
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Часть первая. Принципы и направления морфологических и функциональных исследований
1.1. Морфологические исследования в палеонтологии (Э. И. Воробьева, С. В. Мейен)
Морфологические исследования составляют основу практически всех разделов и направлений развития палеонтологии (систематики, филогенетики, палеоэкологии, палеобиогеографии и др.). Более того, прогресс ни одной другой биологической дисциплины не зависит столь сильно от уровня культуры и методологии морфологических исследований. В то же время ни один из разделов морфологии рецентных форм не имеет столь тесных связей с эволюционной теорией, какие имеет морфологическая часть палеонтологии: большинство закономерностей макроэволюции и видообразования установлены в результате изучения морфологических особенностей ископаемых организмов.
Все это объясняется во многом уникальностью палеонтологического материала. Во-первых, ископаемые организмы доступны для изучения преимущественно (либо исключительно) со стороны их морфологических признаков. Во-вторых, на палеонтологических рядах форм (рядах, связанных морфологической и геохронологической последовательностью) особенно отчетливо удается проследить действительные филогенетические преобразования и направленность эволюции той или иной группы организмов и выяснить генезис доступных для изучения морфологических структур. В-третьих, палеонтологический материал, отражая в морфологических характеристиках организмов временной аспект их истории, помогает вскрыть общие закономерности и результаты эволюционного процесса и подойти к его механизмам.
Морфология решает статические и динамические задачи [187]. В первом случае она описывает и объясняет структурное многообразие организмов в определенном временном срезе, во втором - отражает формообразовательные процессы, т. е. онто- и филогенетическое преобразование структуры. Обычно в центре внимания морфолога стоят: 1) структура организмов на разных уровнях их организации; 2) становление организмов в индивидуальном развитии (морфогенез); 3) взаимосвязь преобразования структуры в онто- и филогенезе; 4) функциональная интерпретация структур. Подобное понимание предмета морфологии несколько шире традиционного, так как клеточный и субклеточный уровни организации обычно включают в предмет цитологии и молекулярной биологии. Таким образом, задача морфологии как науки - отражение картины структурной эволюции на всех уровнях и этапах развития органического мира. Эта задача, очевидно, должна решаться как вширь, учитывая все таксономическое многообразие форм, так и вглубь, охватывая разные уровни организации и все этапы их развития. Исходя из этого, морфология должна занимать ключевое положение в системе биологических знаний, обеспечивая фактологическую и теоретическую базу связанных с нею дисциплин.
Эволюционная морфология современных форм в широком понимании [187, 794, 998] ставит своей главной целью изучение морфологических закономерностей эволюции и объяснения причин и механизмов формообразовательных процессов в эволюции онтогенеза и в филогенезе, опираясь на представление о целостности организма в индивидуальном и историческом развитии [2435]. Эволюционируют не признаки, а организмы в целом [1184]. Палеонтологическая морфология также имеет дело не только со статичной структурой ископаемых организмов, но и с ее становлением в онто- и филогенезе.
В то же время специфика ископаемого материала - отражение в нем длительных исторических периодов развития таксонов и фрагментарность сведений о целостности организма и его развитии - обусловливает особую научно-исследовательскую стратегию. Суть этой стратегии определяется необходимостью описать, связать воедино и объяснить исторически сложившееся разнообразие современных и вымерших групп организмов, располагая сохранившимися их остатками, чаще всего в виде твердых тканей. Естественно поэтому, что палеонтолог, в отличие от неонтолога, вынужден уделять гораздо большее внимание отдельным структурам и анализу их признаков. Успех здесь во многом зависит от того, как будет трактоваться эволюция этих структур и признаков, подразумевая, естественно, что они принадлежат целостным организмам. Таким образом, стратегия палеонтологических исследований неразрывно связана с их методологией, которая определяется как общими установками, так и конкретными материалами.
Общие установки, проблемы и направления морфологических исследований в палеонтологии и неонтологии в целом совпадают. На характер палеонтологических работ влияют в первую очередь принадлежность материала к разным группам организмов (прокариотам, растениям, беспозвоночным и позвоночным животным), его сохранность и цели, которые ставит перед собой исследователь. В разных областях палеонтологической морфологии сложились традиционные пути исследований, отличающиеся методами, подходами и принципами. В палеонтологии и неонтологии позвоночных, например, издавна (со времени Ж. Кювье и особенно после работ В. О. Ковалевского) широко применяют функционально-морфологический анализ. Онто- и филогенетические исследования, использующие для прослеживания структурных преобразований метод тройного параллелизма, выполняют как на позвоночных, так и на многих беспозвоночных (аммоноидеях, кораллах, насекомых и др.), однако они редки в морфологии вымерших растений. По данным применения метода тройного параллелизма (либо двойного - в зависимости от материала) строят эволюционные ряды животных организмов. В морфологии некоторых беспозвоночных (например, радиолярий, кораллов, иглокожих) и водорослей (диатомовых, эбридиевых, кокколитофорид и др.) в центре внимания находится симметрия форм. Широко распространено эколого-морфологическое изучение разных групп современных и ископаемых животных (в частности, в энтомологии).
В палеоботанической морфологии, как и в палеоморфологии животных, исследования чаще всего направлены на установление единичных исходных форм, из которых выводится многообразие всех остальных. При этом функциональный смысл разновидностей формы оставляется обычно в стороне. Соответственно палеоботаническая морфология взяла на вооружение идеи теломной теории, выводя разнообразие органов высших растений из системы псилофитовых теломов. Хотя преобразование форм и объясняют как результат адаптивной радиации, но чаще всего дело не идет дальше общих фраз.
Детальные морфологические исследования структур широко применяют в палеонтологии для целей формальной классификации дисперсных ископаемых остатков недостаточно ясной таксономической принадлежности (миоспор и других палиноморф, конодонтов, многих нанофоссилий и т. д.).
Различия в исследованиях рецентных и ископаемых групп организмов в значительной мере определяются характером сохранности палеонтологического материала (исключительно костные ткани у позвоночных животных, только раковины и следы ползания у моллюсков, преимущественно изолированные органы и отпечатки у растений и т. д.) и общей неполнотой геологической летописи. Безусловно, дефекты материала ограничивают (иногда весьма существенно) возможности морфологических работ. Они в известной мере определяют также их стратегию, принципы подхода к материалу, а иногда служат оправданием теоретической позиции. Так, еще в доэволюционной морфологии сложилось представление о стабильности главных структурных типов животных. Оно поддерживается и сейчас, например стокгольмской школой морфологов [1795], признающей существование 12 основных стволов позвоночных с момента их адаптивной радиации в досилурийское время. Соответственно при реконструкции таких типов используют единичных представителей групп (модельные формы) без учета хронологической последовательности таксонов, а особенности рецентных форм переносят в деталях на предполагаемых предков.
Подобные нарушения геохронологического принципа привели к ошибочным филогенетическим и эволюционно-морфологическим выводам в отношении ряда групп древних бесчелюстных и рыб [752, 764, 2433].
В то же время у палеонтологического материала есть и преимущество перед неонтологическим, поскольку он дает возможность реконструировать хронологию морфологических преобразований в эволюции групп и документировать эти преобразования на фактическом материале. Обращение к палеонтологии позволяет восстановить историческое становление структур и проконтролировать правильность выводов, гипотез и семофилогенетических (касающихся истории органов) схем, созданных на рецентном материале. В эффективности этого контроля мы легко убеждаемся. Достаточно напомнить те существенные изменения, которые внесены палеонтологами в филогенетические и таксономические построения, касающиеся многих групп животных и растений. Именно благодаря палеонтологии доказано единство рыб и рыбообразных, челюстноротые выведены из гетеростраков, тетраподы из остеолепидных кистеперых, осетры причислены к лучеперым, черепахи выделены в особую группу парарептилий, птицы объединены с динозаврами, мозоленогие обособлены в системе копытных, реконструированы ранние этапы эволюции членистоногих, гинкговые растения выделены из пинопсид и отнесены к одному классу Ginkgoopsida вместе с каллистофитовыми и пельтаспермовыми и т. д.
Каждая из подобных перестроек основана на эволюционно-морфологическом материале и, наоборот, влечет за собой важные следствия для эволюционной морфологии. Специфика палеонтологического материала приводит исследователей к необходимости следования принципу актуализма, с одной стороны, и принципу историзма, с другой.
При оценке общего смысла и результатов морфологических исследований в палеонтологии следует также четко разграничивать морфологию как самостоятельную область знаний (исследующую определенный - морфологический аспект органического разнообразия и морфологические закономерности эволюции) и морфологические методы, которые широко используются другими дисциплинами для решения собственных задач.
Почти исключительно на морфологических данных строится филогения и систематика ископаемых организмов. Данные морфологии служат косвенными свидетельствами физиологических процессов, основой палеоэкологических реконструкций, инструментом биостратиграфической корреляции отложений и т. д. Открываются большие возможности внедрения палеоморфологических исследований и в разные разделы неонтологии - в сравнительную анатомию, эмбриологию, экологию, физиологию, этологию, биогеографию и др.
В то же время укрепление связей морфологических исследований рецентных и ископаемых форм благотворно влияет на дальнейшее развитие палеонтологической морфологии. Складывается обратная связь тех и других исследований, продуктивность которой очевидна. Особенно наглядно это видно в области эволюционной морфологии низших позвоночных. В разработке проблемы происхождения тетрапод соединение эмбриологических, сравнительно-анатомических и эколого-физиологических исследований с палеонтологическими подчас приводит к самым неожиданным результатам, заставляющим в корне пересмотреть сложившиеся представления, например в отношении формирования звукопроводящей системы в эволюции позвоночных [964].
Конкретные проблемы эволюционной палеоморфологии в целом совпадают с проблемами морфологии рецентных организмов: установление взаимоотношений формы, функции и адаптивной роли органов; выяснение связей частей и целого; соотношение онто- и филогенеза; изучение целостности организма в индивидуальном и историческом развитии и эволюции его коррелятивных систем; решение проблем гомологизации, выяснение модусов морфологической эволюции и т. д. Однако в силу специфики задач, стоящих перед палеонтологией, пути разработки этих проблем и их относительное значение имеют свои особенности. Прежде чем обсуждать современные проблемы морфологических исследований в палеонтологии целесообразно кратко рассмотреть некоторые общие тенденции развития морфологии в целом.
Тенденции развития морфологии
Хотя морфология рано сформировалась в самостоятельную область знаний, однако со времен античности и до XIX в. морфологические исследования не были четко отделены от физиологических (функциональных); одновременно познавалось как строение главных органов, так и их функционирование. Так, открытие системы кровообращения означало и изучение морфологии кровеносной системы и вместе с тем установление непрерывности кровотока путем проведения аналогии между сердцем и насосом.
Лишь в конце XVIII - начале XIX в. углубление и расширение исследований, а также трудности в непосредственной функциональной интерпретации всех аспектов морфологического разнообразия организмов привели к существенному обособлению морфологических исследований от функциональных (физиологических). Тогда же стали все яснее формулироваться и собственно морфологические проблемы, в том числе - единства морфологического типа разных групп организмов, связи части и целого, метамерии и целостности развития. Появились работы, в которых строились ряды форм, связывающие планы строения разных таксонов. Внимание исследователей особенно привлекало единство в строении внешне весьма различных организмов.
На этой ступени своей истории морфология обрела тесные связи с систематикой. Именно поэтому некоторые фундаментальные теоретические концепции биологии оказываются в равной степени и морфологическими, и таксономическими. Например, учение о планах строения утверждало существование инвариантности в строении организмов и в то же время подчеркивало разнообразие планов строения и их принадлежность определенным таксономическим группам.
Еще в прошлом веке наметились самостоятельные исследовательские программы в морфологии, физиологии и таксономии, что сопровождалось теоретическим осмыслением этих дисциплин. Оформились основные морфологические идеи, иногда совершенна противоположные. Одновременно возникли и теоретические коллизии, касающиеся проблем соотношения форм и функции, аналогии и гомологии, дискретности типов организации или наличия переходов между ними и другие, отчасти удержавшиеся до наших дней.
Хотя эволюционная идея проникла в морфологию задолго до Ч. Дарвина, однако только после торжества дарвинизма она стала играть роль главного теоретического стержня. Эволюционная теория упрочила и придала новый смысл взаимосвязям морфологии с таксономией и физиологией, обеспечив также связи с экологией (что произошло много позже). Тем не менее в самой морфологии проявились и определенные тенденции к теоретическому сепаратизму, свойственные так называемой "идеалистической морфологии". В рамках этого направления, расцветшего в 20-е годы нашего века, почти не уделялось внимания трактовке биологической роли морфологического разнообразия организмов, смысл исследований усматривался исключительно в установлении структурной упорядоченности форм, выведении чисто морфологических инвариант.
"Идеалистическая морфология" противостояла эволюционной морфологии, которую на первых этапах интересовали преимущественно филогенетические проблемы: не столько инвариантность форм и конструктивные закономерности их преобразования, сколько их становление в филогенезе. Противником "идеалистической морфологии" стал морфофункциональный анализ, который постепенно переходил от чисто физиологических аспектов изучения (изучение функциональной взаимосвязи органов) к экологическим (выяснение зависимостей между строением органов и их функционированием во внешней среде, оценка биологической роли структур).
История становления и развития этих теоретических альтернатив подробно изложена в ряде работ [87, 192, 468, 469, 691, 1477, 1936, 2202]. По мере развертывания дискуссий становится все более очевидным, что разные теоретические установки в морфологии не столько противоречат, сколько дополняют друг друга. Синтетическая точка зрения на цели и методы морфологических исследований в свете современных задач биологии дана В. Н. Беклемишевым: "Мы объясняем строение органа, когда указываем условия, которые вызывают его появление в онтогенезе особей данного вида и которые поддерживают его существование в непрерывном потоке обмена веществ (физиология формы). Мы объясняем строение органа, когда указываем, каким образом он приспособлен к своим отправлениям, каково его значение в жизни целого организма; аналогичным образом мы объясняем строение организма, указывая, каким образом он приспособлен к условиям своего существования (экологическая морфология). Мы объясняем строение организма или органа, а также их приспособленность к окружающей среде, когда устанавливаем путь и движущие силы их становления в истории вида. Но кроме физиологического, экологического и исторического объяснения необходимо и возможно также и сравнительно-анатомическое объяснение. Так, понять голову насекомого - это значит понять, как она построена из отдельных сегментов, ее составляющих, что позволит нам точно сравнить ее с головами других членистоногих. Итак, сравнительно-анатомическое или, шире, конструктивно-морфологическое объяснение строения какого-либо органа или организма сводится к более точному и строгому сравнению этих объектов со всеми остальными организмами и их органами, к нахождению естественного места этого объекта в общей системе многообразия организмов.
Из сказанного ясно, что каждый организм, орган, ткань могут и должны быть изучены по меньшей мере с четырех различных точек зрения: конструктивно-морфологической, физиологической, экологической и исторической. Каждая из этих точек зрения вполне законна, необходима и незаменима" [61, с. 7-8]. В этой обширной цитате изложена общая теоретическая установка.
Для определения места каждого организма и его компонентов в общем многообразии органического мира необходима "свертка" морфологического многообразия в систему определенных закономерностей. При рассмотрении этих закономерностей увеличение разнообразия форм за счет дивергенции часто противопоставлялось его сохранению или сокращению за счет параллелизмов, конвергенции и морфофункциональных перестроек. Такое противопоставление издавна служит источником теоретических противоречий между эволюционистами. В селекционизме основой эволюционного процесса считается морфологическая (и иная) дивергенция, тогда как конвергенция и параллелизм рассматриваются как явления второстепенные. Наоборот, эволюционные учения номогенетического толка концентрируют внимание на конвергенции и параллелизме, видя в них проявление основных закономерностей эволюции.
Однако здесь вполне возможна синтетическая точка зрения. Существование конвергенции и параллелизмов и их широкое участие в эволюции - твердо установленное эмпирическое обобщение. В то же время без дивергенции они не могут проявляться. Сами понятия конвергенции и параллелизма хотя бы неявно подразумевают ссылку на дивергенцию, поскольку под этими явлениями понимается возникновение или сохранение тех или иных сходных черт на фоне возрастающего разнообразия. Кроме того, из общесистемных соображений следует, что чистая дивергенция (без вторичного возникновения сходств) в принципе не может быть беспредельной. В дивергенции должен проявляться определенный параллелизм хотя бы потому, что число наиболее общих способов преобразования любых систем конечно [647, 1082, 1950]. К этому добавим, что у организмов способы морфологических преобразований ограничены канализованностью морфогенетических процессов и системой морфофункциональных запретов [189, 326, 533, 1685, 1950, 2445].
Необходимость "свертки" морфологического разнообразия диктует следующие компоненты морфологических исследований ископаемых и современных организмов.
1. Изучение конструкций организмов, в том числе их онто- и филогенетического становления, с целью выявления черт, повторяющихся от таксона к таксону, что позволит свести наблюдающееся морфологическое разнообразие организмов к немногим исходным формам и проследить их соотношение в эволюции.
2. Эколого-физиологические интерпретации выявленных морфологических свойств. Этот морфофункциональный анализ также может быть совмещен с онто- и филогенетическими исследованиями.
3. Выведение некоторых наиболее общих способов (модусов) морфологического преобразования и прослеживание роли этих модусов в разных группах организмов на разных этапах их истории в разных экосистемных условиях.
Полнота реализации такой программы зависит от характера материала и его изученности, в частности, от группы организмов (растения, разные беспозвоночные и позвоночные животные), сохранности объектов (современные или ископаемые формы), исследуемого уровня структурной организации (тканевого, уровня органа или систем органов).
При выдвижении основных позиций мы опирались на исторически сложившиеся исследовательские программы, на давние традиции морфологических работ и на привычный круг морфологических понятий. Вполне возможны и иные пути исследования морфологического многообразия, но при этом важно не противопоставлять конструктивно-морфологические и морфофункциональные исследования, исходя из различения понятий процесса и структуры.
Любая биологическая структура может быть представлена в статике лишь в определенных временных рамках (в принятой метрике времени). Фактически все органические структуры - лишь стадии определенных материально-энергетических и информационных процессов. В некоторых исследованиях само противопоставление понятий "процесс" и "структура" почти неосмысленно (например, на молекулярном уровне). С другой стороны, в палеонтологии мы лишены возможности непосредственно изучать какие-либо физиологические процессы. Органические формы предстают перед нами набором застывших стадий, процессы останавливает не исследователь, а сама природа. С этим приходится считаться, и это создает дополнительные проблемы при формулировке задач и принципов палеоморфологического исследования. Итак, мы снова возвращаемся к специфике палеонтологического материала, на которой остановимся более подробно, прежде чем рассматривать конкретные морфологические исследования в палеонтологии.
Основные особенности палеонтологического материала
Общее свойство палеонтологического материала - его неполнота. Аспекты этой неполноты многообразны. Нам недоступны для изучения многие мягкие части организмов, не сохранившиеся в ископаемом состоянии. Лишь у немногих древних форм удается проследить становление органов в онтогенезе одной особи. В филогенетических рядах имеется немало существенных пробелов. Из геологической летописи выпадают многие таксоны. Невозможно непосредственное наблюдение функционирования организмов. Однако соответствующие недочеты ископаемого материала не следует преувеличивать. История палеонтологии убеждает нас в том, что ее данные позволяют надежно реконструировать сколь угодно сложные и никогда не сохраняющиеся структуры организмов, оценить их биологическую роль, разнообразные физиологические и онтогенетические процессы, проследить и дать общую картину смены форм и их экологических связей.
Примерами широких возможностей палеонтологии могут служить реконструкции нервной, кровеносной, мускульной систем и электрических органов у бесчелюстных [752, 2329], панцирных [766] и кистеперых рыб [186, 1340, 1797], мускулатуры у индрикотерия [107], мягких губ у териодонтов [1033], эволюции летательного аппарата у насекомых [868], репродуктивность цикла многих ископаемых растений [2370] и т. д. (см. гл. 1.4).
В то же время современные организмы также подчас недоступны для детального изучения. На стол исследователя они нередко поступают законсервированными, фиксация материала необратимо разрушает многие структуры и лишает морфолога возможности непосредственно наблюдать функционирование органов и их становление. Об онтогенетических процессах в этих случаях судят путем сопоставления особей разных стадий развития, а о функциональном смысле (биологической роли) органов - по аналогии и при сопоставлении особенностей организма со свойствами среды обитания. В подобных случаях работа с рецентным и ископаемым материалом в целом сходна. Разница лишь в более полной сохранности искусственно фиксированного рецентного материала по сравнению с естественно фиксированным ископаемым. И поскольку палеонтолог больше ограничен в выборе тех признаков, которые он может подвергнуть анализу, его исследование отличается и большей трудоемкостью.
Неполнота ископаемых остатков накладывает отпечаток на все этапы морфологического исследования и нередко заставляет палеонтолога вводить не только специальные технические приемы работы, но и особые "палеоморфологические" понятия.
Полнота рецентного материала вынуждает палеонтолога постоянно обращаться в качестве основного исследовательского приема к принципу актуализма, учитывая, разумеется, свойственные ему ограничения. На принципе актуализма в конечном счете основаны все морфологические и таксономические решения. Без обращения к актуализму мы не смогли бы даже считать изучаемые объекты ископаемыми организмами. Как бы ни были специфичны ископаемые остатки, мы относим их к растениям или животным, а затем к тем или иным их группам или частям организмов только благодаря тому, что все эти таксономические и морфологические понятия установлены для современных организмов.
Главное преимущество палеонтологического материала по сравнению с рецентным - его приуроченность к разным временным интервалам и соответственно возможность изучения исторического становления и развития организмов. Принцип историзма издавна является ведущим в палеонтологии и дополнительным к актуализму. Если актуализм открывает возможности разобраться в прошлом с помощью настоящего, то историзм, наоборот, представляет настоящее в исторической перспективе, указывает на те исторические предпосылки, которые обусловили нынешнее состояние изучаемых объектов. Тезису актуализма "настоящее - ключ к познанию прошлого" можно противопоставить тезис историзма "прошлое - ключ к познанию настоящего".
Реконструкция истории часто возможна путем сравнения ныне живущих форм, построения их эволюционных рядов. Современной морфологии такую возможность дают филогенетика, систематика и биогеография ныне живущих организмов, их физиология и экология. На этой основе покоится эволюционная морфология многих организмов, отсутствующих или крайне редких в геологической летописи (инфузорий, турбеллярий, голотурий, лишайников и др.).
Для исследования истории групп и их происхождения морфологи широко используют собственно морфологические, в том числе эмбриологические данные. Само понятие эволюционной морфологии - науки, исследующей морфологические закономерности эволюционного процесса и историческое становление форм, была введено в отечественную науку А. Н. Северцовым на основе сравнительно-морфологических исследований рецентных низших позвоночных. Соединение сравнительно-морфологических (включая палеонтологические) данных с физиологическими и экологическими позволяет получить ценные сведения о направлениях, путях и особенностях эволюции многих групп организмов и структур. Интересные результаты в изучении исторического становления форм дает в последние годы функциональная морфология позвоночных [446, 500, 1346, 1348, 1529, 1530, 1618, 1685], использующая комплекс функционально-морфологических методов для изучения функций и адаптации.
Однако опыт работы с группами, хорошо представленными в геологической летописи, убеждает в том, что по одним лишь современным формам нельзя составить объективное представление об историческом изменении органов и организмов. Среди выдающихся морфологов-эволюционистов это особенно хорошо понимал И. И. Шмальгаузен [1190], который, разрабатывая проблему происхождения наземных позвоночных на современных амфибиях, постоянно обращался к сведениям об их ископаемых предшественниках.
Случаи, когда ознакомление с палеонтологическим материалом только подтверждало существующие эволюционно-морфологические представления, основанные на рецентном материале, сравнительно редки. Гораздо чаще палеонтологический материал значительно расширяет наши представления об эволюции групп и показывает, что преобразование морфологических типов шло путем, который было невозможно реконструировать без знания ископаемых форм.
Нередко морфологи намеренно игнорируют палеонтологический материал, считая его неполным и непредставительным. Такую позицию занимал, например, известный морфолог растений К. Гебель, который в своих исследованиях практически обходился без обращения к палеоботаническим данным или, во всяком случае, не анализировал их сколько-нибудь тщательно. Отдавал предпочтение рецентному материалу при исследованиях морфогенетических закономерностей эволюции позвоночных и А. Н. Северцов, что привело к ошибочным результатам в реконструкции филогении ряда групп [1032, 1033].
Невнимание к палеонтологической летописи сказывается на обсуждении крупных морфологических проблем, являясь одной из причин тенденциозности и в выборе проводимых исследований, и в оценке получаемых выводов. Так, одним из наиболее дискутируемых вопросов в ботанике издавна является гомология частей цветка и само его происхождение. Поскольку специальные исследования репродуктивных органов древнейших покрытосеменных не проводилось, это породило миф, будто нужные палеоботанические данные вообще не могут быть получены и даже излишни. Для сравнения с современными цветками привлекались лишь некоторые ископаемые голосеменные, выбор которых тоже был случайным или определялся уже имеющейся морфологической концепцией.
Говоря об обращении морфологов к палеонтологическим данным, мы имеем в виду не просто учет вымерших таксонов, но также и их геохронологические соотношения. Случалось, что как морфологи, так и палеонтологи учитывали в своих построениях ископаемые формы, но пренебрегали сведениями об их относительном возрасте (стратиграфическом положении). Такие примеры в отношении древних рыбообразных приводит Д. В. Обручев [765], критикуя работы стокгольмской школы палеонтологов. В частности, этой школой было принято, что удлинение панциря цефаласпид, приведшее к утрате грудных плавников, есть специализация, а примитивное состояние - короткий панцирь с развитыми грудными плавниками. Соответственно формы типа Tremataspis и Thyestes оказываются производными от форм типа Kiaeraspls и Cephalaspis, хотя первые два рода венлокско-лудловские, а вторые - раннедевонские. Последующие исследования показали, что возникновение плавников из боковых складок происходило независимо у бесчелюстных и челюстноротых. Морфологические преобразования форм были совмещены со стратиграфической последовательностью таксонов.
В последние годы идея о вторичности безплавниковых трематаспид снова возродилась, хотя и на другой основе - на филогенетической систематике [1789], которая также игнорирует геохронологическую последовательность форм. В настоящее время на Западе среди морфологов и систематиков особую популярность приобрел "трансформированный кладизм" [1423], соединивший филогенетический метод В. Хеннига [1727] с философскими принципами К. Поппера [835]. Такое соединение привело некоторых специалистов к выводу о вторичной роли палеонтологии в решении вопросов филогенетики и эволюционной морфологии [2071]. Палеонтологической морфологии неокладисты отводят небольшую роль корректирования созданных неонтологами филогенетических построений и теорий. Основные таксономические группы, выделенные на рецентном материале, переносятся на ископаемые формы, которые игнорируются, пока не выяснена их филогенетическая судьба. Так, огромная группа палеонисков, стоящая в основании ствола костистых рыб, расценивается "как корзина для мусора неспециализированных форм" [2211], хотя ископаемые представители этой группы достаточно хорошо охарактеризованы.
Обобщая опыт палеоморфологических исследований, можно утверждать, что, несмотря на недостатки палеонтологического материала, он служит наиболее надежной основой выяснения генезиса структур, реконструкции филогенеза групп, поиска их предковых форм и установления общих макроэволюционных закономерностей. С начала века именно палеонтология стала продуцировать основные морфологические концепции, касающиеся позвоночных, головоногих, иглокожих, фораминифер, большинства классов высших растений и многих других групп организмов. Одновременно усилилась и зависимость палеонтологии от морфологии рецентных организмов, поскольку при реконструкции ископаемых форм приходится обращаться к все более широкому кругу морфологических признаков и прежде всего тех, реконструкция которых без знания современных форм просто невозможна.
Кроме того, рецентный материал дает возможность проследить диапазон вариаций тех или иных структур и выявить степень их эволюционной пластичности, что может служить критерием для оценки их филогенетической значимости. Так, установленная на рецентных амфибиях вариабельность в строении звукопроводящего аппарата Anura и Urodela, обеспечиваемая наличием двойного канала звукопроведения оперкулярной и тимпональной систем, разной степенью их редукции и компенсаторностью, заставляет пересмотреть возможности широкого использования данной структуры для филогенетических целей в палеогерпетологии [964].
В настоящее время наиболее продуктивны те морфологические исследования, в которых глубоко интегрируются сведения об ископаемых и рецентных формах, когда само исследование современного материала в значительной мере определяется запросами палеонтологии и, наоборот, у ископаемых форм ведется поиск тех признаков, значимость которых ясно показана изучением их ныне живущих потомков.
В целом можно сказать, что вопрос о том, нуждается ли морфология в обращении к палеонтологическому материалу, решен позитивно. Обсуждать следует в основном методологическую сторону: как и когда привлекать палеонтологический материал к решению тех или иных морфологических вопросов на конкретных группах. Одним из таких путей могут служить конструктивно-морфологические исследования ископаемого материала.
Конструктивно-морфологические исследования
Перед конструктивно-морфологическими исследованиями - их не следует смешивать с "конструктивизмом" [1685] - стоят две основные задачи. Во-первых, это сравнительное изучение многообразия строения организмов и соотношений их частей путем выделения в целом организме отдельных гомологизированных компонентов (меронов) и установления их свойств, что и составляет предмет тектологии [650]. Соответственно тому, о каких уровнях организации идет речь - органах, тканях, клетках, - можно различать в составе тектологии отдельно органологический, гистологический и цитологический разделы. Более низкие уровни организации обычно уже не включают в морфологию и относят к области изучения молекулярной биологии. Вторая задача связана с выяснением того, как изолированные части организмов (мероны) соединяются, взаимодействуют (коррелируют) и как из них складывается целостный организм (архетип). Эту часть конструктивно-морфологических исследований называют архитектоникой. Тектология и архитектоника расцениваются отнюдь не как самостоятельные морфологические дисциплины, а скорее как аспекты единого морфологического - мерономического [650, 651] - исследования, и разграничение их целесообразно лишь при формулировках морфологических понятий.
Палеозоологические морфологические исследования носят преимущественно органологический и архитектонический характер, тогда как гистологические исследования играют здесь все еще второстепенную роль. В палеоботанической морфологии несравненно большее значение придается (наряду с органологией) исследованию тканей и отчасти клеток (например, утолщению стенок древесины). Морфология ископаемых растений, как и современных, практически сливается с гистологией (анатомией). Некоторые разделы цитологии растений также часто входят в морфологию (например, исследование строения оболочек у спор и пыльцы). Давнее, еще со времен И. В. Гёте, противопоставление морфологии и анатомии растений сейчас сохраняется главным образом в учебных курсах и библиографических рубриках, а также в отдельных исследовательских направлениях (например, при изучении жизненных форм). Архитектонический аспект морфологии (постоянно сопутствующий тектологическому при изучении большинства животных) нередко сводится к минимуму в палеоботанике (например, при изучении дисперсных семян или пыльцы). В этом случае, когда мы не можем реконструировать общий облик организма, архитектонический анализ касается не организма в целом, а лишь частей определенного органа.
В рамках мерономического подхода организм расчленяют на разные типы меронов (разные типы органов, их систем, тканей, клеток), которые классифицируют по определенным критериям. Выделение меронов, т. е. гомологизированных частей, включает операцию гомологизации. Процедурам гомологизации посвящена огромная литература [87, 468, 469, 1279, 1346, 1790, 1944, 2141, 2289, 2431, 2492 и др.]. А. Ремане четко сформулировал известные порознь и до него три основных критерия гомологичности: критерий положения, критерий специального качества и критерий непрерывности (серии переходных форм). Три дополнительных критерия мы здесь не рассматриваем.
Критерий положения означает, что даже разные по собственным признакам части могут быть гомологичными, если они занимают одинаковое положение по отношению к другим частям. Данный критерий применим лишь тогда, когда соседние с изучаемой части уже надежно гомологизированы. Например, когда говорится, что лобные кости (frontale) тетрапод определяются положением между теменными (parietale) и носовыми (nosale), то это значит, что сами теменные и носовые кости надежно идентифицированы (по отношению к нейрокранию, теменному отверстию и ноздрям). Если такого надежно гомологизированного соседства нет, то появляется множество вариантов для гомологизации.
Так, обычно нуцеллюс голосеменных сопоставляется с мегаспорангием споровых по критерию положения (соседство с мегаспорой). Затем была выдвинута гипотеза, что мегаспоровая мембрана голосеменных гомологична тапетуму, т. е. внутренним слоям стенки спорангия. Тогда нуцеллюс можно было сопоставить с органом, вмещающим спорангий, например с интегументом наиболее примитивных голосеменных [517]. Более детальные исследования исторической последовательности семян голосеменных показали [660, 1955], что нуцеллюс гомологичен мегаспорангию целиком только в классе Ginkgoopsida, а в классах Pinopsida и Cycadopsida первичный нуцеллюс полностью слился с первичным интергументом (рис. 1). Установить эту гомологию без обращения к ископаемым голосемянным лишь по критерию положения оболочек у современных семян принципиально невозможно.
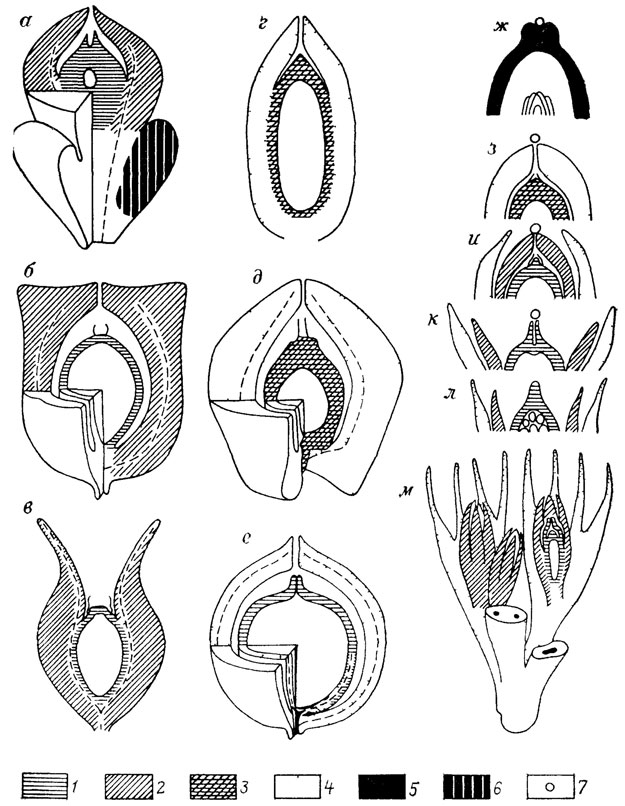
Рис. 1. Гомологизация и смена функции покровов у семян. По С. В. Мейену. а-в - класс Gmkgoopsida а - Ginkgo, б - Callcspermanon, в - Lyraspertna, г-е - класс Pinopsida, г - Ptnus, д - Mttrospermum, е - Nucellangtum, ж - захват пыльцы рыльцем; з - захват пыльцы купулой, первичный интегумент слился с нуцеллюсом, и - захват пыльцы интегументом, доли купулы несомкнутые, к - захват пыльцы сальпинксом, доли интегумента несомкнутые, л - споровый предок голосеменных; м - наиболее примитивные Cycadopsida (семеносная купула типа Hydrasperma). 1 - нуцеллюс, 2 - интегумент, 3 - слившиеся интегумент и нуцеллюс, 4 - купула и производный от нее внешний (вторичный) интегумент, 5 - плодолистик, 6 - воротничок (ложная купула); 7 - зерно пыльцы или предпыльцы
В палеонтологии ценность критерия положения нередко дискредитируется тем, что приходится сравнивать дисперсные части, прижизненное положение которых неизвестно. Именно поэтому нельзя было ответить на вопрос о том, что такое конодонты. Лишь когда был найден отпечаток животного-конодонтоносителя с прижизненным положением конодонтов в его головной части, на этот вопрос был получен частичный ответ, частичный из-за неполной сохранности найденного экземпляра [1379].
Многие органы, принадлежащие разным морфологическим категориям и занимающие разное положение в целом организме (точнее, архетипе), могут быть весьма сходными по внешним признакам. Например, внешне одинаковая чешуя и плавники могут занимать совершенно разное положение на теле рыб. Чешуевидные листья одинакового облика могут быть почечными чешуями и элементами обвертки фруктификации. Не зная их прижизненного положения, нельзя указать тот мерон, к которому они принадлежат. Поэтому палеонтологи вынуждены вводить самостоятельные морфологические понятия и термины (например, конодонты, аптихи, палиноморфы, миоспоры, зубные спирали, фулькры). Подчеркнем еще раз, что дело не в том, что соответствующим органам нет гомологов у современных организмов. Таковые, может быть, и имеются, но мы не можем их указать из-за разрозненности частей у ископаемых форм.
В некоторых случаях дело с выяснением морфологической принадлежности разрозненных частей обстоит сложно даже при наличии, казалось бы, совершенно достоверных гомологов у современных организмов. Например, часть ископаемых ринхолитов имеет такое же строение, как и обызвествленный кончик верхней челюсти у современного наутилуса. В их принадлежности к наутилоидеям никаких сомнений нет. Большая часть групп ринхолитов отличается достаточно, а иногда и весьма значительно от "типичных форм". Относительно них всегда были разные суждения; их приписывали также вымершим наутилоидеям, аммоноидеям, внутреннераковинным головоногим и даже неизвестным группам головоногих (очевидно, безраковинным, а потому и не сохраняющимся). Смущало отсутствие находок ринхолитов вместе с аммонитами и отсутствие ринхолитов в палеозое, хотя аммониты там были в изобилии. Известны были и остатки внутреннераковинных головоногих с крючочками рук, но без ринхолитов. Только находки в последние годы аммонитов с ринхолитами убедительно показали, что все же часть ринхолитов принадлежала этой большой группе головоногих.
Показательны затруднения относительно гомологизации рук, щупалец и воронки головоногих моллюсков с ногой других классов моллюсков. Далеко не все ясно с вопросом о назначении так называемых аптихов аммоноидей. Первоначально была принята точка зрения, что аптихи являются крышечкой, образующейся у аммоноидей подобно тому, как у современного наутилуса возникает черный слой над устьем раковины на вентральной стороне предыдущего оборота. В настоящее время аптихи рассматривают в качестве элементов челюстного аппарата. Такое решение проблемы ставит ряд новых вопросов и прежде всего вопрос о возможности сопоставления аптихов с крышечкой далеких предков головоногих и путях ее перемещения с ноги в глотку или о аптихах как новообразовании у головоногих.
Когда говорят о критерии положения, может подразумеваться не только место данной части в организме (мерона в архетипе), но и место определенной фазы в жизненном цикле. Недавно появились данные, что остатки, описанные под названием Sciadophyton и Zosterophyllum, не разные роды, а гаметофит и спорофит одного и того же растения. То же касается и рода Strunius, который оказался ювенильной формой рода Onychodus среди кистеперых рыб. Можно говорить о гомологии дентиновой чешуи с шагренью, малыми челюстными и клыкоподобными зубами у низших позвоночных.
Серьезные ограничения на применимость критерия положения накладывает не только несовершенство ископаемого материала, но и сама неустойчивость положения мерона в архетипе. Речь идет о явлениях, обозначаемых в литературе как субституция [87], меторизис, гомеозис, гетеротопия и др. Вероятно, роль этих явлений в эволюции формы весьма значительна, хотя проявляется нечасто. В некоторых случаях именно смена положения органа обусловливает возникновение существенно нового мерона и соответственно нового таксона высокого ранга. Есть основания полагать, что у растений гомеозис, переходящий в гетеротопию, - один из основных путей возникновения принципиально новых типов органов. Например, семеносные органы многих голосеменных произошли, очевидно, путем сальтационной пересадки семян на вегетативные листья. Такие органы, возникшие путем рекомбинации онтогенетических программ, занимают в системе морфологических понятий то же место, что и гибриды в системе таксономических понятий [2197]. В результате листоподобный орган с сидящими на нем семенами или полиспермами уже нельзя квалифицировать как лист без коренных изменений в самом понятии листа.
В палеозоологии известны многочисленные случаи, когда органы могут менять свое положение в зависимости от состояния развития и тогда возникают сомнения по поводу их гомологии. Так, ушной отросток палатоквадратум у тетрапод считался негомологичным ушному (а точнее, паратемпоральному) отростку кистеперых рыб в связи с их разным местом прикрепления к черепу. Однако известные на сегодня варианты в состоянии ушного отростка кистеперых рыб и в его прикреплениях [186] вынуждают более широко подходить к определению такого отростка - как любого выступа нёбно-квадратной кости, ограничивающего сзади тригаминальную вырезку (рис. 2). Вместе с тем в состав мозговой коробки рыб входят разные компоненты их висцерального аппарата, что создает мозаику гомологичных и негомологичных участков черепа, обеспечивая в то же время разнообразие морфологических и таксономических их форм на уровне крупных систематических единиц (эласмобранхии, круглоротые, кистеперые, дипнои и т. д.).
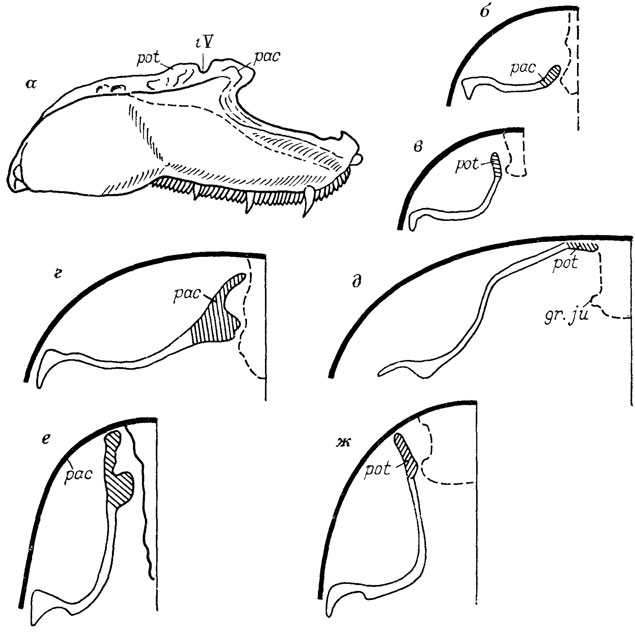
Рис. 2. Нёбно-квадратный комплекс и схемы его прикрепления к черепу (поперечное сечение) с помощью ушного и паратемпорального отростков у разных кистеперых. По Э. И. Воробьевой. а - общий вид комплекса (Eusthenopteron foordi); б, в - Thursius estonicus; г-д - Gyroptychius pauli, е, ж - Eusthenopteron foordi; gr-ju - борозда боковой вены головы, pac - паратемпоральный отросток; pot - ушной отросток, iV - тригеминальная вырезка
Критерий специального качества - это, в сущности, использование всех тех собственных признаков частей, по которым части объединяются в мероны. Усовершенствование исследовательской техники резко расширило круг используемых признаков, включив в него тонкую структуру (микроструктуру и ультраструктуру), биохимические особенности, детальное прослеживание онтогенеза частей. Сказанное касается и палеонтологического материала. Изучение микроструктуры становится важным компонентом морфологических исследований в палеонтологии. В палеоботанике соответствующие исследования широко проводились еще в прошлом веке и сейчас обязательны для любой квалифицированной морфологической работы. Без изучения микроструктуры невозможно правильно интерпретировать очень многие части растений. Требуется знание их васкуляризации, эпидермальных и других микроструктурных признаков. В палеозоологии изучение микроструктур для целей гомологизации органов предпринимают реже. Как показывают исследования зубов и покровных костей у древних рыбообразных и рыб, их микроструктура может иметь весьма сходное строение в заведомо далеких филумах и поэтому должна использоваться в филогенетике в сочетании с комплексом других признаков [752, 2432].
Использование критериев положения и специального качества порождает обычные проблемы демаркации: какое положение считать разным или одинаковым, какие качества следует считать сопоставимыми или несопоставимыми. Между достаточно отличающимися органами либо системами органов, относимыми к разным меронам, могут наблюдаться постепенные переходы.
Сама сложность переходов послужила формулировке третьего основного критерия гомологии, названного критерием непрерывности (серии переходных форм). Действительно, можно привести много примеров, когда благодаря переходным формам гомологизируются разительно отличающиеся органы: гиомандибула (подвесок) и слуховая косточка; плавательный пузырь и легкое; полиплакоидная чешуя и зубы: субкраниальный мускул и полярные хрящи [1340]. Небольшой воротничок, окружающий семя гинкго, напоминает купулу лагеностомных и совершенно не похож на семеносные вайи каллистофитовых. Тем не менее переходные формы, наблюдаемые среди каллистофитовых и пельтаспермовых, ясно показывают, что воротничок гинкго должен быть гомологизирован с семеносной вайей, а не с купулой (см. рис. 1).
Построение серий переходных форм для гомологизации частей применялось еще на заре становления морфологии. Именно построение таких серий позволило выявить еще в XVIII в. общность плана строения (архетипа) в разных группах организмов и в то же время сделать заключение о несводимости архетипов, свойственных разным таксонам высшего ранга. Гиатусы между главными архетипами были в дальнейшем заполнены путем обращения, с одной стороны, к эмбриологии (здесь главную роль сыграла теория зародышевых листков при всех недостатках ее первоначального изложения) а с другой стороны, - к палеонтологии. Установление вымерших таксонов, заполнявших (хотя бы частично) гиатусы между современными таксонами, быстро привело к тому, что палеонтологический метод стал одним из основных методов морфологического исследования. Однако одно дело мысленно реконструировать переходные формы и другое - находить такие формы среди когда-то существовавших организмов. Именно после таких поисков и сформировался окончательно метод тройного параллелизма в морфологии и филогенетике, сохраняющий ведущее значение при поиске переходных форм и в наши дни. Главное изменение, которое претерпел этот метод с середины прошлого века, это замена его эмбриологической компоненты. Ведущее место рекапитуляции (биогенетического закона Геккеля) заняли феномен зародышевого сходства (закон Бэра) и модусы филэмбриогенеза.
Метод тройного параллелизма, конкретизируя критерий переходных форм, указывает, что одного лишь наличия переходов между дефинитивными стадиями разных таксонов может быть не всегда достаточно для гомологизации. Надежную гомологизацию мы получаем преимущественно лишь тогда, когда переходы демонстрируются, кроме того, в эмбриологических (если они не затемнены ценогенезами) и палеонтологических рядах. Установление гомологии стало методом и компонентом филогенетики.
Связь между теорией гомологии и филогенетикой породила новые теоретические проблемы, обсуждаемые до сих пор. Первоначально понятие гомологии не увязывалось с исторической преемственностью гомологизируемых форм, а означало лишь одинаковое положение органов в их системе (меронов в архетипе). Введение историзма в морфологию привело к переводу многих морфологических понятий на язык исторических соотношений. Последовательные ряды форм стали интерпретировать как филогенетические последовательности, архетип стали трактовать как образ предковой формы, сходство прямо связывали с родством. Возникшие теоретические противоречия достаточно подробно рассмотрены в литературе [87, 468, 469, 1161, 1894].
Современная ситуация в этом отношении мало отличается от той, которая сложилась еще в прошлом веке. Правда, намечаются сближения разных точек зрения. Обращение к палеонтологическому материалу ясно показало, что практически неразличимые органы разных современных таксонов, обнаруживающие совпадение в собственных признаках и в их положении в системе других органов, тем не менее могут быть негомологичными.
Наиболее яркий пример отсутствия гомологии между органами, всегда гомологизировавшимися до ознакомления с палеонтологическими рядами, - семена голосеменных. По морфологическим и анатомическим признакам, как и по критерию положения, оболочки семян современного гинкго прекрасно сопоставимы с оболочками семян современных хвойных. Особенно близкое сходства отмечается между семенами гинкго и тиссовых, которые размещены во внешне сходных органах (воротничке в первом случае и ариллюсе во втором). Однако знакомство с ископаемыми голосеменными убеждает в том, что это сходство поверхностное (см. рис. 1). Интегумент у гинкго первичный, а у хвойных вторичный; первичный интегумент хвойных слился с нуцеллюсом, а вторичный образовался за счет купулы, которой никогда не было у предков гинкго и нет у него самого [1955]. Таким образом, сходство оболочек у семян гинкго и хвойных скорее всего относится к феномену псевдоциклического сходства; сюда же относится, например, сходство цветков и соцветий [1279, 1627, 2141, 2397].
С помощью ископаемых материалов удалось установить разное происхождение мясистых удлиненных лопастей в парных плавниках дипной, кистеперых рыб и полиптеруса, хотя в целом архетипы их сопоставимы, представляя внешне сходные модификации грудного и брюшного плавников [194]. Ставится под сомнение гомологичность хоан у тетрапод и разных групп рыб (дипной, кистеперых, лучеперых). Понять их подлинную природу помогают переходы у ископаемых форм. Так, находки древнейших дипной в КНР свидетельствуют в пользу происхождения хоан двоякодышащих рыб из сместившейся в ротовую полость задней наружной ноздри, как и у некоторых современных лучеперых рыб, в частности некоторых нототениевых. В то же время у древних кистеперых (поролепид) наружная задняя ноздря соединена с хоанной вырезкой [1792].
У остеолепид задняя наружная ноздря отсутствовала, но хоанная вырезка в нейрокрании имелась (рис. 3) Топографически эта вырезка соответствует хоане древнейших тетрапод - ихтиостегид [2436] Уточнение гомологии всех этих структур весьма важно и неизбежно ведет к пересмотру существующих филогенетических связей названных групп.
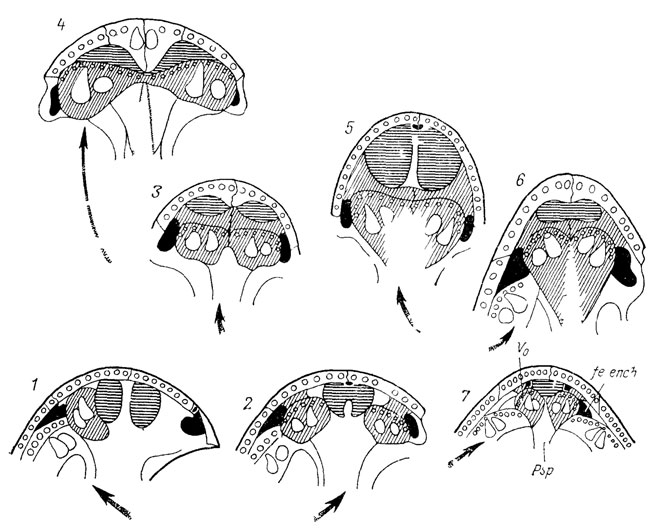
Рис. 3. Схема эволюционных преобразований и модификаций в строении нёбной поверхности черепа среди кистеперых рыб По Э. И. Воробьевой. 1 - Holoptychnda 2-7 - Osteolepidida 2 - Thurstus estotucus (примитивное состояние), 3 - Megistolepis klementzt 4 - Megahchthys nitidus 5 - Pandenchthys stolbovt, 6 - Eusthenopteron foordi, 7 - Gyroptychius pault 1-2 - исходные типы fe ench - хоана, V0 - сошник Psp - парасфеноид
Приведенные примеры не просто подтверждают необходимость обращения к вымершим предковым формам, когда речь идет о гомологизации органов существующих ныне их потомков Вопрос надо ставить глубже. Ведь и сами эти вымершие формы мы подбираем с учетом филогенетических представлений Мы не можем набирать переходные формы, не позаботившись о том, чтобы наши сравнения проходили в определенных таксономических рамках. Это означает, что надежность гомологизации зависит от филогенетических концепций
Противники существования тесной зависимости гомологизации от установления филогенетической преемственности таксонов обычно подчеркивают, что сами филогенетические представления в конечном счете опираются на то, как мы гомологизируем органы. Операция гомологизации считается первичной, а исторические реконструкции - производными. Отчасти так оно и есть. Сначала проводят гомологию между семенами гинкго и тисса (гиоидным аппаратом рыб и органом слуха наземных позвоночных и т. д.), а лишь затем говорят о сходствах и отличиях соответствующих организмов, осуществляют распределение их по таксонам и высказывают суждение о филогенетической связи последних. Но таким способом мы получаем провизорные таксоны и архетипы. При более детальных исследованиях подобные прямые гомологии могут оказаться ошибочными
Между операциями гомологизации, таксономического отождествления и установления филогенетической преемственности имеется тесная обратная связь. Складывается некий общий контекст, в котором уже бессмысленно считать одни положения логически первичными (фундаментальными), а другие - логически вторичными (производными). Тогда уже нельзя говорить, что мы сначала гомологизируем органы, а затем судим об их происхождении, как нельзя сказать и того, что только обращение к филогении позволяет проводить гомологизацию органов. Обсуждение того, что раньше и что важнее в этих операциях, лишено смысла.
Обнаружение вымерших переходных форм между современными таксонами позволяет ставить задачу анализа исторического становления формы в ракурсе исследования характера и динамики преобразования морфологических структур в эволюционных рядах. Это одна из центральных задач эволюционной морфологии, распространяющаяся на все организмы и на все их компоненты. Едва ли нужно пояснять, что для работы в этом направлении палеонтологические данные совершенно незаменимы.
Установление рядов морфологических переходов в процессе эволюции - важнейший инструмент и репер в изучении разнообразия организмов, поскольку, исследуя переходы, мы ставим их в соответствие с филогенетическими связями форм. Отдельные морфологические понятия, соответствующие органам различного типа, оказываются при этом объединенными в логическую систему. Следующий шаг в том же направлении - установление инвариантности преобразований, происходящих в разных таксонах, далее - сворачивание наблюдаемого многообразия до лаконичных схем или формул, где отдельные модальности формы выступают как значения переменной
Такого рода исследования предпринимали неоднократно, но обычно выполняли скорее в рамках не собственно морфологии, а в рамках систематики и эволюционного учения (имеются в виду исследования явления, называемого гомологическими рядами наследственной изменчивости, аналогической изменчивостью и т. п.). В результате исследователей чаще всего интересовала не столько упорядоченность изменчивости данного органа в морфологическом ряду, сколько сам феномен повторения разновидностей одного органа в разных таксонах, например, вариабельность в относительных размерах и протяженности сошника и парасфеноида в разных отрядах, семействах и родах кистеперых рыб (см. рис. 3). Для морфологии же более важна именно упомянутая упорядоченность форм одного ряда, тот факт, что и набор разных форм, и правила преобразования, которые связывают формы одного ряда, удерживаются в разных таксонах. Такие морфологические ряды, для которых удается установить правило преобразования форм, подчинить и выяснить изменчивость и определенную закономерность их изменчивости, названы рефренами [651, 1952].
Рефрены можно продемонстрировать на рядах расчленения листьев современных и ископаемых растений (рис. 4). Переход от простого листа к перистому, пальчатому или дихотомическому и к более сложным типам расчленения (как и обратное упрощение расчленения) осуществляется у всех растений примерно по одним правилам. При этом не имеет значения, наблюдается ли этот переход на разных листьях одной особи, в онтогенезе одного листа, у разных особей одного или разных видов, в синхронном срезе или в филогении. Разнообразие листьев по признаку их расчленения укладывается в ограниченное число типов, за пределы которого морфологические преобразования в принципе не могут "вырваться". Установив эти немногие правила преобразования, мы свертываем наблюдаемое разнообразие листьев до значений некоторой переменной. Рефрены устанавливаются в формах раковин фораминифер и головоногих моллюсков, в морфологии зубов и хвостовых плавников эласмобранхий (рис. 5), в разных системах органов современных ресничных червей-турбеллярий, в преобразованиях альвеолярных желез у млекопитающих, онто- и филогенетических рядах усложнения складчатости зубов у кистеперых рыб группы рипидистий. У последних многообразие усложнения зубной структуры охватывается тремя типами (эустенодонтным, полиплакодонтным и дендродонтным), связанным переходами (рис. 6). Различия между этими типами сводятся к трем параметрам: сложности в "ветвистости" складок денгина; заполнении пульпарной полости остеодентином; степени проникновения периферической кости между складками дентина [2432] либо, напротив, их полного смыкания.
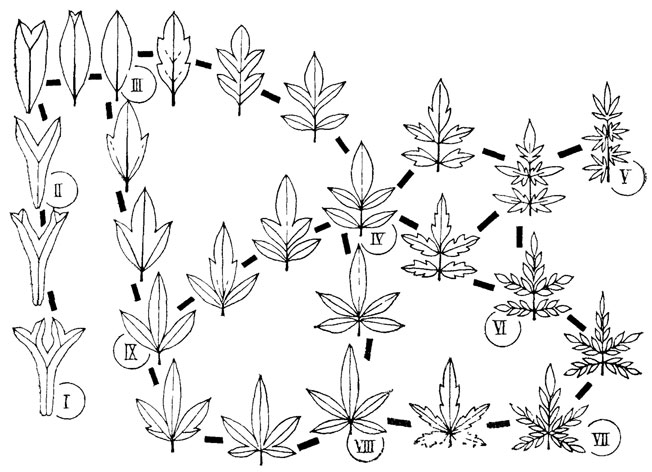
Рис. 4. Схема расчленения листьев и листоподобных органов высших растений. По С. В. Мейену. Главные модусы преобразования: дихотомирование (III-I), пиннация (III-IV-VI; IX-VI; VIII-VI; VIII-VI) и пальмация (III-IX-VIII; IV-VIII; IV-V; IV-VI-VII); к типам листьев V и VII приводят сочетания пальмации и пиннации

Рис. 5. Морфология и эволюция хвостового плавника у палеозойских эласмобранхий (вариации и сводимость к трем основным типам). По Р. Цангерлу. а - исходное состояние; б - Goodrichthys eskdalensis; в - Badringa herdiпае; г - Hybodus hauffianus; д - Cobelodus aculeatus; е - ксенакант из нижнего карбона; ж - Triodus sessilis; з - Cladocelache fyleri; и - Romerodus orodontus; к - Caseodus basalis; л - Eugeneodus. richardsoni
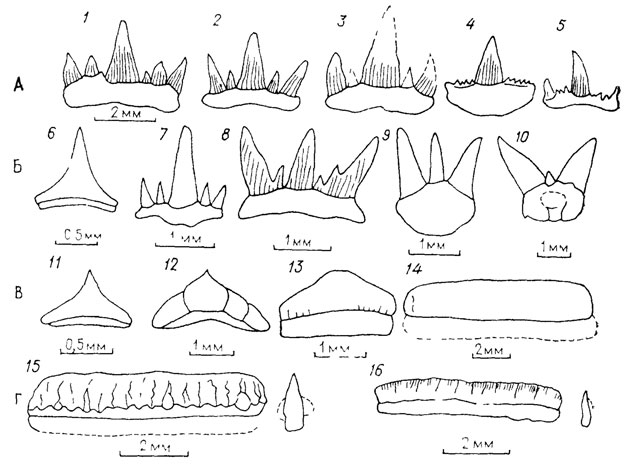
Рис. 6. Четыре типа формы зубов и их вариации у палеозойских эласмобранхий. По Р. Цангерлу. Тип А: 1-3 - Cladodus mirabilis (индивидуальные вариации), 4, 5 - Cladodus striatus (индивидуальные вариации); тип Б: 6 - Hybocladodus tenuicostatus, 7 - Cladodus exilis, 8 - Pnsticladodus strtngeri armalus, 9 - Trtodus dessilis, 10 - ксенакант; тип В: 11 - Hybocladodus pluatihs, 12 - Leiodus calcaratus, 13 - Orodus vancostatus, 14 - Caseodus basalts; тип Г: 15 - Eugeneodus richardsom, 16 - Gilliodus peyeri
Все эволюционные преобразования структуры зубов, начиная с кистеперых рыб и включая амфибий и рептилий, сводятся к комбинациям этих правил как в онто-, так и в филогенезе (рис. 7). Разнообразие очертаний раковин гастропод удавалось связать всего с двумя параметрами. Изменяя эти параметры, можно воспроизвести на аналоговом компьютере все основные реальные формы их раковин. Все разнообразие пыльцы и спор укладывается в конечное число типов, связанных очень простыми преобразованиями (перемещением апертур, изменением их числа и др.).
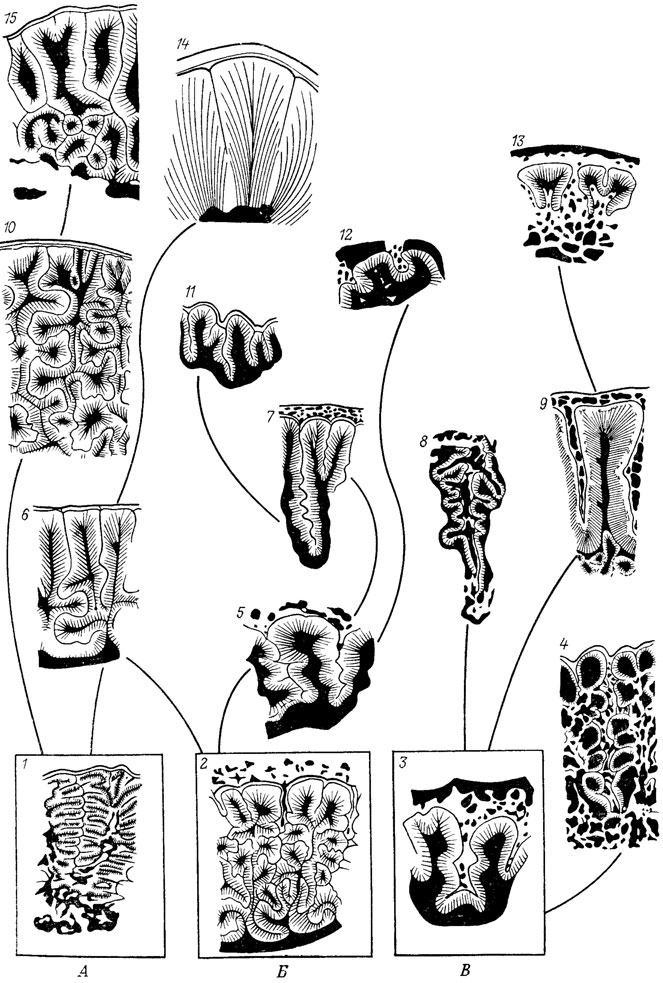
Рис. 7. Схема эволюционных преобразований зубной структуры позвоночных на основе трех исходных типов. Модифицирована Э. И. Воробьевой по Г. П. Шультце. Исходные типы А - дендродонтный, Б - полиплакодонтный, В - эустенодонтный. Кистеперые рыбы. 1 - Porolepis, 2 - Panderichthyas, 3 - Eusthenopyeron, 4 - Eusthenodon, 8 - Strepsodus, 9 - Pycnoctenius factitious; амфибии: 5 - Ichthyostega, 6 - Eogyrinus, 7 - Loxomma, 10 - Mastodonsaurus, 11 - Kotlassia; рептилии: 12 - Mixosaurus, 13 - Eurypteryguus, 14 - Mosasaurus, 15 - Varanus
Путем выявления подобных типов и их параметров можно подойти к выявлению того, что называют имманентными закономерностями формы организмов. К сожалению, такие исследования выполняют в незначительном объеме отчасти из-за того, что из самого факта упорядоченности форм в живой природе нередко делают слишком прямолинейные эволюционные выводы, утверждая предначертанность эволюции, отрицая связи организмов с экосистемной обстановкой и т. д. Кроме того, такой путь доступен далеко не для всех палеонтологических материалов и групп.
Следует подчеркнуть необходимость четкого разграничения морфологических переходов от таксономически переходных ("мозаичных") форм, обычно стоящих на стыке крупных таксонов: рыб и амфибий, рептилий и птиц, рептилий и млекопитающих. Встречаемые в палеонтологической летописи так называемые переходные ("мозаичные") формы оказываются нередко боковыми тупиковыми ветвями (например, археоптерикс и ихтиостега, многие стегоцефалоподобные кистеперые рыбы). Они обнаруживают сложную мозаику примитивных и продвинутых морфологических признаков, которые нередко сочетаются с чертами особой специализации. Так, анализ более 30 скелетных и других признаков у археоптерикса [2207] показал, что большая их часть оставалась рептилийной, меньшая птичьей и только единичные признаки занимали действительно переходное положение между этими группами.
Морфофункциональный анализ
Многие аспекты морфофункционального анализа разбираются в специальных главах (см. 1.2 и 1.3), а также в главах, посвященных палеофизиологии, палеоэкологии и реконструкции внешнего облика организмов (см. 1.4, 1.6, 3.1). Здесь этот анализ будет рассмотрен лишь как составная часть морфологического исследования. В морфологии хорошо разработана система понятий и исследовательских процедур, касающихся гомологизации частей, выявления архетипа и прослеживания исторического преобразования частей, но столь же разработанной теории морфофункционального анализа не существует, хотя литература, посвященная соотношению формы и функции, необозрима. Большая ее часть непосредственно исследует это соотношение и почти не касается применяемых исследовательских принципов.
Морфофункциональный анализ доэволюционной морфологии унаследовал идеалы и принципы раннего телеологического рассмотрения живой природы, когда каждому органу, каждой черте организмов ставилась в соответствие предустановленная свыше цель. Место последней в рамках селекционистского эволюционизма (от классического дарвинизма до синтетической теории эволюции) заняла адаптивность, ведомая и контролируемая естественным отбором. Тем самым морфофункциональный анализ стал связующим звеном между морфологией и эволюционной теорией. Задачей его стало не только вскрытие функции органов, но и установление тех факторов отбора, которые ответственны за становление и эволюцию органов. Проблема соотношения формы и функции дополнилась проблемой приспособления, обусловленного борьбой за существование. В этой роли морфофункциональный анализ выступает и сейчас, соединяя морфологию с экологией, физиологией, теорией эволюции и таксономией.
Морфофункциональный анализ интересует нас как некоторая специально организованная исследовательская деятельность, а не просто имеющая место практика легковесного приписывания органам неких функций и их адаптивности без сопутствующего исследования того, действительно ли орган несет ту или иную функцию и какую биологическую роль орган выполняет. Поверхностное отношение к морфофункциональному анализу сослужило плохую службу и скомпрометировало его настолько, что целые крупные морфологические школы фактически отказывались обращаться к этому виду исследований, полагая, что там, где начинается "физиология", кончается морфология. На этой позиции стоял, в частности, один из виднейших морфологов растений В. Тролль; звучат такие представления и у некоторых современных морфологов, занимающихся позвоночными животными.
Трудности теоретического осмысливания морфофункционального анализа начинаются с раскрытия основных понятий, прежде всего самого понятия функции. В самом общем смысле функция в биологии - компонент (аспект, вид) материального взаимодействия между частями организма, а также между организмом и той экосистемой, в которую он входит, хотя здесь все чаще говорят об адаптивной роли органов [416, 1349]. Совокупность функций образует функциональный архетип, который лишь отчасти соответствует морфологическому архетипу. Установление соответствий между обоими архетипами и есть морфофункциональный анализ. Этой цели служат следующие исследовательские приемы: 1) прямое наблюдение и эксперимент; 2) установление корреляций между органом и какой-либо из функций, в том числе при повреждении или отключении органа (это принцип "черного ящика"); 3) использование технических или иных моделей (сведения, полученные на модели, экстраполируются на изучаемый орган; в качестве модели могут выступать и сами организмы); 4) сравнительно-функциональный анализ, когда функциональные выводы делаются по аналогии с выводами, полученными на сходных органах других групп организмов.
На использовании перечисленных приемов морфофункционального анализа в отношении рецентных организмов мы не будем останавливаться и сразу обратимся к палеонтологическому материалу. Прежде всего отметим, что у вымерших организмов с не встречающимися ныне типами органов (например, челюсть-"корыто" у платибелодона, "койи" у носорога-халикотерия) могли быть связаны и исчезнувшие ("вымершие") функции. Вполне вероятно также, что для многих органов вымерших организмов нам никогда не удастся определенно установить их функции (например, рогоподобных костных выростов на черепе Uintathenum из диноцерат). Еще труднее установить такое различие в функциях между разными модификациями органов, из которого можно вывести определенный фактор отбора. Это совсем иная задача, чем просто связать орган с функцией. Например, ясно, что экзина спор и пальцы осуществляет защитную функцию, однако мы не имеем представления о том, какие преимущества могли дать растению определенная структура экзины, однолучевая или трехлучевая апертура, общее очертание зерна и подавляющее большинство других признаков. Трудно указать, какие функциональные преимущества давала утрата дентиноподобных тканей в наружном скелете рыб, та или иная закрученность раковины моллюска или разный характер микроструктуры зубов у древних амфибий. Мы не можем априорно исключать возможность, что многие аспекты разнообразия организмов вообще не могут рассматриваться как прямой ответ на определенное селективное давление. Хотя приемы морфофункционального анализа, используемые в палеонтологии, те же, что и при изучении с его помощью рецентных организмов, но их относительная роль иная. В палеонтологии невозможно обратиться к прямому наблюдению над функционированием органа, так же как дать оценку его действительной адаптивной роли. В лучшем случае можно наблюдать исполнение функции как бы на моментальном снимке (например, проросшую пыльцу в пыльцевой камере семезачатка; прикрепление раковины брахиопод к субстрату с помощью ножки: нахождение эмбриона ихтиозавра в чреве матери и т. п.). Весь материал прямых наблюдений привносится в палеонтологический морфофункциональный анализ из неонтологии, поэтому актуализм, а точнее, функциональные сопоставления ископаемых и рецентных форм, оказываются одним из главных исследовательских принципов данного анализа в палеонтологии. То же касается и установления корреляции. Правда, возможности палеонтолога несколько шире, когда речь идет об экологической функции. Часто удается констатировать выдерживающуюся корреляцию между определенным строением органа и определенной особенностью экосистемы. Например, присутствие воздухоносных каналов в корнях растений коррелирует с литологическими признаками сильно увлажненных субстратов. Поэтому можно предполагать, что у вымерших растений, как и у современных, эти воздухоносные каналы обеспечивали аэрацию корней в условиях плохо аэрируемой почвы. Переходы от брадиодонтности к гипсодонтности у хоботных и от пятипалой к однопалой конечности у копытных хорошо увязываются с изменениями климатических и ландшафтных условий их обитания.
Для выяснения и оценки функциональных характеристик органов иногда используют моделирование. Например, на искусственных моделях изучалась гидродинамика раковин аммонитов. На увеличенных копиях семян исследовалась динамика воздушных потоков со взвешенной в них пыльцой для установления функционирования выростов на покровах семезачатков и несущих их органах. Используют также математическое моделирование.
Все эти методы морфофункционального анализа в палеонтологии имеют, однако, подчиненное значение по сравнению с актуалистическими параллелями. Чаще всего палеонтолог проста экстраполирует на свой материал данные, получаемые на современных организмах. Этот метод экстраполяции выводов широко применяется и в морфофункциональных исследованиях рецентных форм; он необходим и даже неизбежен, хотя именно он наиболее уязвим теоретически. Дело в том, что многолетний опыт морфофункционального анализа рецентных форм ясно показывает отсутствие взаимно-однозначного соответствия (изоморфизма) между морфологическими и функциональными меронами, тем более, что многие функции остаются неизученными. В наиболее общем случае каждому морфологическому мерону может быть поставлено в соответствие более одного функционального мерона, и наоборот. Иными словами, связь формы и функции достаточно сложна и не может быть описана такими однозначными зависимостями, которые допускают неограниченные экстраполяции. Поэтому, установив функцию определенного органа в каком-то таксоне, нельзя утверждать, что и во всех других таксонах этот орган несет именно эту функцию или что при одинаковой функции орган выполняет ту же биологическую задачу, или что все признаки сложных адаптивных комплексов адаптивны.
Даже органы с одной резко доминирующей функцией могут одновременно использоваться для дополнительных, иногда с трудом устанавливаемых функций и выполнять несколько биологических ролей. Например, рога копытных, клюв и оперение птиц либо глаза, которые помимо зрения служат для синхронизации и регуляции метаболизма, гормональных и ферментативных процессов [1525], а у высших позвоночных играют еще и этологическую роль, - участвуют в мимике угрожающего поведения, привлечения брачного партнера и т. д. Вместе с тем одна и та же функция может иметь разный адаптивный смысл в разных группах организмов. Примерами здесь могут служить сходный принцип (хватательно-прикрепительные устройства эпителиально-мышечной природы) мужских совокупительных органов и хоботков ресничных червей [1815] и собственно прикрепительные органы плоских червей, в том числе паразитических. Среди позвоночных иллюстрацией могут служить сходные пропорции конечностей, приспособленные для разных форм бега, или сходные функции квадратно-мандибулярной систем у хамелеонов и слепозмеек, используемых для разной биологической роли: у первых выдвижение нижней челюсти вперед обеспечивает свободу выбрасывания языка, у вторых челюсть подсовывается под добычу при слабом открывании пасти [446].
В. А. Догель [326] предлагал различать механизм функционирования тех или иных образований и их биологическую роль, показывая, что поскольку морфологический субстрат несет ограниченный набор функциональных единиц, то "способы сборки" и "способы строительства" из одних и тех же "деталей" могли повторяться. Положение о необходимости включения в морфофункциональный анализ специального использования функций (как специфического действия структур) и соотношений их с биологической ролью органов практически общепринято [500, 1350, 1617]. При этом подчеркивается, что морфофункциональный анализ не должен предоставлять собой лишь дедукцию из особенностей морфологии, что в особенности касается палеонтологического материала.
Прекрасным примером трудности выяснения функции органов у вымерших организмов даже при возможности актуалистических сопоставлений может служить сифонный аппарат древних головоногих. В настоящее время достаточно хорошо поняты строение и функции сифона современного наутилуса. Однако у ряда групп из раннего палеозоя сифон был устроен значительно сложнее. Так, у эндоцератид так называемый сифон мог достигать значительной ширины и, вне всякого сомнения, включал в себя значительную часть заднего конца мягкого тела. Иногда в таком "сифоне" имеется даже сложная система каких-то вертикальных "стержней".
К. Тейхерт еще в начале 30-х годов описал у актиноцератоидей очень сложную систему из трубочек, что послужило поводом для длительной дискуссии. Было высказано предположение, что это следы сложной кровеносной системы, существовавшей в сифоне. Позднее в правильности данного предположения стали сомневаться, так как по остальным особенностям названная группа принадлежит, видимо, к достаточно архаичным головоногим. Высказывалось мнение, что структуры, принимаемые за трубочки, являются просто местом смыкания разрастающихся внутрисифонных отложений (довольно частых у древних головоногих). Однако в последнее время Ф. А. Журавлева на девонских актиноцератоидеях удивительной сохранности вновь обнаружила совершенно реальные трубочки в сифоне. Очевидно, что так называемые "сифоны" эндоцератоидей и актиноцератоидей были полифункциональными и играли несколько иную роль по сравнению с сифоном современного наутилуса и ископаемых наутилид.
Сложные случаи отношения формы и функции объединяются понятиями морфофункционального параллелизма (одна функция при разных формах), морфофункционального полиморфизма (разные функции у одной формы), полиэкологичности функции и параконвергентного параллелизма (сходство специфических функций при разной адаптивной роли). Первое понятие привлекало внимание морфологов преимущественно в отношении аналогии (исполнение одной функции органами, не являющимися гомологичными). Второе понятие анализировалось более интенсивно и охватывает разные явления. Это мультифункциональность (одновременное исполнение органом разных функций), смена функционирования - попеременное использование органа для разных функций [1134], смена функции - смена основного типа функционирования в истории органа [1512], иерархичность функции - ее интегративность на разных структурных уровнях органа [500].
Третье - предполагает "разрывы" между функцией и ее экологическим значением, обеспечивая множественность адаптации и лабильность связей организма со средой [824]. Четвертое предполагает устойчивость функциональных меронов [446].
В палеоботанике хороший пример аналогии функций дают голосеменные. В ходе их эволюции можно видеть (особенно в классе Cycadopsida), как функция захвата пыльцы переходит от сальпинкса у наиболее примитивных форм к интегументу, затем к купуле, преобразованной во внешний интегумент, и, наконец, к рыльцу уже у покрытосеменных. Эти воспринимающие органы заведомо не могут быть гомологичными по критерию положения, так как относятся последовательно ко все более внешним оболочкам вокруг мегаспорангия.
Хорошие примеры морфофункционального полиморфизма можно найти и в палеозоологии.
Все перечисленные явления могут сочетаться друг с другом в разных комбинациях. Так, экзо- и эндоскелет позвоночных, как известно, выполняет опорную и защитную функции. Существует представление, что последняя функция была исходной и первично скелетные образования возникли в голове в связи с защитой органов чувств [1619], т. е. в связи с сенсорной функции скелета. Предполагается также, что у древних рыб экзоскелет выполнял электросенсорную функцию. Кроме того, у многих рыб он участвовал в сейсмосенсорной функции, поскольку соответствующие каналы проходили в экзоскелете. Последняя функция могла утрачиваться в процессе эволюции. Например, у рецентных дипной сейсмосенсорные каналы переместились в кожу. Опорная функция также претерпевала изменение в истории группы: у древних (в частности, у панцирных) рыб она выполнялась преимущественно экзоскелетом, у более поздних - эндоскелетом.
Вопрос о том, как при работе с вымершими таксонами обойти трудности, порождаемые морфофункциональным полиморфизмом, освещен в литературе весьма слабо. В качестве общего рецепта можно рекомендовать поиск независимых показателей функции данного органа, а не просто перенос функций с современных организмов. Это тем более важно, что прямые экстраполяции здесь чреваты ошибками. Примером может служить реконструкция у кистеперых рыб по аналогии с современными амфибиями оперкулярной косточки [1796], которая является скорее новообразованием у тетрапод в связи с формированием у них наземной оперкулярной слуховой системы. Косвенным свидетельством здесь может служить то, что у ряда современных хвостатых и бесхвостых амфибий в связи с переходом к водному образу жизни opercula вторично утрачена [963, 964].
Независимые указания на функцию может дать в ряде случаев тафономическое и вообще палеоэкосистемное исследование [298]. Функциональный смысл меронов становится более понятным, когда мы помещаем организм в конкретную экосистему. Так, морфофункциональный анализ конечностей лошадиных обрел убедительность только тогда, когда был подкреплен реконструкцией палеоландшафтов, населенных предками современных лошадей. Разумеется, сами палеоэкосистемные реконструкции также широко используют результаты морфофункционального анализа. Здесь мы снова сталкивается с обратной связью разных направлений и методов исследования.
Наконец, морфофункциональный анализ в палеонтологии может в известной мере строиться на сравнительно-морфологических исследованиях по ходу эволюции разных филумов. Такие исследования дают материал для выводов о биологической значимости структур и особенностях их функционирования. Например, в разных филумах кистеперых рыб в процессе эволюции наблюдается замена лигаментозных связей нёбно-квадратного комплекса с мозговой коробкой на суставные и синхондрозные, увеличение числа этих связей и изменение их положения, сопровождающееся упрочнением контакта черепных блоков и консолидацией покровных костей крыши нёба и щечной области [186].
Эти изменения явно указывают на усиление акинетичности черепа в эволюции кистеперых, что соответственно привело к изменению функций и отдельных его конструкций. На базе этих морфофункциональных перестроек формировался стегальный череп древних амфибий.
Выводы, получаемые с помощью морфофункционального анализа, в палеонтологии интегрируют палеоморфологическое знание с морфологией современных форм, с физиологическим и экологическим знанием и с эволюционной теорией. Морфофункциональный анализ позволяет поместить вымершие организмы в конкретные палеоэкосистемы и более правильно реконструировать последние. Если учесть, что нынешняя стратиграфия все в большей степени превращается в экостратиграфию, то чисто практическое значение морфофункциональных исследований станет очевидным. Через палеоэкологию морфофункциональный анализ проникает в разработку проблем глобальной экологии, палеобиогеографии, палеогеографии, седиментологии и многих других. Во всех этих случаях морфофункциональный анализ служит важным компонентом причинно-следственного (каузального) исследования, которое по давней естественнонаучной традиции и по самой логике научной работы завершает любое исследование природных эволюционных систем.
Модусы морфологических преобразований
Прослеживание морфологических преобразований в истории филумов позволяет выявить ряд чисто эмпирических закономерностей или тенденций развития, которым нередко придают общебиологический и общеэволюционный смысл и описывают как правило, принципы, законы или закономерности.
Прежде всего нужно отметить, что в трактовке этих понятий биологами существует изрядная путаница, что ведет к трудностям в их упорядочении. Кроме того, в современной логико-гносеологической литературе эти понятия интерпретируют также самым разным образом. Принцип морфологических преобразований воспринимается нами с методологических позиций: как некоторое основополагающее положение в процессе познания, которое обобщает некую познавательную операцию и приписывает способ исследования с целью дальнейшего развития теории, основы которой он составляет. Закон строго и точно описывает результаты исследования, он формулируется для идеализированных объектов и поэтому не может иметь исключений (по определению).
В эмпирическом исследовании, имеющем дело не с идеализированными, а с реальными объектами, проявление закона регистрируется как статистическая устойчивость определенных связей между разными объектами и явлениями, охватываемыми данной теорией. Такая статистически устойчивая связь и называется закономерностью. Она выступает как результат проекции идеального отношения на реальный мир. Отклонения (то, что принято называть исключениями) не вынуждают отказываться от самой закономерности. Правилами можно называть такие повторяющиеся эмпирические совпадения, которые пока не связываются с действием какого-либо закона.
Если исходить из такого разграничения, то многие явления, которые описывают как принципы, следует перевести в число закономерностей или правил. С другой стороны, принципы морфологии не выявлены с такой полнотой, чтобы их можно было свести в логически связанный и достаточно полный список.
В качестве морфологических принципов, правил или законов выдвигали многие положения: историзм, актуализм, единство типа, целостность, корреляцию, единство формы и функции, рекапитуляцию, тройной пареллелизм, конвергенции, олигомеризацию, полимеризацию, исходное морфологическое многообразие, адаптивную радиацию, мозаичность, опережение типового состояния, фетализацию, гетеробатмии и др. Все эти обобщения не могут считаться исключительно морфологическими и обычно рассматриваются в контексте теории эволюции. Кроме того, они весьма разнородны по природе, многие из них, строго говоря, не являются принципами. В большинстве случаев это скорее утверждения об организмах и особенности их эволюции, а не предписание для исследования и развития теории. Разумеется, все перечисленные выше обобщения отчасти служат также руководством к действию, указывая на направление исследований. Так, к типично морфологическим принципам относится тройной параллелизм, когда он рассматривается не как явление, а как рабочий инструмент. Специфически морфологическим можно считать критерий положения в гомологии, два других ее критерия в равной степени свойственны таксономии.
Хотя большинство указанных явлений действительно должны охватываться теорией эволюции, они имеют отношение не столько к эволюции целых организмов и онтогенезов, сколько к определенным аспектам морфологических преобразований (тех или иных органов). В самом деле, едва ли можно говорить о конвергенции и параллелизме целых таксонов, поскольку соответствующие преобразования касаются лишь отдельных признаков некоторых меронов. Например, в классическом примере конвергенции рыб и водных млекопитающих вторично возникающее сходство у последних не касается их наиболее существенных диагностических признаков. В результате конвергируют не таксоны, а в основном контуры тела входящих в таксоны организмов. То же самое касается и сходных состояний структур, независимо достигаемых в относительно близких филетических линиях ископаемых и рецентных организмов, например, среди позвоночных сходное состояние отростков в носовой капсуле ископаемых кистеперых рыб и современных амфибий или гистоструктур зубов среди кистеперых, амфибий и рептилий [2432, 2434]. Также обстоит дело и в отношении многих других эволюционных модусов, которые, безусловно, следует рассматривать в рамках эволюционной морфологии, а не только филогенетики (поскольку речь идет о структурах).
В данной главе невозможно даже кратко проанализировать модусы морфологической эволюции, хотя палеонтология дает для познания многих из них весьма существенные сведения. Мы остановимся лишь на тех модусах, значение которых часто недооценивают и которые особенно хорошо прослеживаются именно на фоссильном материале либо представляют в палеонтологических исследованиях особый смысл. Это сальтационные морфологические преобразования, гетеротопии (и связанные с ними преобразования), дедифференциация органов и транзитивный полиморфизм органов, опережение структурных состояний в филумах.
Открытие переходов в палеонтологических и сравнительно-морфологических рядах породило убеждение, что все морфологическое разнообразие организмов в конечном счете можно свести к системе градуалистских преобразований. Соответственно древний принцип "природа не делает скачков" был положен в основу не только эволюционных, но и морфологических исследований. Именно на нем покоится принцип гипотетического единого предка и единства структурных типов в палеозоологии и наиболее популярная среди ботаников и палеоботаников теломная теория. Последняя ставит задачу выведения всего многообразия форм высших растений из одной исходной формы путем приложения немногих модусов преобразования. Если у одного растения спорангии расположены на концах осей, а у другого на боковом придатке, то строится гипотетический ряд, в котором спорангий постепенно "переползает" с одного места на другое. То же касается "перемещений" плавников и зубных спиралей у древних рыб. Такое мысленное конструирование длинных рядов переходных форм весьма обычно в морфологических исследованиях, хотя универсальность подобного подхода никогда не была убедительно показана.
Между тем можно привести немало случаев, когда градуалистское преобразование органов и признаков немыслимо даже теоретически. Например, образование целых проростков с корнями ("деток") из клеток краевой меристемы листьев у каланхое (бриофиллум) - морфологическая особенность, которая не могла появиться градуалистски. Появление эпифильных проростков у каланхое и других толстянковых - один из характерных признаков семейства - в морфологическом плане было явно сальтационным, хотя в плане таксономическом и филогенетическом могло стать характерным для таксона достаточно постепенно: сначала в виде аберраций и лишь затем в качестве нормы. Это разграничение таксономически-филогенетического и морфологического аспектов сальтационных преобразований особенно важно.
Нарушение постепенности в историческом преобразовании органов происходило, очевидно, тем легче, чем слабее были сложившиеся корреляционные связи между органами в онтогенезе организма. Чем более независимы отдельные онтогенетические блоки, тем легче могут меняться пространственные (гетеротопия) и временные (гетерохрония) их отношения, тем легче нарушается непрерывность морфологических рядов. Соответственные ограничения налагаются как на критерий положения, так и на критерий переходных форм: положение изменяется скачком, а переходы вообще отсутствуют.
Подобные скачкообразные изменения системы, сменяющие периоды гомеостаза, могут изменить доминантные элементы (блоки) всего организма в течение нескольких поколений [1529]. Реконструировать морфологические скачки особенно трудно, так как всегда остается возможность того, что мы просто еще не открыли переходных форм между крайними вариантами. Большую помощь здесь может оказать обращение к тератологическому материалу, к мутационным преобразованиям, изучаемым генетикой, явлениям рекапитуляции и реверсии, наблюдаемым на сравнительно-анатомическом и эмбриологическом материалах. Таким путем можно хотя бы составить общее представление о том, какие способы сальтаций принципиально возможны и действительно реализуются, насколько обычны подобные явления и к каким последствиям для организмов они приводят. Однако реальные семофилогенетические преобразования, протекающие скачкообразно (по типу пороговых реакций), удается констатировать подчас только с помощью палеонтологии. В качестве примера можно привести интенсивное ветвление затылочных вен на крыше черепа у ехидны и бесхвостых амфибий, которое до сих пор встречено среди вымерших позвоночных только у палеозойской кистеперой эустеноптерон [186].
Изучая последовательные ряды ископаемых форм, нередко удается проследить достаточно постепенные перестройки органов. Это мы видим, например, при возникновении аммоноидей от бактеритоидей. Известен ряд форм от совершенно прямого Lobobactrites до свернутого в полную спираль аммонита. Переходы столь полны, что до настоящего времени нет единства мнений о ранге и систематическом положении бактритов. Таково также растянувшееся на несколько эпох формирование семеносных листьев (филлоспермов) в специализированные семеносные органы (кладоспермы) в классе Ginkgoopsida (рис. 8). Однако само образование филлоспермов из более ранних безлистных семеносных органов едва ли происходило столь постепенно. То же можно сказать о спорофиллах папоротников или появлении короноидоподобной адсимфизной пластинки на переднем конце нижней челюсти вместо обычной шагреневой у большинства остеолепид [189]. Эта пластинка топографически соответствует симфизной пластинке у поролепид (рис. 9) и зубным спиралям у некоторых эласмобранхий (например, у геликоприона). Морфологических переходов здесь практически нет и можно предполагать сальтационное появление всех этих образований. Косвенным свидетельством здесь может служить факт присутствия шагреневого выступа (рудимента адсимфизной пластинки) у некоторых стегоцефалов, сохраняющих, таким образом, примитивное состояние. Во всех подобных случаях наблюдается внезапное появление в геологической летописи новообразований.
Рис. 8. Происхождение воротничка гинкговых от семеносных вай (филлоспермов) каллистофитовых По С. В Мейену. а - филлосперм Dicksonttes pluckenetu (Schloth ) Sterz из среднего карбона, б - модифицированное семеносное перышко D pluckenetu (Schloth ) Sterz из верхнего карбона, в - сильноредуцированная семеносная вайя (кладосперм) Biarmopiens pulchra Zal (Peltaspermales) из нижней перми, г - то же, семеносные перышки, д - двусеменный кладосперм Peltaspermum thomasu Harris (Pcltaspermales) из триаса, е - парные купуловидные кладоспермы Pilophorosperma (Peltaspermales) из триаса, ж - парные семезачатки с воротничками современного Ginkgo biloba L. Семенные рубцы (а-в) и семезачатки (г-ж) зачернены
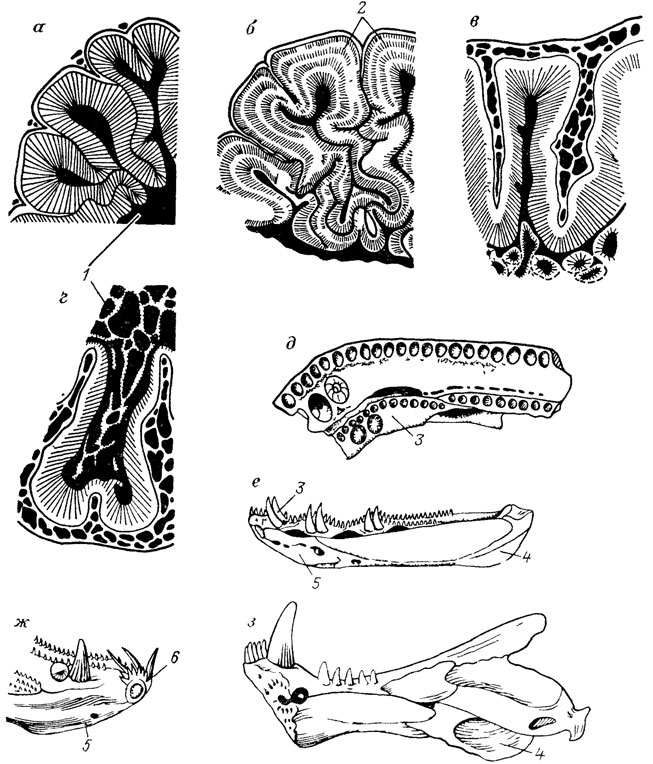
Рис. 9. Морфологические сальтации и параллелизмы среди позвоночных По Э. И. Воробьевой. а-г - поперечные шлифы через основание челюстных зубов (1 - пульпа, 2 - складчатый Дентин), а - девонской кистеперой рыбы Pandertchthys, б - девонского стегоцефала - ихтиостеги, в - каменноугольной кистеперой рыбы Pycnoctenwn, г - пермского ихтиозавра, д-ж - разные состояния развития адсимфизной пластинки (3) кистеперых рыб и ее аналога пресимфизной пластинки (6). д - короноидноподобная адсимфизная пластинка девонского Obruchevtchthys, е - шагреневая пластинка (примитивное состояние) девонского Platycephaltchthys, ж - пресимфизная 'спираль' девонских поролепид, е, ж - параллелизмы у кистеперых рыб в развитии переднего мандибулярного фланга (5), е, з - параллелизм у кистеперой рыбы (е) и териодонта (з) в развитии заднего ангулярного фланга (4)
В ряде случаев предшествующие и последующие состояния структур распространены на значительных временных интервалах, поэтому и переход между ними, если бы он был градуалистским, должен был бы растянуться и выразиться в серии переходных форм. Этого мы как раз и не видим. В частности, пока нет доказательств постепенного образования ростра белемнитов. Известны группы внутреннераковинных, у которых весь ростр состоит из не скольких слоев, облекающих фрагмокон. Но это особые группы и переходы между ними и типичными белемнитами отсутствуют; это небольшие слепые ветви в развитии внутреннераковинных. Но серии форм с постепенно возрастающим размером ростра в группе белемнитов в узком смысле слова пока нет.
Интересный пример дают аммоноидеи. Протоконх отделяется от остальной камерной части раковины первой септой-просептой. В процессе филогенеза у аммоноидеи сменяли друг друга три формы первой лопастной линии (края просепты) - просутуры: аселлатная - почти прямая линия (девон); латиселлатная - с широким седлом на вентральной стороне, соответствующим ему седлом на дорсальной стороне и одной или двумя лопастями между седлами на каждой стороне (девон - триас); ангустиселлатная - с узким вентральным седлом, дорсальным седлом и двумя или тремя лопастями между ними (триас - мел). Такая форма характерна для первой лопастной линии. Постепенности превращения просутур пока не отмечено.
Разумеется, в подобных случаях всегда можно отнести прерывистость в рядах форм за счет неполноты палеонтологической летописи. Если именно эта неполнота ответственна за отсутствие переходов, то в общем случае полнота рядов должна находиться в прямой зависимости от частоты встречаемости тех остатков, из которых строятся ряды. Соответственно на довольно редко встречающихся фруктификациях высших растений должны получаться гораздо более неполные ряды, чем, например, на остатках спор и пыльцы. Однако такая зависимость наблюдается далеко не всегда. Так, описанный выше переход от филлоспермов к кладоспермам достаточно постепенный, не требующий гипотез о сальтационных перестройках, а происходящие в том же филогенетическом ряду перестройки пыльцы такой постепенности не обнаруживают. Свойственная палеозойским представителям класса Ginkgoopsida ребристая экзина появляется в геологической летописи внезапно, без каких-либо переходных форм. Если учесть, что пыльца захороняется в породах и извлекается из них в аналитических целях в огромных количествах, то отсутствие переходов от гладкой экзины к ребристой трудно связать с одной лишь неполнотой геологической летописи.
Рассмотренный пример показывает, что установление сальтационных преобразований требует параллельного анализа многих органов в филогенетических рядах. О сальтациях признаков можно говорить лишь в таком последовательном ряду форм (таксонов), в котором его достаточно высокая непрерывность показана и по другим признакам.
Эволюция семеносных органов гинкгоопсид интересна еще и тем, что произошедший здесь переход фруктификаций с безлистных органов на листья приводит к появлению органов нового типа, которые уже не могут быть описаны в прежних терминах. Нарушаются сразу все три критерия гомологизации, так как предшествующие органы оказываются в новом пространственном соотношении (критерий положения неприменим), меняют собственные признаки (лист с семенами уже не прежний вегетативный лист) и нельзя говорить о переходных формах (их нет). Следовательно, появившийся орган требует нового морфологического понятия.
По-видимому, гетеротопии - перемещение органов с нарушением критерия положения - могут сопровождаться сменой собственных признаков и перемещающегося органа и тех органов в соседстве с которыми он оказывается. Такие преобразования можно назвать постгетеротопными. К ним можно причислить и упоминавшиеся ранее изменения места прикрепления нёбно-квадратного комплекса к мозговой коробке в ушной области черепа у разных кистеперых рыб и тетрапод (из вентрально-нижебоковой вены головы, к дорсальному - над веной, см. рис. 2), связанные, вероятно, с разным вхождением висцеральных элементов в состав черепа. Диапазон и длительность постгетеротопных преобразований органов могут быть очень значительными. В ряду преобразований органов размножения гинкгоопсид филлоспермы, неотличимые по морфологии их листовой компоненты от стерильных листьев, преобразовались в конечном счете за время от среднего карбона до юры в небольшой воротничок гинкговых, удержавшийся у современных форм. Утрата листовых признаков привела к восстановлению стеблеподобного облика семеносных органов. Аналогичные преобразования претерпели мужские фруктификации. В органах обоих типов как бы восстановилось состояние, свойственное далеким предковым формам. Органы размножения снова стали безлистными.
Такой возврат примитивного состояния, по-видимому, не может быть надежно реконструирован без обращения к ископаемому материалу. Возможны и другие способы возврата к примитивным состояниям, не зависящие от постгетеротопных преобразований. Видимо, один из наиболее распространенных способов - дедифференциация органов, отмеченная и у животных, и у растений [2141]. Дедифференциация имеет место в филогенезе низших позвоночных рыб и амфибий, когда мы наблюдаем в их скелете локальное вторичное появление кости на месте хряща. Дедифференциация среди ископаемых и рецентных позвоночных нередко осуществляется путем фетализации - сбрасывания поздних стадий онтогенеза.
В частности, за счет такого "сбрасывания", которое происходит мозаично, формируется широкий спектр звукопроводящей системы у амфибий. Возможно, что тот же механизм, свидетельствующий о пластичности системы, использовался и при формировании вариантов акустического аппарата у древних амфибий - стегоцефалов [964].
Очень глубокая дедифференциация органов наблюдается у хвойных. Еще у предков хвойных (в порядке Cordaitanthales) происходит дифференциация органов на вегетативные листья, брактеи, стерильные чешуи пазушного комплекса и семяножки. Эти четыре типа органов отличаются и по микроструктурным, и по макроморфологическим признакам (не только по критерию положения, но и по критерию специального качества). Уже у древнейших хвойных происходит частичная дедифференциация этих органов. Брактеи Lebachia становятся идентичными листьями на осях предпоследнего порядка, стерильные чешуи пазушного комплекса уподобляются листьям осей последнего порядка, более листоподобными становятся и семяножки. У более поздних лебахиевых (род Sashinia из верхней перми) все четыре типа органов идентичны по эпидермальным признакам и общему облику, отличаясь лишь величиной и по критерию положения (свое относительное положение на побеге они сохраняют). Даже семяножки уподобляются листьям и отличаются от них только загнутой верхушкой, под которой пряталось семя (рис. 10).
Рис. 10. Брактеи (1), семяножки с семенным рубцом (2) и стерильные чешуи пазушного комплекса (3) верхнепермских хвойных; У Ortiseia (А) перечисленные органы сильно дифференцированны, а у Sashinia (Б) - дедифференцированны. По И. А. Клемент-Вестерхаф
Возможность такой глубокой дедифференциации разных органов нельзя установить по современным хвойным. Поэтому она не учитывалась при интерпретации женских фруктификаций. Соответственно обнаружение листовых признаков у тератологических экземпляров семенных чешуи современных сосновых считалось доказательством того, что семенная чешуя - орган листового происхождения. В тератологических формах видели атавизмы, частичный возврат к предковому состоянию. Знание ископаемых форм ясно показывает, что указанные тераты - не атавизмы, а аберрантные формы, испытавшие частичную дедифференциацию органов.
Строя взаимосвязанные ряды форм, морфологи обычно стремятся свести наблюдаемое разнообразие каждого органа к одному исходному состоянию. Этот морфологический идеал соответствует принципу монофилии в таксономии и филогенетике и, казалось бы, стоит с ним в тесной связи. В действительности эта связь необязательна Дело в том, что у исходного филогенетически таксона тот или иной орган может быть достаточно полиморфным, хотя этому полиморфизму и не придается существенное таксономическое значение. Из этого исходного полиморфизма органа затем происходит выщепление различных доминирующих модальностей, каждая из которых может затем проявиться в таксонах-потомках. Существует представление, что различия в тех признаках, на которых основано выделение таксонов высокого ранга, первоначально выступали как внутрипопуляционная изменчивость - принцип генетического униформизма [2322]. Действительно, симметрия купулы была признаком очень невысокого веса у первых голосеменных, но в конечном счете стала одним из важнейших признаков, по которым различают классы Cycadopsida и Pinopsida [1955]. Аналогичную картину можно наблюдать и в онтогенезе. Так, наблюдения над эмбриональным развитием современных хвостатых амфибий показывают, что варианты развития (в частности, покровных костей черепа) у примитивных представителей группы могут стать нормой у ее продвинутых (саламандриды) представителей [566].
У всех древних палеозойских мшанок (стенолемат) обнаруживается широкий полиморфизм, проявляющийся в форме зооециев основных скелетных особей колонии, которые могут быть представлены разными морфами, авто-, эксиле-, мезо-, мета-, акантозооециями. В ходе эволюции прослеживается усложнение колоний за счет объединения числа морф в разных комбинациях (диморфные, триморфные и тетраморфные колонии). Наибольшие преобразования в эволюции претерпевают автозооеции, тогда как Другие морфы различаются в основном по числу и размерам. При этом наблюдается явное усиление полимеризации.
У палеозойских мшанок Fenestrida многообразие форм сводится к девяти структурным типам колоний (рис. 11) в момент расцвета (в карбоне), которые выводятся от четырех ордовикских типов (анастомозирующая сетка, сетка с перекладинами, перистая колония и ветвистая). Причем все девять типов формируются за счет комбинирования ряда исходных признаков путем почкования и пространственной ориентации зооидов (образование перекладин, анастомозов и т. д.). Вместе с тем в формировании этих типов прослеживаются глубокие параллелизмы в разных семействах.
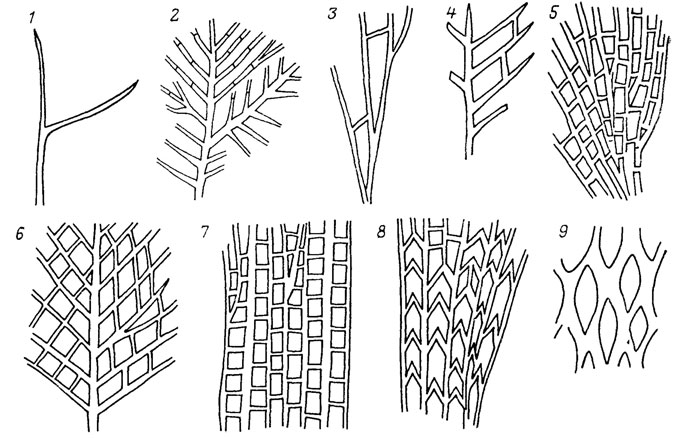
Рис. 11. Основные структурные типы колоний мшанок отряда Fenestrida. По И. П. Морозовой. 1-4 - колонии из единичных прутьев с разными типами ветвления, 5-9 - сетки, 5 - из бифуркирующих прутьев, соединенных перекладинами, 6 - с центральным утолщенным прутом, 7 - с упорядоченно расположенными утолщенными прутьями, 8 - с прямыми прутьями, соединенными сливающимися, направленными вверх перекладинами, 9 - с анастомозирующими прутьями
Идее прототипа, таким образом, противопоставляется принцип исходного морфологического многообразия, основанный на признании пластичности примитивных форм и наличии среди этой пластичности исходных вариантов всех морфофункциональных типов организации, возможных на данной структурной основе [1918]. Согласно этой концепции, формирование нового происходит в рамках конкретной морфологической организации при наличии сходных формообразовательных механизмов. Пластичность форм демонстрируется на разных системах органов ресничных червей - турбеллярий и их жизненных формах, обнаруживающих широкий спектр вариаций, на базе которых и происходит дифференцировка новых более единообразных типов. Например, упорядочение положения ротового отверстия, которое занимает первично самое разное положение (от переднего до заднего конца тела), множественности вариантов строения глотки, полового аппарата, выделительных органов, органов чувств и т. д. Правда, в отношении турбеллярий существует и противоположное мнение - о возможности их вторичного упрощения [1286]. И здесь мы сталкиваемся с примером обычной ограниченности неонтологического материала в отношении макроэволюционных выводов, поскольку из-за "горизонтального" видения (в "плоскости" современных видов) они во многом остаются гипотетичными без подтверждений на ископаемых формах.
При анализе формирования морфологических типов важно также учитывать явление, обобщенное под названием "принцип опережения" [189]. Суть его состоит в том, что внутри большого филума отдельные виды могут достигать по ряду структур состояний, свойственных вышестоящим преемственным филумам, значительно опережая, таким образом, типовые состояния своего таксономического уровня.
В результате таких, обычно единичных, морфологических сальтаций в филогенезе могут мозаично воспроизводиться состояния как относительно близких, так и далеких филетически групп. Например, среди остеолепидных кистеперых рыб некоторые виды имеют развитый задний ангулярный фланг, где, вероятно, помещался задний вентральный дивертикул спиракулярного мешка (см. рис. 9). Аналогичный фланг развит у териодонтов, топографически он сопоставим с вместилищем среднего уха млекопитающих [1033]. Среди кистеперых у палеозойского Platycephalichthys дерминтермедиальный отросток в носовой полости в силу своего прогрессивного развития смыкается с медиальной стенкой капсулы, полностью разделяя ее на верхний и нижний отделы. Подобное состояние встречено пока лишь только у одной современной бесхвостой амфибии Hypophachus [1809]. Другой пример: поразительное сходство микроструктуры зубов девонской кистеперой пандерихта и древнейшего девонского стегоцефала - ихтиостеги (или карбоновой кистеперой рыбы Pycnoctenion) - с триасовым ихтиозавром, которое, естественно, не могло быть унаследованным, а приобреталось независимо по всей видимости, на общей морфогенетической основе, сохранившейся на гистологическом уровне в филетически преемственном стволе позвоночных (кистеперые рыбы - амфибии - рептилии).
Вся эволюция кистеперых рыб, по крайней мере остеолепидных, была своего рода "генеральной репетицией" выхода на сушу, на протяжении всей их истории появлялись морфологически "стегоцефалоподобные" формы, у которых различные тетраподные черты мозаично появлялись на общем фоне рыбной организации. Примерно то же мы видим и у растений, когда черты голосеменной организации появляются в разных группах птеридофитов, а черты, свойственные покрытосеменным, - у разных голосеменных.
Таким образом, мы необязательно должны стремиться свести наблюдаемое многообразие форм к одной исходной форме. Скорее речь должна идти о другом; следует показать, как полиморфизм органа у предковой группы организмов преобразовался в то распределение признаков того же органа, которое наблюдается у потомков. Идеал "вывести разнообразие из однообразия" может быть заменен иным идеалом - "вывести разнообразие потомков из многообразия предков", учитывая при этом как постепенность, так и скачкообразность преобразований признаков и частоту их встречаемости по ходу эволюции. Если данная смена идеалов будет взята на вооружение морфологами, то это приведет к существенным изменениям во всем стиле их мышления.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'