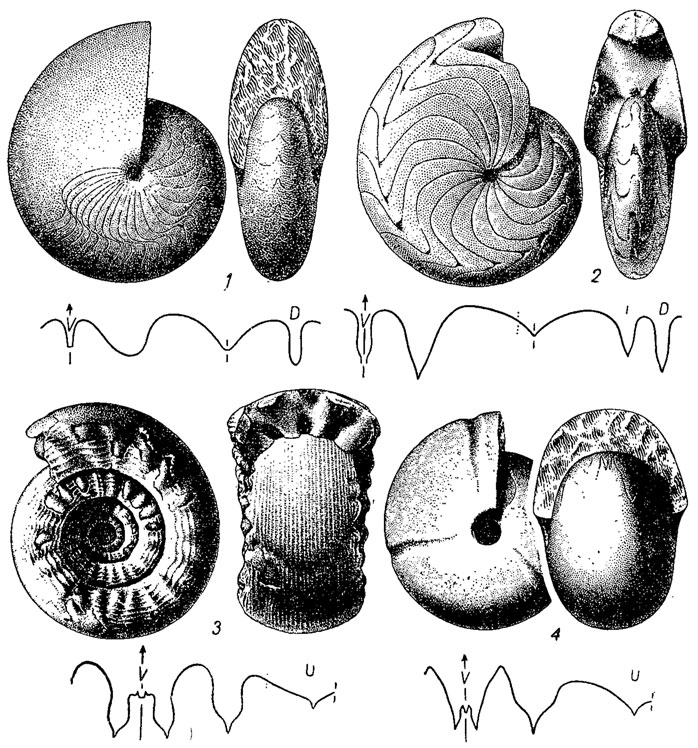
Рис. 205. Отряд Goniatitida: 1 - Tornoceras (девон); 2 - Imitoceras (поздний девон - пермь); 3 - Paragastrioceras (пермь); 4 - Goniatites (ранний карбон)
Отряд Clymeniida. Климении (гр. klimax - лестница, подчеркнуто ступенчатое строение пупка) отличаются от всех аммоноидей дорзальнокраевым положением сифона. Протоконх латиселлатный; сифон у ранних форм на первом обороте занимал вентральнокраевое положение, а затем смещался к дорзальной стороне. Сифональные трубки ретрохоанитовые, длинные. Раковины преимущественно эволютные с широким умбиликом, в одном случае с треугольным навиванием; имеются полностью инволютные, шаровидные раковины. Просутура латиселлатная; лопастная линия гониатитового типа; в отличие от всех аммоноидей у надсемейства Clymeniacea в филогенезе вначале вентральная лопасть заменяется вентральным седлом, а позднее тоже происходит с дорзальной лопастью. Климении возникли в начале позднего девона в зоне manticoceras от агониатитов (подотряд Anarcestina), стали особенно разнообразными во второй половине позднего девона (фаменском ярусе) и вымерли на границе с карбоном. Известно около 30 родов (рис. 206).
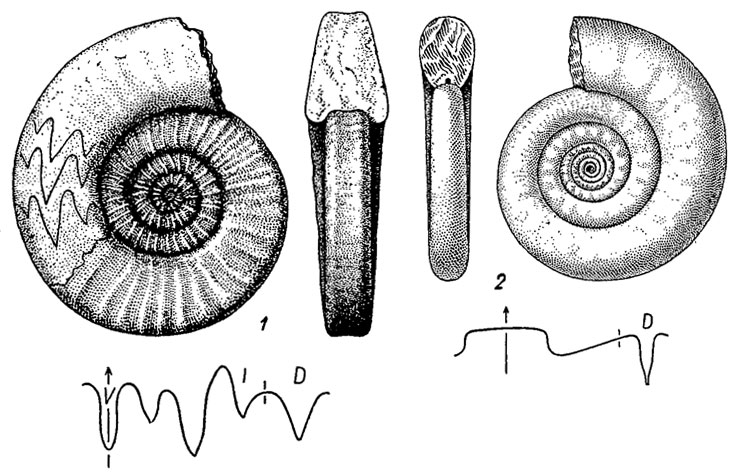
Рис. 206. Отряд Clymeniida: 1 - Gonioclymenia (поздний девон); 2 - Clymetiia (поздний девон)
Отряд Ceratitida. Цератиты (гр. keros - рог) объединяют около 400 родов, главное распространение которых относится к триасу. Первые цератиты появились в конце ранней перми, в артинском веке, отделившись от агониатитов. Протоконх с латиселлатной просутурой, имеет одну или две лопасти; на взрослых стадиях лопастная линия цератитовая или аммонитовая. Сифон в начале первого оборота занимает любое положение - от центрального до дорзально или вентральнокраевого; на последующих оборотах смещается к вентральной стенке; септальные трубки вначале ретрохоанитовые, на поздних оборотах прохоанитовые. Раковины разнообразной формы - от эволютных до инволютных, гладкие или скульптированные, с ребрами, бугорками, шипами, часто на вентральной стороне наблюдаются кили; в конце триаса появляются немногочисленные цератиты с гетероморфной раковиной - турриконовой, криоконовой или бакуликоновой. Ранняя пермь - триас (рис. 207).
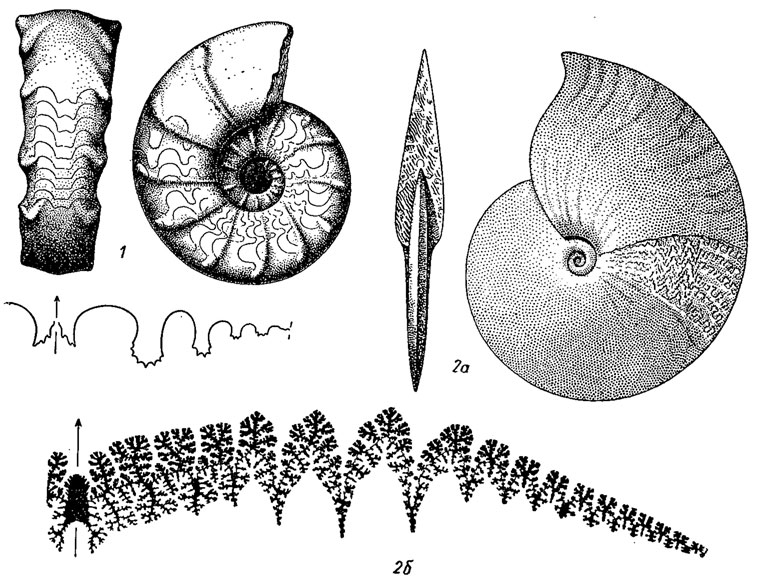
Рис. 207. Отряд Ceratitida: 1 - Cerahtes (средний триас); 2 - Pinacoceras (поздний триас): 2а - общий 2б - лопастная линия
Отряд Phylloceratida. Филлоцератиды (гр. phyllon - лист) возникли в конце перми или начале триаса от цератитов и представляют собой небольшой отряд (около 30 родов), существовавший без значительных изменений до конца мела. Протоконх шаровидный или валикообразный с ангустиселлатной просутурой. Лопастная линия взрослых, форм аммонитовая, с большим количеством лопастей я седел; седла заканчиваются округленными листочками и никогда не бывают двураздельными; вентральная лопасть разделена срединным бутылеобразным седлом на две ветви, боковая - трехраздельная, дорзальная - с прямыми нерассеченными стенками. Сифон начинается шаровидным цекумом, который прикрепляется к стенке протоконха многочисленными, но короткими тяжами (сложным просифоном); на первых двух оборотах сифон занимает почти центральное положение, а затем постепенно смещается к вентральной стенке; oсептальные трубки на первых оборотах ретрохоанитовые, а затем прохоанитовые, короткие, сопровождаются манжетами - особыми соединительными кольцами, направленными назад. Раковины преимущественно инволютные, тонкоребристые, редко груборебристые. Жилая камера занимает от 1/2 до 3/4 оборота. Пермь (?), триас - мел (рис. 203, 208).
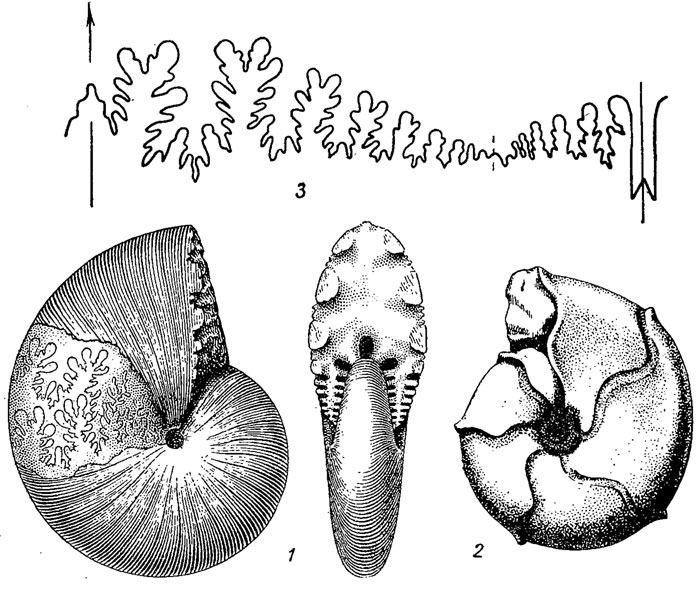
Рис. 208. Отряд Phylloceratida: 1 - Phylloceras (юра - мел); 2 - Sowerbyceras (поздняя юра); 3 - лопастная линия Salfeldiella (мел)
Отряд Lytoceratida. Литоцератиды (гр lytos - неплотный, keras - рог) возникли в начале юры от филлоцератид и дали большое количество родов и видов в юрском и меловом периодах. Протоконх валиковидный с ангустиселлатной лопастной линией. Просутура с двумя или тремя лопастями, расположенными между вентральным и дорзальным седлами. Лопастная линия взрослых форм с ограниченным числом лопастей и седел. Вентральная и боковая лопасти, а также седла - двураздельные; дорзальная лопасть крестообразная, с септальными крыльями; в вентральной лопасти вторичное седло с прямыми стенками. Сифон начинается шаровидным цекумом, который прикрепляется к протоконху, подобно таковому филлоцератид, многочисленными, но короткими тяжами и почти сразу, в отличие от филлоцератид, занимает краевое положение. Септальные трубки вначале ретрохоанитовые, а после второго оборота прохоанитовые; септальные трубки длинные, манжеты расположены внутри септальных трубок и лишь немного выступают назад. Раковины преимущественна эволютные или полуэволютные и многие гетероморфные; среди последних криоконовые, турриконовые, крючкообразные. У всех гетероморф лопастная линия состоит только из четырех лопастей: вентральной, дорзальной и двух, лежащих между ними; все лопасти и седла двураздельные. Раковины гладкие или тонкоребристые, часта с воротниками различной конструкции и размеров, от очень больших до маленьких. Юра - мел. Около 100 родов (рис. 209, 211, 1).

Рис. 209. Отряд Lytoceratida: 1 - Lytocercus (юра): 1а - раковина с воротниками, 1б - лопастная линия, 1в - септальные крылья; 2 - Pictetia (ранний мел); 3 - Hamulina (ранний мел); 4 - Turrilites (мел); 5 - Anisoceras (мел); 6 - Nipponites (поздний мел)
Отряд Ammonitida. Аммонитиды (Ammon - египетский бог Солнца) объединяют всех юрских и меловых аммоноидей, кроме филлоцератид и литоцератид (свыше 800 родов), онтогенез которых изучен во многих случаях еще недостаточно. Протоконх валикообразный с ангустиселлатной просутурой, в последней две или три лопасти. Лопастная линия взрослых форм отличается большим разнообразием - от сложно расчлененной аммонитовой до упрощенной псевдоцератитовой у некоторых позднемеловых форм. Вентральная лопасть двураздельная, латеральная трехраздельная, дорзальная сложно рассеченная. Новые лопасти возникают в области умбилика за счет расчленения умбиликального седла или лопасти. Раковины преимущественно спирально-плоскостные, от эволютных до полностью инволютных, гладкие и ребристые, с килями и без них, с бугорками, шипами; устье простое или сложное, с ушками или вентральным ростром. В юре и особенно в мелу возникает ряд аммонитид с гетероморфной раковиной - криоконовой, анцилоконовой, гетероконовой, скафиконовой и многие другие формы. Аммонитиды возникли от литоцератид в начале юры, достигли расцвета в юрском периоде (известно до 650 родов), были разнообразны и многочисленны в меловом периоде и вымерли в конце мела (в Маастрихте). Этапы развития аммонитид используются для зонального деления ярусов юры и мела (рис. 210, 211, 2, 3).

Рис. 210. Отряд Ammonitida: 1 - Stephanoceras (средняя юра); 2 - Perisphinctes (поздняя юра); 3 - Cadoceras (поздняя юра); 4 - Scaphites (мел); 5 - Cardioceras (поздняя юра); 6 - Virgatites (поздняя юра); 7 - Acanthoceras (поздний мел); 8 - Tissotia (поздний мел); 9 - Ancyloceras (ранний мел)
История развития аммоноидей. Аммоноидеи возникли в начале раннего девона, по-видимому, от бактритов, имевших прямую или слабосогнутую раковину. Наличие такой раковины чрезвычайно сужало эволюционные возможности этого подкласса. Возникновение аммоноидей было связано с образованием вначале согнутой, а затем криоконовой раковины, состоявшей из несоприкасающихся оборотов свернутых в одной плоскости. Появление планоспиральной раковины имело большие эволюционные последствия; раковина стала более компактной, удобной для маневрирования, тонкая апикальная часть трубки оказалась защищенной последующими оборотами - все эта значительно расширило эволюционные возможности аммоноидей и привело их к адаптациям к самым разнообразным условиям существования, подобно тому как возникновение планоспиральной раковины у гастропод обеспечило. их эволюцию. Первые агониатиты имели раковину криоконового типа, с протоконхом аселлатного типа. Затем возникли планоспиральные раковины вначале с умбиликальным отверстием, потом без него, с протоконхом латиселлатного типа. Па особенностям развития лопастной линии наметились две параллельна развивающиеся группы - агониатиты и гониатиты (рис. 217). Основное различие между ними сводится к развитию латеральной лопасти, которая у агониатитов развивается за счет омнилатеральной, унаследованной от бактритов и вначале сохраняющейся у первых агониатитов; новые лопасти развиваются в области умбилика за счет расчленения лопасти или седла. У гониатитов, возникших от агониатитов в среднем девоне, латеральная лопасть остается маленькой в области умбилика, а новые лопасти, так называемые адвентивные, возникают за счет седла, лежащего между латеральной и вентральной лопастями, внешний вид которых на взрослой стадии не отличим от настоящей латеральной. От агониатитов в начале позднего девона возникает небольшой отряд климений, отличающийся дорзальным положением сифона и развитием иногда вместо вентральной и дорзальной лопастей соответствующих седел. Климении вымирают на границе девона и карбона.
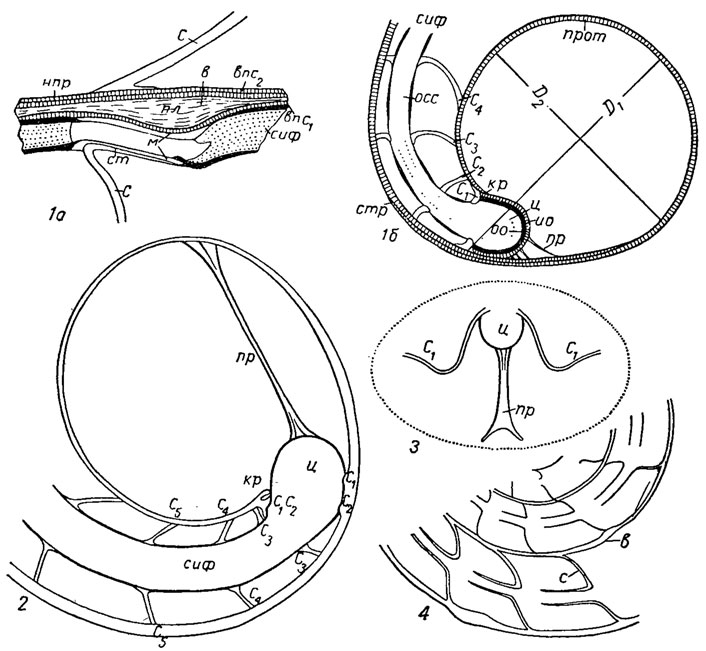
Рис. 211. Внутреннее строение раковины литоцератид (1), аммонитид (2-3) и гониатитов (4): 1 - Tetragonites: 1а - взаимоотношение органической оболочки сифона, манжеты и септальной трубки, 1б - протоконх и начало первого оборота; 2 - Sphaeroceras, протоконх и начало первого оборота; 3 - Parkinsonia, протоконх и первая септа; 4 - Agathiceras, часть продольного сечения; в - валик, впс1, впс2 - внутренний призматический слой одного оборота и следующего, нпр - наружный призматический слой, пл - пластинчатый, или перламутровый, слой. Остальные обозначения те же, что на рис. 203
В конце ранней перми от агониатитов развивается основной ствол триасовых аммонитов - цератиты, имевшие протоконх с латиселлатной, а в конце их существования - с ангустиселлатной просутурой. От них в конце перми или начале триаса возникают филлоцератиды - консервативный ствол медленно эволюционирующих аммоноидей, но чрезвычайно важный в филогении аммоноидей: от него в начале юры возникли литоцератиды, а от последних - собственно аммониты (отряд аммонитид). Вначале цератиты были представлены только одним родом, в поздней перми значение этой группы постепенно возрастало и в конце перми она заняла ведущее положение, полностью вытеснив гониатитов и частично агониатитов. В раннем триасе цератиты широко распространяются по всем морям земного шара, возникают новые группы разного таксономического ранга. В конце триаса произошло вымирание цератит; перед их массовым вымиранием появились немногие цератиты с гетероморфной раковиной (криоконовой, турриконовой и бакуликоновой); часть цератитов приспособилась к новым условиям жизни. Общее число родов аммонитов, обитавших в триасе, приближается к 400. Среди этой цифры несколько родов филлоцератид. В конце триаса (рэтском ярусе) от большого отряда цератит сохраняется немногим более 5 родов. Раннеюрский этап характеризуется полным обновлением систематического состава; от филлоцератид возникают литоцератиды и от последних аммонитиды. Если филлоцератиды и литоцератиды в юрское время были распространены только в средиземноморской области, то аммонитиды образуют характерные группировки для средиземноморского и бореального поясов, часто по составу совершенно различные и неповторимые. Аммонитиды дают в юре небольшое число родов с гетероморфной раковиной (надсемейство Spirocerataceae).
Многие аммонитиды в юрском и меловом периодах были очень широко распространены, быстро эволюционировали и используются для зонального деления юрских и меловых отложений. В конце юры происходит почти полное обновление родового состава среди аммонитид. Но филлоцератиды продолжают свое существование без особенно крупных изменений, в то же время наблюдается адаптивная радиация среди литоцератид и аммонитид. Среди последних двух отрядов в меловом периоде возникали многочисленные роды с гетероморфной раковиной, указывающие на приспособление к самым разнообразным условиям жизни. Литоцератиды с турриконовой раковиной (туррилитиды), по-видимому, приспособились к бентосному подвижному образу жизни, с клубкообразной раковиной (Nipponites) были, вероятно, малоподвижны, с крючкообразной раковиной (Ptychoceras, Hamites) вели нектонно-бентосный образ жизни. Кроме перечисленных известны формы с криоконовой, анцилоконовой, скаoфиконовой, бакуликоновой и другими типами раковин. Особенный интерес представляют гетероцератиды, которые на ранней стадии имели турриконовую раковину и, вероятно, ползали по дну, а затем изменяли образ жизни и приобретали выпрямленную раковину с крючком на конце. В конце мела, в Маастрихте, все три отряда аммоноидей (филлоцератиды, литоцератиды и аммонитиды) полностью вымирают.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'