
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Подкласс Coleoidea. Колеоидеи
К подклассу колеоидей (гр. koleos - футляр, ножны) относятся все, за исключением наутилуса, современные головоногие (рис. 212), насчитывающие до 650 видов и большое число вымерших родов и видов; среди них можно назвать белемнитов, кальмаров, каракатиц, осьминогов. Все они пелагические или придонные хищники, ведущие нектонный, нектонно-бентосный и частично планктонный образ жизни. Тело мешковидное или веретеновидное, самых разнообразных размеров - от нескольких сантиметров до 18 м, часто снабженное плавниками. В отличие от современного наутилуса колеоидеи имеют только две жабры (отсюда название, двужаберные, или дибранхиаты), две почки, главное сердце, состоящее из желудочка и двух предсердий, и два венозных, или жаберных, сердца, накачивающих кровь в жаберные сосуды, два женских половых отверстия или одно мужское, парные протоки нидаментальной железы. Голова несет восемь или десять крупных щупалец, или рук, снабженных роговыми крючочками или присосками; у десятиногих две руки длиннее остальных восьми, они называются ловчими. Внутри кольца рук имеется, буккальная мембрана, состоящая из 7-10 своеобразных выростов. Рот вооружен парой роговых челюстей и радулой, несущей 7-9 рядов хитиновых зубчиков. В задней части тела у большинства имеется чернильный мешок, сохраняющийся в ископаемом состоянии. Лопасти воронки срослись в цельную трубку, и мантийная щель замыкается особым замком, построенным по типу запонок. Нервные узлы сконцентрированы в единую компактную массу, заключенную в хрящевую капсулу, напоминающую хрящевой череп позвоночных. Высоко развиты органы чувств, особенно глаза, максимальные размеры которых достигают 30 см. Хрящевые образования имеются около глаз, у основания рук, около плавников и в других местах. Под кожей развиты особые клетки - хроматофоры, содержащие различные пигменты, окрашивающие кожу в защитные цвета.
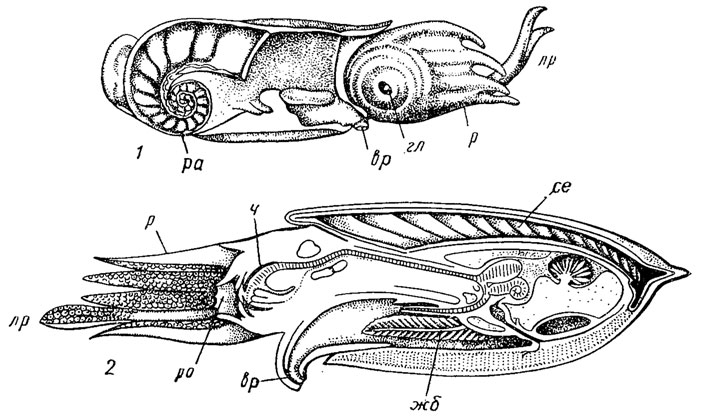
Рис. 212. Подкласс Coleoidea. Схема строения современных Spirula (1) и Sepia (2): вр - воронка, гл - глаза, жб - жабры, лр - ловчие руки, р - руки, ра - раковина, ро - ротовое отверстие, се - сепион, ч - челюсти
Колеоидеи отличаются от всех вышерассмотренных подклассов наличием внутренней раковины, с различной степенью редукции, вплоть до полного ее отсутствия (отсюда еще одно название этого подкласса - эндокохлии, или внутреннераковинные). Раковина может состоять: а) из фрагмокона, проострака и ростра, б) из фрагмокона, жилой камеры и телума, в) из видоизмененного проострака, или гладиуса, г) из видоизмененного фрагмокона - сепиона, д) полностью отсутствовать. Наконец, у одной группы колеоидей (аргонавты) самки за счет эпителия рук образуют специальную раковину для вынашивания оплодотворенных яиц, очень напоминающую раковину аммонитов, но без перегородок. Развитие всех современных колеоидей прямое, без личиночных стадий. Яйца заключены в сравнительно толстую капсулу и кладки яиц обычно прикрепляются самкой к различным подводным предметам.
Колеоидеи, как отмечалось выше, возникли от бактритоидей в раннем карбоне и вначале развивались сравнительно медленно, но после того как раковина превратилась полностью во внутренний скелет и тело приобрело торпедовидное очертание и плавники, колеоидеи образовали одну из наиболее распространенных и высокоорганизованных групп среди всех беспозвоночных.
На основании особенностей строения скелета и мягкого тела подкласс колеоидей разделен на пять отрядов: Aulacoceratida, Belemnitida, Teuthida, Sepiida и Octopodida.
Отряд Aulacoceratida. Аулакоцератиды (rp. aulax, род. aulaxos - борозда). Скелет состоит из фрагмокона, жилой камеры и телума. Жилая камера длинная, трубчатая, с перистомом, имеющим на вентральной и дорзальной сторонах по языкообразному выступу, подобно таковому у одного рода бактритов (Pseudobactrites). Жилая камера переходит в узкоконический фрагмокон (угол от 5 до 12°, как исключение 15-20°), состоящий из многочисленных гидростатических камер, разделенных септами с прямой перегородочной линией. Протоконх шаровидной формы, отделен от фрагмокона тонкой мембраной, цекум и просифон не установлены. Сифон занимает краевое положение; септальные трубки прохоанитовые; соединительные кольца связаны с внутренней поверхностью септальной трубки и направлены вперед и назад. Фрагмокон заключен в телум, по форме напоминающий ростр белемнитов, но отличающийся от последнего органическим составом; телум построен из слоев органического вещества (конхиолина), чередующихся с прослоями карбоната кальция; у одной группы (хитинотеутид) телум целиком состоит из конхиолина. Аулакоцератиды появились в раннем карбоне (возможно, в позднем девоне) и вымерли в конце юры. Отряд включает около 10 родов (рис. 213).
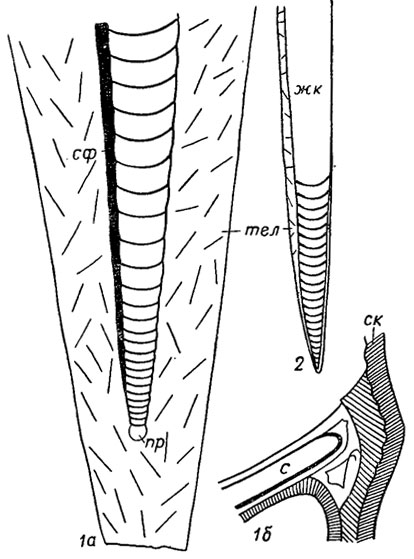
Рис. 213. Отряд Aulacoceratida. Схема строения скелета (Елецкий, 1966): 1 - Metabelemnites (поздний триас, Канада): 1а - продольный разрез, 1б - строение септы и стенки кон отеки; 2 - Aulacoceras (поздняя пермь), общий вид; жк - жилая камера, пр - протоконх, с - септа, ск - стенка конотеки, сф - сифон, тел - телум
Отряд Belemnitida. Белемниты (гр. belemnon - дротик, копье) образуют наиболее важный отряд колеоидей, возникший в карбоне, давший большое число родов и видов в юре и мелу и почти полностью вымерший к концу мела, за исключением одной небольшой реликтовой группы, дожившей до эоцена. В отличие от аулакоцератид белемниты имели скелет, состоявший из фрагмокона, проострака и известкового ростра, расположенного внутри тела на его дорзальной стороне (рис. 214). Тело, судя по отпечаткам, имело удлиненную цилиндрическую форму и обладало, как минимум, 8, а скорее 10 руками, снабженными двумя рядами роговых крючков каждая; у них не установлены ловчие руки, известные у современных кальмаров; по-видимому, они не обладали еще подобными дифференцированными руками. Фрагмокон с апикальным углом от 12 до 32° по строению напоминает раковину бактритов и имеет коническую форму; он разделен септами на отдельные гидростатические камеры. Стенка фрагмокона носит название конотеки. Протоконх полусферической формы, отделен резким пережимом от остальных камер; устьевой край закрыт сплошной вогнутой мембраной, которая, по-видимому, является просептой; стенка протоконха состоит из двух тонких призматических слоев; снаружи протоконх окружен перламутровым слоем, переходящим в наружный слой конотеки.
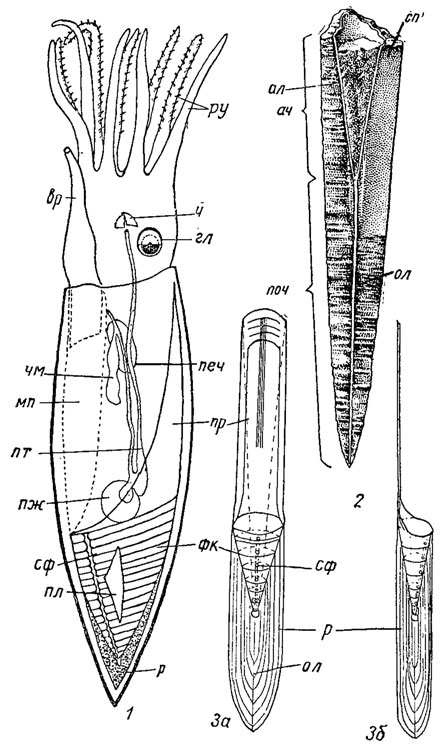
Рис. 214. Отряд Belemnitida: 1 - Belemnoteuthis syriaca из позднего мела Сирии, реконструкция (Роже, 1944); 2 - продольный разрез ростра; 3 - схема строения скелета; ал - альвеола, ач - альвеолярная часть ростра, вр - воронка, гл - глаза, мп - мантийная полость, ол - осевая линия ростра, печ - печень, пж - половая железа, пл - плавник, поч - постальвеолярная часть ростра, пр - проострак, пт - пищеварительный тракт, р - ростр, ру - руки с крючочками, сп - спайка, сф - сифон, фк - фрагмокон, ч - челюсть, чм - чернильный мешок
В отличие от аммоноидей, у которых после освобождения полости протоконха возникал просифон и соответственно цекум, у белемнитов тело прикреплялось вдоль замыкающей мембраны и поэтому сифон начинается в первой камере грибообразным расширением, которым он прикрепляется к замыкающей мембране; это расширение сифона гомологично цекуму; вторая септа без септальных трубок, ахоанитовая, через ее отверстие проходит сифон, окруженный оболочкой. В третьей и остальных септах септальные трубки ретрохоанитовые, короткие; септы трехслойные, состоят из перламутрового слоя и двух органических. Вначале сифон расположен эксцентрично, но уже между 4-6 камерами смещается к краю и между 25-30 прилегает к вентральной стенке. Конотека, гомологичная стенке раковины остальных цефалопод, состоит из двух слоев, из которых наружный, по-видимому, перламутровый, содержит большое количества органики, а внутренний призматический. Поверхность конотеки гладкая или ребристая, иногда волнистая. Дорзальный край конотеки у белемнитов образует лопатовидной формы выступ, или проострак, который очень редко сохраняется в ископаемом состоянии. Проострак гомологичен гладиусу современных кальмаров. Ряд исследователей предполагают, что проострак представляет собой дорзальную стенку жилой камеры, брюшная часть которой редуцировалась. У белемнитов она, вероятно, выполняла защитную функцию и служила местом прикрепления ряда мускулов.
В гидростатических камерах, начиная с третьей и кончая 13-20, у преимущественно юрских белемнитов наблюдаются камерные отложения, коррелятивно связанные с формой тонкого, узкоконического, относительно легкого ювенильного ростра. У геологически более молодых белемнитов имелся воронковидный более тяжелый эмбриональный ростр и камерные отложения отсутствовали.
Ростр - часть скелета белемнитов, гомологичная телуму аулакоцератид, чаще всего сохраняется в ископаемом состоянии. Ростр построен из концентрических слоев кальцита, лежащих среди органической основы. Поэтому ростры обычно окрашены в коричневые цвета различных оттенков. В онтогенезе вначале возникает эмбриональный ростр, имеющий разную форму: тонкую, узкоконическую, стреловидную или клиновидную, воронковидную; не исключена возможность, что молодой белемнит, только что покинувший яйцевые оболочки, имел протоконх и эмбриональный ростр; последний играл роль своеобразного органа равновесия у активно плавающего молодого белемнита. Ростр взрослого белемнита имеет очень разнообразную форму и состоит из чередующихся призматических и перламутровых слоев. В передней части ростра находится углубление - альвеола, в которой помещался фрагмокон; размеры и глубина альвеолы очень различны и зависят от размеров фрагмокона. Часто края альвеолы разрушаются и не отвечают первичному строению. От конца альвеолы через весь ростр к его заднему концу проходит осевая линия. Поперечное сечение ростра также очень разнообразно: от круглого до угловатого, но преобладает круглое и овальное; на поперечных срезах хорошо видны концентрические кольца, отражающие форму сечения ростра на разных возрастных стадиях; изменение формы ростра с возрастом можно также проследить на продольных разрезах.
На наружной поверхности ростра имеются продольные борозды, а также наблюдаются отпечатки кровеносных сосудов. Две борозды, расположенные по бокам, соответствуют местам прикрепления боковых плавников. Бороздки на конце ростра - дорзальная и вентральная- могут быть короткими и длинными. В приальвеолярной части также бывают бороздки - вентральная или дорзальная. Чаще всего ростры раскалываются вдоль вентральной борозды; у некоторых форм, между ней и альвеолой существует спайка - гладкая плоскость вдоль линии раскола, напоминающая спайность кристаллов; у позднемеловых белемнителл на месте спайки развивается вентральная щель. Спайка и щель отражают какие-то особенности строения мягкого тела белемнитов.
Ростр выполнял, по-видимому, три функции: служил противовесом подъемной силе фрагмокона, защищал фрагмокон и служил для усиления прочности тела.
В основу выделения подотрядов положены такие признаки, как наличие или отсутствие продольного альвеолярного канала и связанных с ним спайки или альвеолярной щели: у подотряда Belemnitina они отсутствуют, у подотряда Belemnopseina имеются либо спайка, либо альвеолярная щель; по особенностям строения ростра и проострака выделен подотряд Diplobelina с необычно узким проостраком, толстым, коротким ростром, без радиальной призматической структуры, столь характерной для двух предыдущих подотрядов.
Белемниты появились в начале карбона, до сих пор неизвестны в перми, редки в триасе, очень многочисленны и разнообразны в юре и мелу; редкие находки известны из эоцена. Отряд объединяет около 2500 видов (рис. 215).
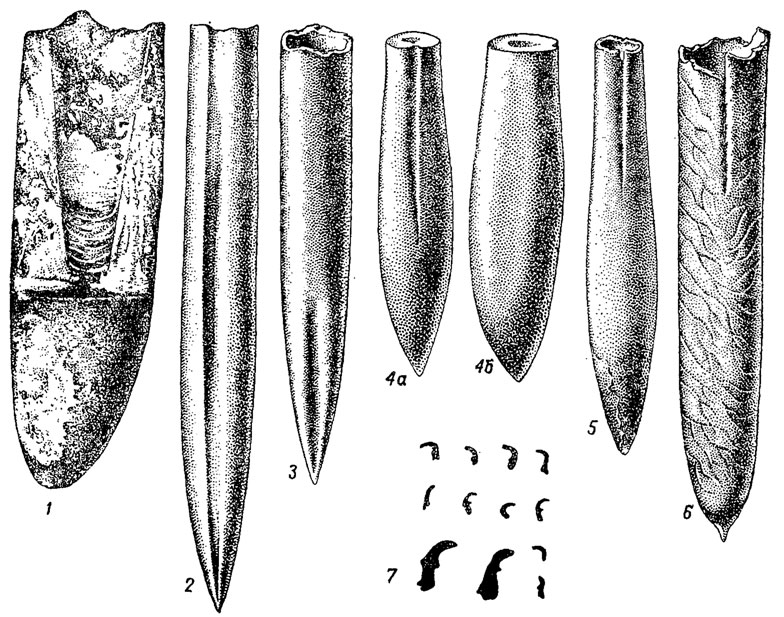
Рис. 215. Отряд Belemnitida. Ростры (1-6) и онихиты (7): 1 - Paleoceras (ранний карбон); 2 - Cylindroteuthis (юра); 3 - Pachyteuthis (поздняя юра); 4а, б - Duvalia (поздняя юра - ранний мел); 5 - Neohibolites (мел); 6 - Belemnitella (поздний мел); 7 - онихиты - конхиолиновые крючочки
Отряд Teuthida. Теутиды, или кальмары. К теутидам (гр. teuthis - кальмар) относятся современные кальмары, обладающие 10 руками, из которых одна пара ловчих, и небольшая группа глубоководных вампироморф, ранее относившихся к октоподам. По мнению Ю. А. Елецкого (1966), несмотря на восемь рук, эта группа обладает многими признаками, сближающими их с теутидами. Руки несут два-четыре продольных ряда присосок на стебельках, снабженных опорными роговыми кольцами, которые превращены у многих в крючочки (онихиты); у вампироморф имеется один ряд присосок и справа и слева от них по чувствительному усику; щупальца у них соединены перепонкой. Тело кальмаров удлиненное, торпедовидное, с ромбовидными, весловидными плавниками по бокам. Раковина имеет вид тонкой пластинки - гладиуса, гомологичного проостраку белемнитов (рис. 216,4-5). Гладиус - роговой или частично обызвествленный, разделен на три части - осевую и две боковых. У первых представителей на заднем конце сохраняется короткий фрагмокон с сифоном и маленьким ростром. У последующих фрагмокон и ростр редуцируются и сохраняются лишь остатки конотеки в виде конической или ложкообразной воронки, без перегородок. Осевая часть гладиуса продольными линиями разделена на несколько полей; латеральные части очень варьируют по размерам, форме и обычна отделяются от срединного поля (рис. 216, 4). У ископаемых форм иногда сохраняется чернильный мешок, столь характерный для всех современных кальмаров (рис. 216, 5).

Рис. 216. Отряды Sepiida (1-2), Teuthida (3-5) и Octopodida (5): 1 - Sepia, сепион (совр.); 2 - Spirula, раковина (совр.); 3 - онихиты - крючочки рук ископаемых кальмаров; 4 - Geopeltis, гладиус (юра); 5 - Parabelo- peltis с сохранившимся чернильным мешком (юра Поволжья); 6 - Argonautaargo L., самка (совр.); бп - боковое поле, п - перегородки, пк - первая камера, рр - рудимент ростра, сп - срединное поле, сф - сифон, чм - чернильный мешок
В отряд теутид включены фрагмотеутиды (рассматриваемые Елецким в качестве самостоятельного отряда), существовавшие от перми до триаса и обладавшие фрагмоконом и редуцированным ростром. Они рассматриваются в качестве боковой ветви теутид. Настоящие теутиды известны в ископаемом состоянии с поздней юры. Значительный интерес представляет один из современных кальмаров (род. Moroteuthis), у которого имеется рудимент ростра, напоминающий, по форме ростр белемнитов, но построенный из эластичного хрящевидного вещества, образующего концентрические слои; строение этого ростра напоминает велум аулакоцератид. Это послужило основанием, для предположения, высказанного Ю. К. Кабановым, о возможной прижизненной эластичности ростра белемнитов; ростр, по мнению этого исследователя, обызвествлялся посмертно. Однако это предположение. не является пока общепринятым. Известно свыше 20 родов (рис. 216,5-5).
Отряд Sepiida. К сепиидам (гр. и лат. Sepia - каракатица) относятся каракатицы, спирулы, аргонавты и ряд вымерших форм, известных в ископаемом состоянии. Тело современных сепиид сжато в дорзовентральном направлении, окаймлено плавниками; вокруг рта расположено кольцо из 10 коротких щупалец-рук, из которых одна пара: ловчих, обычно втянутых около головы в особые карманы; присоски на стебельках с роговыми кольцами, крючочков нет. Скелет современных сепиид трех типов. У одних (спирул) он представлен маленькой: спирально эндогастрически свернутой раковиной, расположенной внутри тела на спинной стороне. Раковина имеет протоконх, просифон и цекум; спиральная часть (2,5 оборота) простыми перегородками разделена на 30-40 гидростатических камер; сифон расположен: вдоль внутренней стороны оборотов; ростр и проострак отсутствуют. У других (собственно сепиид) раковина имеет вид широкой и довольно толстой овальной, пористой пластинки, целиком скрытой под спинной частью мантии, выполняющей защитную функцию. Эта пластинка, или сепион, является видоизмененным фрагмоконом предков сепиид. Третий тип скелета, установленный у позднемеловых сепиид (семейство Groenlandibelidae, выделенное Елецким), имел прямой длинноконический фрагмокон, начинающийся протоконхом, и длинный узкий проострак; сифон начинается цекумом, который прикреплен к протоконху просифоном; сифон занимает вентральнокраевое положение; септальные трубки от ахоанитовых до коротких ортохоанитовых. От подобного типа раковины можно проследить переходные формы R двум вышеописанным типам. В первом случае (спирулы) наблюдается постепенное эндогастрическое закручивание фрагмокона (рис. 216, 2), постепенное уменьшение ростра и редукция проострака, приводящее в конце концов к их полной редукции и развитию планоспиральной раковины. Во втором случае, при образовании сепиона редукции подвергались ростр и брюшная часть фрагмокона (рис. 216, 1); септы на дорзальной стороне постепенно удлинялись и приняли наклонное положение, фрагмокон был уплощен в дорзовентральном направлении, а брюшная часть септ уменьшалась и, наконец, полностью исчезла; возник сепион, внутренняя полость которого разделена косонаправленными сближенными септами, представляющими собой тонкие, слегка обызвествленные, конхиолиновые пластинки, соединенные поперечными перекладинами; сзади в виде маленького шипика иногда сохраняется маленький ростр. Сепииды встречаются в ископаемом состоянии редко, начиная с юры. Известно свыше 20 родов (рис. 216, 1-2).
Отряд Octopodida. Октоподиды, или осьминоги (лат. octo - восемь). К октоподидам относятся осьминоги, имеющие в отличие от сепиид восемь щупалец. Тело мешковидное, короткое, щупальца с присосками; раковина обычно отсутствует; у современного рода Argonauta самка значительно крупнее самца, она несет тонкостенную экзогастрически свернутую известковую раковину, лишенную камер, - образованную особой железой, расположенной на дорзальной стороне обеих щупалец. Раковина служит для вынашивания оплодотворенных яиц и молоди, она не гомологична таковой остальных цефалопод. Чернильный мешок у современных форм отсутствует, у позднемелового осьминога (см. рис. 2, 4), отпечаток тела которого (длиной 4,5 см без щупалец) обнаружен в верхнемеловых отложениях Сирии, сохранились остатки чернильного мешка, челюстей и рудиментарной раковины. Мел - ныне. Около десяти родов (рис. 216, 6).
История развития колеоидей. Геологическая история колеоидей до последних лет представлялась в следующем виде: от бактритоидей в конце девона или начале карбона возникли первые белемнитиды, имевшие фрагмокон, проострак и ростр; от них в процессе эволюции, которая шла по пути редукции раковины, возникли три дивергентные группы: теутиды, сепииды и октоподиды; у первых сохранился проострак и редуцировались фрагмокон и ростр, у вторых сохранился видоизмененный фрагмокон и отсутствуют ростр и проостраки, наконец, у третьих скелет редуцировался почти полностью.
В последние годы, благодаря исследованиям канадского палеонтолога Ю. А. Елецкого (1966), в филогению и систематику колеоидей внесены существенные изменения. По мнению этого исследователя, от бактритоидей возникли сразу две ветви: одна ветвь дала аулакоцератид с велумом, фрагмоконом и жилой камерой, вторая - белемнитид с известковым ростром, фрагмоконом и проостраком. От общего ствола с белемнитидами возникли теутиды, сепииды и октоподиды (рис. 217).
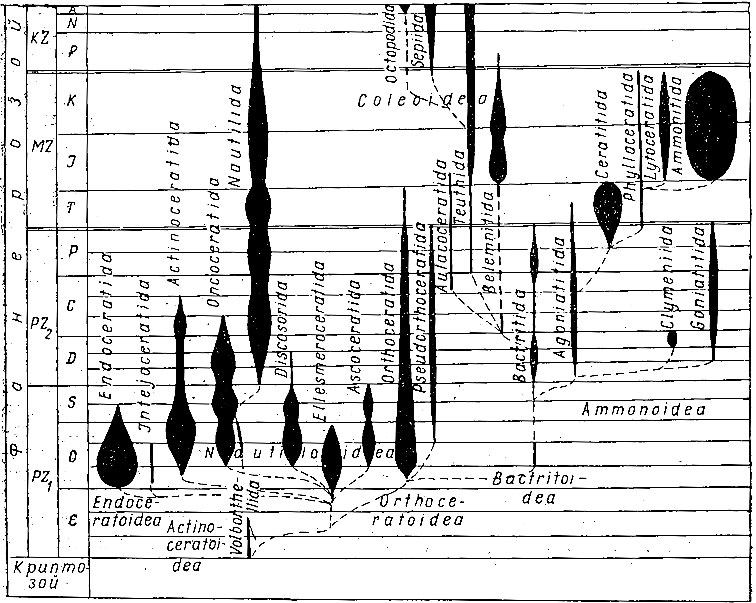
Рис. 217. Геохронологическое распространение головоногих
Сепииды и октоподиды, вероятно, ответвились от общего ствола с теутидами в начале мезозоя. У теутид и сепиид имеется много общих черт, говорящих о их общем происхождении: наличие ловчих щупалец и буккальной мембраны, присоски на стебельках с роговыми кольцами, замок по типу хрящевых кнопок для замыкания мантийной щели, нидаментальные железы, практически замкнутая кровеносная система, без лакун, сходное строение жабр и сердца. Однако сепииды отличаются от теутид по строению скелета. Особый интерес представляет строение раковины современной Spirula и мелового рода Groenlandibelus, имеющих протоконх с просифоном и цекумом, напоминающим такие же структуры у аммоноидей; как уже отмечалось, подобные структуры неизвестны ни у аулакоцератид, ни у белемнитид, ни у теутид. Вероятно, это может служить основанием для предположения о более раннем возникновении сепиид от общего предка - бактритоидей. Последний род отнесен к отдельному семейству Groenlandibelidae (поздний мел). Особенности строения ростра и прот острака послужили вначале основанием к отнесению этих колеоидей к отряду белемнитид; но по особенностям строения скелета их рассматривают в качестве боковой ветви сепиид с сохранившимися чертами предков. Возможно, от общих предков с ними развились две ветви сепиид, рассмотренные выше.
Октоподиды представляют особую ветвь колеоидей, которая возникла от общего ствола с теутидами; их развитие пошло по пути полной редукции раковины, с сохранением иногда двух хрящевых палочек, лежащих под мантией, прогрессивного развития нервной системы, концентрации ганглиев в компактную массу ("мозг"), с одновременным наличием ряда примитивных черт, таких как веслообразные плавники, зоб, наличие присосок без стебельков, без роговых колец и крючков, одно- и двухрядное расположение присосок. В современной фауне колеоидеи, таким образом, представлены тремя отрядами - теутидами, сепиидами и октоподидами - обитателями Преимущественно теплых морей всех глубин, от зоны мелководья до ультра абиссали, общее число видов которых составляет около 750.
Ринхолиты
Ринхолиты (гр. rhynchos - клюв, lithos - камень) представляют собой обызвествленные кончики челюстей вымерших головоногих, систематическая принадлежность которых обычно устанавливается с большим трудом. Ринхолит (см. рис. 196, 5) состоит из передней части - капюшона и задней - рукоятки. Капюшон - треугольный ромбовидный или стреловидный; рукоятка - либо почти прямоугольная с выпуклой верхней поверхностью, либо треугольная, с бороздкой посредине. Рукоятка, по-видимому, была скрыта в челюсти, а капюшон выставлялся из роговой челюсти наружу и служил головоногим моллюскам для прокусывания добычи, отрыва кусков или раздавливания твердых скелетов.
В ископаемом состоянии ринхолиты известны в отложениях от конца карбона до середины палеогена (эоцена). Особенно часто остатки ринхолитов встречаются в отложениях среднего триаса, юры и мела средиземноморской области. Сравнительно недавно ринхолиты были впервые описаны из меловых и эоценовых отложений Северной Америки. Из керна скважины, пробуренной на дне Атлантического океана из нижнего мела с глубины 5323 м, были извлечены три маленьких ринхолита длиной до 4 мм. И, наконец, из нерасчлененных отложений карбона - перми Южной Америки был описан конхиолиновый ринхолит, найденный в жилой камере аммонита Eoasianites.
Ринхолиты, как правило, встречаются отдельно от раковины и установить их принадлежность к соответствующей группе головоногих моллюсков - аммоноидеям, наутилоидеям или колеоидеям очень трудно. Различают две основные группы ринхолитов: Phyncholithes (5 родов) и Rhynchoteuthis (7 родов). Представители первой группы, вероятно, представляли челюсти наутилоидей. Челюсти современного наутилуса принадлежат роду Rhyncolite. Ринхолиты группы Rhynchoteuthis принадлежали другим головоногим (возможно аммоноидеям), они более многочисленны и разнообразны и могут быть использованы для стратиграфических целей при изучении юрских и меловых отложений средиземноморской области; за пределами этой области они пока неизвестны.
При описании ринхолитов, так же как и других частей скелета (например, аптихов), пользуются искусственной систематикой, выделяя формальные роды и виды, основанные на особенностях строения и формы капюшона и рукоятки, на их взаимоотношениях и относительных размерах; известно около 15 родов.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'