
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
1.2. Основные особенности функциональной морфологии колониальных организмов (Е. И. Кузьмичева)
Колониальные организмы - губки, гидроиды, кораллы, мшанки; и другие - играли важную роль в составе населения дна Мирового океана в течение всего фанерозоя; преобладают они здесь над одиночными организмами и в настоящее время. Объясняется это способностью колониальных форм к росту за пределы особи, связанному с бесполым размножением. Естественно, что изучение последнего, обусловливающего возникновение колоний - своеобразных ассоциаций морфологически и физиологически связанных друг с другом особей, - наиболее актуальное направление морфофункциональных исследований колониальных организмов.
Бесполое размножение как самая первичная и древняя форма размножения, хотя и типично для организмов, находящихся на низком уровне организации, очень многообразно и выполняет ряд важных функций, таких, как регуляция размеров тела и увеличение численности вида; оно может служить также средством расселения организмов и переживания неблагоприятных условий.
Согласно О. М. Ивановой-Казас [426, 429], стимулом к началу бесполого размножения служит определенное физиологическое состояние (в разных случаях различное), возникающее под действием как внешних, так и внутренних факторов; иногда это состояние может быть охарактеризовано как общее понижение интеграции. Возникновению бесполого размножения способствуют также слабая дифференциация и полимеризация структур, которые делают возможным разделение тела на жизнеспособные части.
В биологическом отношении возникновение колониальности оказалось, несомненно, явлением прогрессивным. Колония как морфологически целое лучше, чем разобщенные формы обеспечивает питание, а следовательно, жизнеспособность и продолжительность существования вида.
Явление колониальности сложно. Его изучение ведется в различных аспектах - морфологии, формообразования, периодичности роста, индивидуального развития особей в колониях (бластогенез) и колоний как единого целого (астогенез), регенерации, полиморфизма, эволюции и т. д. В данной главе мы сосредоточим внимание на центральной проблеме - выяснения отношений между индивидуальностями отдельных особей и индивидуальностью колонии. Именно этот подход приводит к познанию биологической сущности и общей закономерности развития колониальных форм на разных уровнях их организации.
Особенно большой вклад в разработку данной проблемы внес В. Н. Беклемишев [61], установивший посредством сравнительно-морфологического анализа, что возрастание степени колониальности у губок, целентерат и других колониальных организмов идет тремя основными путями: 1) ослабления индивидуальности особей; 2) усиления индивидуальности колонии; 3) возникновения кормидиев - своеобразных осложнений колониальной системы ("колоний внутри колоний"). У разных организмов этот сложный процесс протекает неодинаково и его особенности заслуживают специального изучения.
Согласно В. Н. Беклемишеву, "...в типичных случаях колония отвечает следующим признакам: 1) колония состоит из особей, т. е. индивиды низшего порядка, из которых она образована, обладают достаточно выраженной индивидуальностью; 2) все члены колонии возникают бесполым путем за счет одной особи - основательницы; 3) все члены колонии состоят в органической связи между собой; 4) в силу этого между членами колонии в той или иной мере осуществляется прижизненный обмен веществ; 5) поведение, обмен веществ и формообразования членов колонии в той или иной мере согласованы между собой, т. е. связаны системой корреляции, направленной на поддержание существования колонии как таковой" [61, с. 75].
Колониальные формы известны на всех эволюционных уровнях, начиная с докембрийских прокариот и вплоть до хордовых животных. Если для одних организмов колониальная форма существования является постоянной, то для других временной [61]. К последним относятся жгутиковые из родов Gonium, Volvox, некоторые гидроиды, плоские и кольчатые черви. К примеру, у гидры вновь возникшие почки образуют колонию совместно с материнской особью. Существует она, однако, временно, так как, достигнув полного развития, дочерние почки отделяются от материнской особи и продолжают самостоятельное развитие. Образование таких колоний у гидр обычно рассматривают как зачаточное, но не исключено, как полагает В. Н. Беклемишев, что оно является остаточным, рудиментарным по аналогии с некоторыми лептолидами из атекат.
Колониальность, столь несвойственная для билатеральных высоко интегрированных организмов, все же имеет место среди низших их представителей, обладающих способностью к бесполому размножению (временные колонии линейного типа некоторых плоских червей и аннелид). Например, бесполая особь кольчатого червя Myrianida fasciata имеет цепочку из 29 зооидов, которые, отрываясь, превращаются в свободноплавающие половые особи. При этом важно заметить, что компонентами таких линейных колоний являются не сегменты тела, а многосегментные особи кольчатого червя.
Следовательно, временные колонии периодически распадаются на свободноплавающие особи, которые в дальнейшем могут вновь продуцировать колониальные формы.
Таким образом, существование временных колоний как бы фиксирует становление процесса колониальности в ходе эволюции и характерно для групп организмов, находящихся на низком организационном уровне.
В ископаемом состоянии временные колонии неизвестны. Встречающиеся в юрских и меловых породах "сростки" Serpula socialis есть не что иное, как скопление особей по пищевому принципу.
Изучение явления колониальности продвинуто значительно дальше на тех группах организмов, представители которых живут в настоящее время. Кроме отмеченных уже трудов В. Н. Беклемишева следует упомянуть работы Т. Вогана и Дж. Уэллса [2427, 2476], Т. Флюгеля [1588], М. А. Воронцовой и Л. Д. Лионозер [197], Б. П. Токина [1065], О. М. Ивановой - Казас [425, 426, 429], У. Оливера [2025, 2026], Т. Хамада [1698], Д. Г. Полтевой [826], Д. В. Наумова [728], Б. В. Преображенского [841, 842] и др. В течение последних 30-35 лет в связи с расширением исследований органогенных построек интерес к колониальным жизненным формам резко возрос не только в неонтологии, но и в палеонтологии, хотя специфические особенности ее объектов сильно затрудняют изучение рассматриваемого явления. Палеонтолог лишен возможности непосредственно анализировать взаимосвязи особей и вынужден судить о них, основываясь на строении и степени упорядоченности в расположении принадлежащих им ячеек скелета, а также по аналогии с современными формами. Кроме того, не все ныне живущие колониальные формы известны в ископаемом состоянии.
Согласно И. И. Шмальгаузену [1187] и Б. П. Токину [1066], всем живым существам свойственно стремление к целостности. Исходя из этого, изучение каждой колониальной формы начинается с выяснения уровня ее интеграции, определяемого соотношением образующих ее особей.
К низкоорганизованным колониальным многоклеточным организмам относят губки. Их особи, входящие в состав колоний, обнаруживают чрезвычайную пластичность, в связи с чем структура колоний довольно проста и лишена того многообразия взаимоотношения особей, которое имеет место в колониях организмов, стоящих на более высоком эволюционном уровне.
Согласно сравнительно-анатомическим исследованиям В. Н. Беклемишева [61], развитие колониальности у губок шло от примитивных, мало интегрированных колоний с хорошо обособленными зооидами (Sycon ctliatus, Leucosolenia fragihs), к колониям, где отдельные особи совершенно не обособлены. Таким образом, у последних при вхождении в колонию от каждого зооида сохраняется только устье с прилегающей к нему частью отводящей системы. Слабая интегрированность колоний губок состоит также в том, что растущие рядом колонии, соприкасаясь, могут сливаться в одну колонию. Возможен и обратный процесс, когда колония вследствие разрушительной силы волн или других механических воздействий распадается на несколько жизнеспособных колоний.
В отличие от более высокоорганизованных колониальных многоклеточных слабо выраженная индивидуальность зооидов в колониях губок не ведет к усилению индивидуальности колонии в целом. В связи с отсутствием специализации особей зооиды как бы растворяются в колониальном целом, прежде чем могли бы приобрести под его влиянием какую-либо специализацию. Этим же объяснимо и отсутствие определенной формы колоний губок, характерной для каждого вида.
Поскольку явление колониальности изучено на разных группах организмов, встречающихся в ископаемом состоянии весьма неравномерно, мы сосредоточим основное внимание на более изученных кишечнополостных, мшанках и граптолитах. Это позволит, с одной стороны, показать логическую направленность исследований, а с другой - выяснить общие закономерности эволюции.
Очень наглядно сказанное можно проиллюстрировать прежде всего на склерактиниях, известных с триаса по настоящее время, по которым имеются данные как сравнительно-анатомических исследований, так и изучения конкретных филогенезов.
Наиболее низким уровнем колониальности среди склерактиний характеризуются фацелоидные колонии, в которых кораллиты - ячейки отдельных особей - удалены друг от друга и совершенно изолированы. Следующая ступень интеграции представлена плокоидными колониями, кораллиты в которых сближены путем соединения внеячеистой тканью - перитекой. Дальнейшее увеличение уровня интеграции прослеживается в цериоидных колониях, в которых кораллиты сближены до касания друг с другом непосредственно стенками. Вместе с тем на всех трех названных стадиях интеграции кораллиты (полипы) все-таки индивидуализированы. Последующее возрастание уровня колониальности идет в сторону увеличения связей между соседними кораллитами, вплоть до слияния приуроченных к ним полипов гастроваскулярными полостями в единое целое. Таким кульминациям колониальности среди склерактиний соответствуют колонии тамнастероидного меандроидного, гиднофороидного и ценесаркового типов, в которых индивидуальность отдельных особей подчинена индивидуальности колонии как единого целого.
Такова общая направленность рассматриваемого процесса. Нельзя вместе с тем не отметить, что интеграция колониальных систем у разных групп склерактиний и даже внутри одного отряда осуществлялась разными путями (рис. 12). В качестве примера рассмотрим пути интеграции колоний в их основных филогенетических стволах [552]. Так, в подотряде Archeocoeniina, существующем с триаса поныне, усложнение колониальности шло от фацелоидных колоний к плокоидным, а далее от плокоидных, с одной стороны, к цериоидным и, с другой - к ценосарковым, т. е. к наиболее прогрессивному направлению, обеспечившему широкое распространение данных форм в современном Мировом океане.
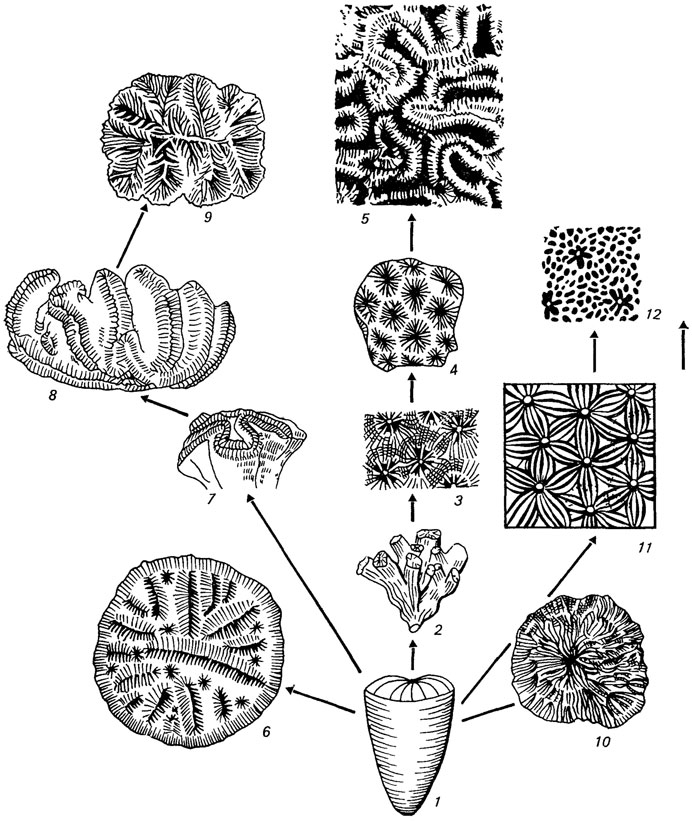
Рис. 12. Пути интеграции колоний склерактиний от одиночного организма (1) Составила Е. И. Кузьмичева. Наиболее распространенный путь - от фацелоидной колонии (2) и плокоидной (3), цериоидной (4) и меандроидной (5), более короткие пути, ведущие к высоким уровням колониальности с одной стороны, от одиночной формы (1) к гиднофороидной колонии (6), к флабеллоидной (7), от нее к флабелло-меандроидной (8) и меандроидной (9), с другой стороны - к тамнастероидным (10), (11) и ценосарковой (12)
Цериоидное направление развития на первых порах выглядит примитивным и архаичным. Но, как показали наши наблюдения на примере рода Actinastraea (юра - эоцен) из семейства Actinastraeidae, архаизм и низкий уровень колониальности оказываются тут кажущимися, так как на стыке соседних кораллитов в структуре их широких стенок различимы лакуны-каналы, которые по аналогии с современной Acropora могут указывать на наличие ценосаркальных каналов, существовавших при жизни в краевой зоне полипов, что в свою очередь указывает на колониальную целостность. В подотряде Stylinina, появившемся в триасе и вымершем в меловом периоде, этот процесс развивался от фацелоидных колоний к цериоидным и далее к меандроидным - вершине колониальности в этой группе. В подотрядах Astraeoina и Fungiina, наиболее широко представленных в мезозойских и кайнозойских рифовых фациальных комплексах, разнообразие колониальных форм и путей повышения их интеграции значительно шире. В подотряде Astraeoina (триас - ныне) нами прослежен один из упомянутых выше путей - от фацелоидных колоний к плокоидным, а затем, с одной стороны, к цериоидным, меандроидным и гиднофороидным, а с другой - к ценосарковым. Другой путь - от одиночных форм (род Montlivaltia) благодаря внутричашечному почкованию к тамнастероидному типу (род Monocyclastraea), при котором наличие мезентериальных связей между полипами соседних кораллитов приводило к высокому интеграционному уровню всей колонии.
Также разнообразно, сменяя одно усложнение другим, шло повышение уровня интеграции в подотряде Fungiina (триас - ныне), завершая развитие самыми высокими колониальными кульминациями: меандроидными, гиднофороидными, тамнастероидными и ценосарковыми формами. Кстати, развитие меандроидных форм не всегда проходило через стадию цериоидных колоний. Нередки случаи возникновения меандроидных колоний и от одиночных флабеллоидных форм.
Следует сказать, что интеграционный уровень меандроидных колоний не сохраняется совершенно постоянным, одинаково выраженным морфологически. У форм, характеризующихся неполным слиянием кораллитов в ложбины, таких, как оксфордские, Myriophyllia rastellina Mich, и M. angustata d'Orb., он, несомненно ниже, чем у форм, отличающихся завершенностью процесса формирования ложбин. Многое в этом отношении зависит от высоты стенок-гребней между ложбинами и степени развития на них амбулакров. Чем гребни ниже, а амбулакры развиты сильнее, тем выше уровень колониальности, так как здесь налицо уже возникновение ценосарковой ткани.
Ценосарковый тип колонии достиг большого разнообразия среди кайнозойских, в том числе ныне живущих склерактиний. Существование у этих форм общеколониальных связей между особями, свидетельствующее о наиболее высокой степени их интеграции, было экспериментально доказано Н. Н. Марфениным [624] на двух видах рода Acropora из Австралийского Большого барьерного рифа. Ему удалось обнаружить в ценосарке этих склерактиний канальную распределительную систему, регулярно снабжающую колонию жидкостью. В этой системе Н. Н. Марфенин выделил четыре морфофункциональные зоны: верхушки ветвей, сами ветви, стволы и основание колонии, характеризующиеся различным распределением движущихся токов жидкости и репродуктивными способностями полипов. Канально-распределительная система ценосарка включает морфологически обособленные полости в той части скелета, которая непосредственно граничит с мягким телом: аксиальный канал, циркумаксимальные полости, полости радиальных кораллитов и интеррадиальные полости. Подобная морфологическая и функциональная дифференциация колоний Acropora, несомненно, свидетельствует не об условной их целостности, как полагали ранее, а о реальной и высокой физиологической интеграции.
В этой связи есть основания полагать, что многие роды Fungiina и в первую очередь обладающие пористым скелетом также имели подобную соединительную систему.
Вместе с тем в других группах склерактиний наблюдается иной ход развития колониальности. Так, в сравнительно молодом подотряде Eupsammiina, появившемся в начале мелового периода и существующем ныне, первые колониальные формы, относящиеся к данию, так и остались на низком уровне интеграции, соответствующем фацелоидному и субплокоидному типам. В датском веке возникли также первые фацелоидные колонии (Faksephyllia) в другом подотряде Caryophylliina, что служит примером синхронного параллелизма в морфогенезе названных склерактиний. Синхронный параллелизм известен и в развитии более древних склерактиний. Он выразился, в частности, в одновременном появлении в среднетриасовую эпоху колониальных форм сразу в четырех подотрядах: Archaeocaeniina, Stylinina, Astraeoina и Fungiina. Наряду с синхронным нередко давал о себе знать и гетерохронный параллелизм. Одним из его примеров служит возникновение первых колониальных форм среди Meandriina в поздней юре, у Fungiina, например, в семействе Acrosmiliidae (p. Bacsanophyllia) - в валанжинском веке, а в семействе Cunnolitidae (p. Aspidastraea) - в туронском веке.
Процесс развития интеграции у колониальных склерактиний шел параллельно с процессом полимеризации, причем также разными способами, хотя приводил к одинаковой биологической сущности - лучшей адаптационной способности в облове пространства щупальцами, что в свою очередь обусловливало совершенствование специализации самих особей на путях создания целостной системы. Например, в подотряде Archaeocaeniina упорядочение в расположении кораллитов на ветвях колоний шло от спирального у эоценового рода Stylophora к расположению рядами у неогенового рода Seriatopora (рис. 13, а); в подотряде Stylinina - от беспорядочного у позднеюрского рода Tiaredendron к перистой системе у позднеюрского - раннемелового рода Enallohelia. В подотряде же Astraeoina процессы полимеризации и упорядочения кораллитов идут параллельно. У позднемелового рода Diblasus ветви колоний немногочисленные и толстые, а кораллиты расположены на них беспорядочно; у эоценового рода Diplohelia ветвление колонии более частое, в расположении кораллитов намечаются ряды; наконец, у рода Neohelia (миоцен - ныне) ветви колоний тонкие и на них отчетливо видны ряды кораллитов (см. рис. 13, б, в).
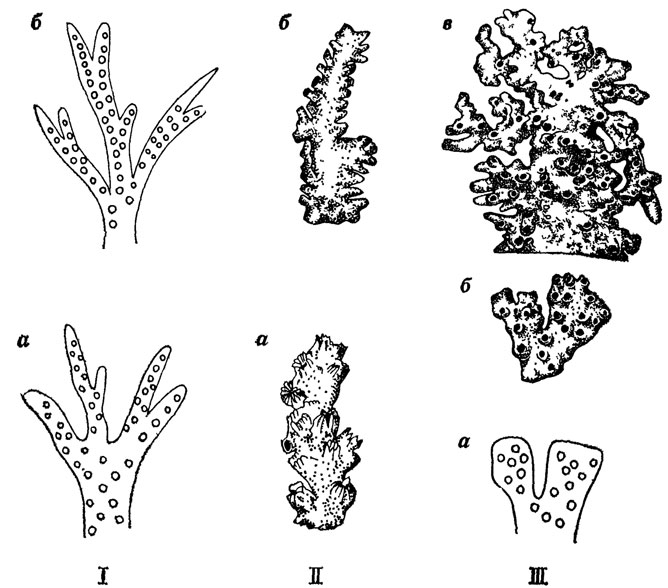
Рис. 13. Пути полимеризации и упорядочения расположения кораллитов на ветвях колоний склерактиний. По Е. И. Кузьмичевой. I - в сем. Pocillopondae: спиральное (а) у рода Stylopora (эоцен - ныне), к расположению рядами (б) у рода Seriatopora (миоцен - ныне), II - в сем. Stylinidae: a - беспорядочное у рода Тtaradendron (верхняя юра), к перистому (б) у рода Enalloheha (верхняя юра - нижний мел), III - в сем. Oculinidae: беспорядочное (а) у рода Diblasus (верхний мел) к расположению рядами (б) у Diplohelia (эоцен) и далее к большей полимеризации (в) у рода Neohelia (миоцен - ныне)
Таким образом, колониальность есть следствие генетически запрограммированного процесса бесполого размножения, идущего по определенной схеме (или схемам). В отличие от таких колоний, которые следует называть истинными, существуют еще колонии ложные, возникающие незапрограммированно, случайно. Как следствие этого почкующиеся особи ложной колонии развиваются неупорядоченно. Примером ложных колоний могут служить часто встречающиеся в нижнем берриасе Горного Крыма экземпляры разных видов одиночных кораллов из рода Montlivaltia. У них наблюдается развитие небольшого количества (обычно две-три) дочерних почек, спровоцированное внедрением червей-трубкожилов. Ложные колонии монтливалтий обычно трудно отличимы от встречающихся в этих же слоях истинных колоний Monocyclastraea alpina (Koby). В астогенезе последних дочерние почки, как правило, в количестве восьми - девяти возникали внутричашечным путем последовательно по спирали [549].
Наиболее часто образование ложных колоний происходит вследствие травматических повреждений и внедрения организмов-комменсалов с последующей регенерацией скелета. Последняя представляет собой частный случай регуляции - одного из наиболее общих свойств организма и состоит в его способности отвечать на разнообразные нарушения путем их устранения и возвращения к прежнему состоянию, либо приспособления к новым условиям.
На развитие колониальности в подклассе ругоз (ордовик - нижний триас) также влиял процесс интеграции от фацелоидных колоний к цериоидным, затем астероидным, тамнастероидным и афроидным [1578, 2019, 2026]. Меандроидные формы представлены лишь одним раннепермским родом Wentzelella. Таким образом, тамнастероидный и афроидный типы колоний представляют собой у основной массы ругоз самые высокие уровни интеграции.
Обычно колонии ругоз мономорфны. Однако М. Минато и К. Роветт [1962] первыми обратили внимание на диморфизм колоний ругоз, выраженный в различии строения протокораллита, в котором находилась особь - продукт полового размножения, и других кораллитов, продуцированных в ходе бесполого размножения. Несколько позднее И. Федоровски [1578] и Л. М. Улитина [1076] стали различать в некоторых колониях ругоз явление полиморфизма. Наряду с обычными кораллитами, которые с ростом достигают зрелости и сами образуют дочерние кораллиты, у них прослеживаются более мелкие недоразвившиеся кораллиты. Последние, отпочковавшись от материнского организма, остаются незрелыми до конца своего роста. Такие кораллиты И. Федоровски [1578] назвал "потерянными структурами" (lost structure). Согласно наблюдениям Л. М. Улитиной [1076], незрелые кораллиты формировались на различных стадиях астогенеза. В массивных колониях Evenkiella cincta Ul. они появлялись в основном на неастенической стадии, а у ветвистой Minussuella beljakovi Bul. - только на эфебастической стадии.
Недавно у цериоидных колоний ругоз из рода Lonsdaleia обнаружена своеобразная система каналов [2258], служившая, по-видимому, для сообщения гастральных полостей полипов. Эти трубчатые каналы (диаметром от 0,25 до 0,8 мм и длиной около 11 мм) располагаются в диссепиментариуме наклонно к оси родительского кораллита в местах ответвления от него почек. Иногда каналы соседних кораллитов сливаются друг с другом с образованием щелей.
Сравнительно более низким интеграционным уровнем колоний среди кораллов характеризуется подкласс Tabulata (кембрий - пермь), в котором известны фацелоидные, цериоидные и катениформные формы. Последние с точки зрения расположения кораллитов занимают промежуточное положение по отношению к первым двум типам, хотя, принимая во внимание морфологическую диморфность кораллитов, стоят, по-видимому, выше в функциональном отношении [2026]. Повышение интеграционного уровня фацелоидных и цериоидных колоний табулят шло путем совершенствования коммуникативных связей и увеличения количества обладавших ими форм [2026; рис. 6 на с. 15]. Так, у фацелоидных колоний из отряда Syringoporida существовала активная, хотя и временно функционировавшая связь соседних кораллитов посредством трубок. В составе другого отряда Favositida известны некоммуникатные и коммуникатные колонии, причем в развитии последних прослеживается постепенный переход пор с ребер на грани кораллитов, способствовавший усилению физиологической общности располагавшихся в них полипов.
Несколько особняком среди рассмотренных групп кораллов стоят гелиолитоидеи (ордовик - девон), характеризующиеся высоким уровнем колониальности, связанным с развитием колоний полиморфного типа. Замечено [96, 99, 101, 102, 789], что морфологическое разнообразие гелиолитоидеи в силуре по сравнению с ордовиком обусловлено появлением и развитием гетероморфных особей: диморфных колоний (генезисный полиморфизм), три- и тетраморфных колоний (генезисный и, вероятно, функциональный полиморфизм). Полиморфные колонии возникали независимо в разных ветвях гелиолитоидей, причем триморфные колонии произошли от диморфных в результате превращения кораллитов в качественно новые компоненты; тетраморфные колонии развивались из триморфных в результате модификаций цистолитов и сифонолитов. Кроме того, у тетраморфных колоний наблюдается упорядоченность в расположении компонентов, приводящая к возникновению образований, аналогичных кормидиям современных книдарий.
Насколько можно судить по ныне живущим формам, высокой степенью колониальности обладают восьмилучевые кораллы, достоверные остатки которых известны с мела по настоящее время. Наличие ценосарковой системы каналов, диморфизм особей у некоторых представителей и развитая нервная система, способная к передаче раздражений от одного участка колоний к другому, ставят эту группу кораллов на ступень выше в колониальной интеграции по сравнению со склерактиниями. Подтверждением сказанного служит упорядоченность расположения диморфных полипов в некоторых колониях альционаций. В частности, в грибовидной колонии Anthomastus автозооиды - неспециализированные полипы, выполняющие функции питания, - располагаются в краевой части "шляпки" колонии, тогда как сифонозооиды - специализированные полипы, выполняющие в основном функцию "насосов", снабжающих колонию жидкостью, - сосредоточены в центре ее. В развитии же семейства Helioporidae (баррем - ныне), обладающего наружным скелетом с четким диморфизмом ячеек, намечается тенденция к полимеризации, состоящая в переходе в меловом периоде от мелких полусферических колоний к преимущественно рамозным и кустистым колониям, существующим и ныне. Диморфизм же полипов в колониях гелиопорид не обнаружен. Мелкие трубчатые образования в их ценостеуме отвечают не специализированным полипам, а пальцевидным выростам нижней части тела ценосарка. К сожалению, фрагментарность остатков внутреннего скелета большинства других ископаемых октокораллов (горгонид, пеннатуллид) не позволяет достаточно точно прослеживать эволюцию в степени их колониальности.
Совершенно неосвещенным в палеонтологической литературе остается вопрос о развитии колониальности у ископаемых гидроидных, хотя современные их представители хорошо изучены с этой точки зрения [61, 473, 623, 728]. Установлено, что колонии гидроидов из подотряда Tecaphora представляют собой целостную систему, хотя физиологическая основа ее интеграции нуждается в дальнейшем уточнении. Известно также, что интеграция колоний у гидроидов осуществляется благодаря перемещениям гидроплазмы - жидкости, заполняющей полость тела. В колониях этих организмов один и тот же клеточный материал часто используется для поочередного формирования разных их частей. Клетки рассасывающихся старых гидрантов используются для образования новых; за счет прекращения роста одних ветвей гидроризы при неблагоприятных условиях может поддерживаться рост других и т. д. Известно также, что клетки могут мигрировать с одного конца колонии в другой. Сказанное относится и к пище, получаемой одним гидрантом и переходящей затем к другим. Все это убедительно свидетельствует, что колония гидроидов не есть условное целое, состоящее из комбинации однотипных "блоков" как полагали ранее, а представляет собой реально целое, где индивидуальность отдельных особей подчинена индивидуальности колонии.
Особое положение среди гидроидных занимают гидрокораллы, характеризующиеся редукцией медузоидного поколения и развитием известкового скелета. На специализацию особей в колониях указывает функциональный диморфизм зооидов. Вершиной колониальности в классе Hydrozoa, несомненно, являются сифонофоры, у которых функциональная специализация особей привела к образованию целостной системы, стоящей на грани интегрированной колонии и целого организма с самостоятельными органами. Согласно Л. А. Зенкевичу [407], подвижный образ жизни явился для этих животных основным стимулом к высокой организации тела.
После книдарий мшанки являются наиболее продвинутыми организмами в плане высокой интеграции колоний, где отдельные особи несут различные специализированные функции. В целом развитие колониальности во всех трех классах этих организмов шло в прогрессивном направлении, состоящем в последовательном усилении структурной интеграции от мономорфных колоний (зоарий) к колониям с высоко развитым полиморфизмом [712, 713].
Процесс колониальной интеграции мшанок шел разными путями. Так, у пресноводных, преимущественно современных форм из класса Phylactolaemata, лишенных полиморфизма, развитие шло по пути слияния цистидов (вместилищ для полипидов) и формирования ценесарка.
Примером предельно высокой интеграции колоний этого класса может служить род Cristatella - ползающая мшанка со слившимися цистидами и с хорошо развитой общеколониальной локомоторной мускулатурой.
У мшанок из класса Gymnolamata (ордовик - ныне), отличающихся от филактолемат обызвествлением стенок цистидов, особенностями строения зооидов и значительным упрощением строения полипида, колониальная интеграция шла по пути усиления полиморфизма и усложнения типов почкования, обусловливающих разные способы расположения зооидов в колонии (зоарий). Эволюция цистида протекала здесь от примитивно трубчатой к компактно-укороченной форме с параллельным усложнением структуры стенок.
В целом полиморфизм мшанок можно рассматривать как важное биологическое приспособление, возникшее еще в ордовике, а различные пути и формы его проявления - как основное звено эволюционного процесса в этой группе организмов.
Остановимся несколько подробнее на формировании гетерозооециев, а следовательно, и полиморфизма в историческом развитии на примере трех основных филогенетических стволов мшанок, рассматриваемых в ранге подотрядов одного из крупных отрядов Trepostomata, существовавшего с раннего ордовика по триас. Большая роль в изучении этого вопроса принадлежит Г. Г. Астровой [34]. У самых примитивных мшанок, относящихся к подотряду Esthonioporoidea (ордовик - девон), на фоне еще полностью мономорфных колоний с автозооидами уже появляются зачатки видоизмененных особей - акантозооециев, выполняющих, по-видимому, защитную функцию. Вторая ветвь трепостомат - подотряд Halloporoidea (ордовик - карбон) - отделилась от эстониопороидей Развитие колониальности и полиморфизма в этой группе мшанок шло по пути возрастания ветвистых форм колоний и усиления роли гетерозооециев. Так, у самого древнего семейства этого подотряда Dittoporoidae впервые появившиеся метозооеции (видоизмененные особи, сидящие в трубках с большим числом диафрагм) еще не выходили на поверхность колоний (зоарий), они закрывались отложением известковой ткани, а присутствие акантозооециев было весьма неопределенным и неустойчивым. Зато в двух других семействах - Heterotripidae и Trematoporidae (ордовик - карбон) - мезозооеции и акантозооеции развиты наиболее полно. Третья ветвь трепостамат - подотряд Amplexoporoidea (ордовик - пермь) - в своем развитии утрачивает мезозооеции полностью, но зато впервые появляется новый тип гетерозооециев - эксилязооеции (правда, в ордовике они еще редки, но уже в девонских зоариях достигают полного развития). Наибольшего расцвета эксилязооеции достигли в каменноугольный и пермский периоды у амплексопороидей.
Высокой колониальной целостности достигли формы из отряда Cheilostomida, характеризующиеся большой пластичностью полиморфных зооидов, что, по-видимому, и обеспечило им процветание в Мировом океане. Среди них наибольшую ступень колониальности представляют крибриморфные мшанки (надсемейство Cribrilinidacea), обладающие не только коммуникационными порами, снабжающими пищей непитающиеся самостоятельно индивиды, но и высоким полиморфизмом [1865]. Последний развит настолько, что каждый автозооид с окружающими его авикуляриями, порядком камер, овицеллами и измененными оральными шипами можно рассматривать как "субколониальную единицу" В. Н. Беклемишева [61].
Колонии многих палеозойских мшанок также организованы в дискретные единицы - субколонии, возникающие от монтикул [714]. Их рост, таким образом, есть результат роста субколоний. Рост же последних саморегулируется, с одной стороны, путем физиологического контроля над темпами почкования в монтикулах, с другой - влиянием условной среды. Монтикулы представляют собой полюс морфогенетического поля, от которого к границам субколонии происходит последовательная смена морфогенетических градиентов, влияющих на размеры, форму и плотность расположения зооидов.
На указанных примерах полиморфизм мшанок не ограничен в своем развитии. У некоторых форм он достигает онтогенетического полиморфизма с развитием на разных стадиях жизненного цикла одного зооида различных функций, вплоть до репродуктивной [1278].
Как и у книдарий, процесс интеграции колонии мшанок шел параллельно с упорядочением расположения зоэций на ветвях. Результатом этого явились вертикально-спиральные типы колоний, весьма экономные с точки зрения площади субстрата и эффективно функционирующие при захвате пищи щупальцами. Этот тип обеспечивает и механическую прочность колоний путем вторичного многослойного обрастания зоария за счет почкования экзозональных зооидов, дистальные концы которых находятся ближе к основанию зоария, чем проксимальные [2366].
Вызывает интерес также явление полиэмбрионии, прослеженное на колониях современных и некоторых пермских мшанок [1941]: слияние соприкасающихся колоний одинакового диаметра (колонии с различным диаметром этого эффекта не дают). Надо полагать, что гистологическая совместимость возможна только у полиэмбрионических колоний, развивающихся из личинок одного генотипа.
В целом, несмотря на регрессивные признаки мшанок, связанные с сидячим образом жизни (редукцию движений, нервной, выделительной и кровеносной систем), развитие данного типа шло по пути биологического прогресса. Регрессивные признаки компенсировались развитием высокой колониальной целостности, следствием чего явилось длительное существование этой группы организмов.
Хотя граптолиты относятся к давно вымершим организмам, они изучены с морфофункциональной точки зрения довольно детально Сказанное относится прежде всего к филогенетически связанным друг с другом отрядам Dendroidea из подклассов Stereostolonata и Graptoloidea, различающимся степенью интегрированности колоний. У большинства представителей дендроидей, которые вели прикрепленный образ жизни, почкование шло упорядоченно путем образования очередных триад. Наличие у них трех типов тек, рассматривавшееся первоначально как проявление функционального полиморфизма особей колоний, в настоящее время связывают с тем, что столотеки не были заняты особым типом зооидов, а соответствовали юным зооидам. Таким образом, текальный триморфизм, в сущности, соответствовал диморфизму особей, связанному с половым различием [1845]. В отличие от дендроидей, у граптолоидей, которые вели, вероятно, планктонный или дрейфующий образ жизни, наблюдается элиминация битек и соединение столотек с автотеками, что обусловило возникновение мономорфных тек и, следовательно, гермафродитизм зооидов.
Для всех граптолоидей свойственна правильная морфологическая последовательность в строении тек. Согласно А. Урбанеку [2410], у самых примитивных граптолитов она проявляется в постепенном, закономерном увеличении тек в дистальном направлении и в незначительных изменениях их пропорций. Теки проявляют регулярный морфологический градиент (градиент величины тек) вдоль длины рабдосомы. Важной чертой в организации колоний уже примитивных граптолоидей является полярное (гетерополярное) строение. Подчеркнуть это следует и потому, что у их предков - дендроидей - теки не обнаруживают никаких изменений, зависящих от их положения вдоль длины рабдосомы. Даже у Anisograptidae - группы, занимающей промежуточное положение между Dendroidea и Graptoloidea, никаких признаков полярной организации обнаружено не было. Несомненно, что полярность в организации колонии знаменует высокий уровень ее организации и достижение ее явилось важным событием в эволюции Graptoloidea.
Биологически осмысливая рассматриваемое явление, А. Урбанек установил [2404, 2406-2408], что у наиболее простых одноосевых колоний монограптидов можно различать: 1) проксимально-прогрессивный тип, где проксимальные теки отличаются от дистальных присутствием определенных признаков или их более сильным выражением; 2) дистально-прогрессивный тип, в котором, напротив, наиболее сложное строение приобретают именно дистальные теки. Наличие двух типов полярной организации имеет большое значение для понимания способов эволюции колоний граптолитов.
Обобщенное представление о развитии граптолитов дает морфофизиологическая теория организации их колоний, разработанная А. Урбанеком [2404, 2405]. В основу ее положено предположение, что морфологический градиент возникал у граптолитов на фоне физиологического градиента. Проявление последнего связано, по-видимому, с выработкой и постепенным уменьшением количества или концентрации определенных веществ, распространявшихся вдоль рабдосомы. Гетерополярная организация колоний граптолоидей наталкивает на мысль, что эти морфологически активные субстанции возникали на одном из их полюсов и затем продвигались к другому, проявляя определенный градиент. Вероятно, указанные вещества вырабатывались сикулозооидом и распространялись от него в ткани бластозооидов. Следовательно, сикулозооид составлял центр, физиологически доминирующий над остальными особями колонии. Подтверждение этому А. Урбанек видит в регенерационных морфозах, изученных им на примере поврежденных ветвей Linograptus [2405]. Ветви колоний сложены двумя сериями тек, растущих в противоположные стороны. Одна из них представляет собой фрагмент первичной рабдосомы с оторванной сикулярной частью. Вторая серия, возникшая в результате регенерации, развилась при отсутствии индукционного центра (сикулозооида) и сформирована однообразными теками, напоминающими теки дистальной части первичной рабдосомы.
В течение всего жизненного интервала граптолитов, со среднего кембрия до среднего девона включительно, в развитии колониальности этих организмов прослеживаются две тенденции.
1. Явно интеграционный процесс, связанный с переходом от бентосного к планктонному образу жизни. Способность к вертикальной миграции в слоях богатой пищей фотической зоны во многом благоприятствовала приобретению граптолитами полярной организации колоний и связанных с этим двух способов их роста:
а) продолжением немы или виргулы и почкования зооидов;
б) перехода от двухрядности к однорядности ячеек в колонии.
2. Процесс ослабления колониальности, вплоть до ее исчезновения, прослеживающийся у среднедевонских форм и связанный, вероятно, с вымиранием группы. Среди разнообразных причин, обусловивших исчезновение граптолитов, немаловажную роль играло, по-видимому, сокращение пищевых ресурсов в планктонной биомассе, нашедшее отражение в уменьшении содержания в породах углефицированного органического материала.
Если в ходе эволюции ряда групп граптолитов имело место повышение степени интеграции колоний, сменившееся на стадии вымирания ее снижением, то у других организмов, включая табуляты, ругозы и вымершие таксоны склерактиний, аналогичные морфогенетические преобразования не обнаружены. Это заслуживает дальнейших наблюдений и осмысления.
Образование колоний известно не только у многих беспозвоночных, но и у примитивных позвоночных, относящихся к подтипу Tunicata (оболочников). Появление у них бесполого размножения - наиболее древнего способа размножения в истории животного мира, согласно О. М. Ивановой-Казас [427], - связано, несомненно, с сидячим образом жизни. П. Бриан [1378] заметил, что из всех многоклеточных животных именно у туникат наблюдается наибольшее разнообразие форм бесполого размножения: сосудистое (почки возникают на сосудах туники); паллеальное (почкование путем выпячивания наружной стенки туники); пилорическое (формирование бластозооида из двух почек, закладывающихся порознь); столониальное (почкование стелящегося основания - столона); стробиляция (поперечное отшнуровывание части материнского организма). В ископаемом состоянии остатки туникат представлены только спикулами, изучение которых ничего не дает для суждений о колониальности этих организмов. Поэтому мы ограничиваемся ссылками на работы, посвященные их рецентным формам.
Колонии как формы существования живых организмов, способных к бесполому размножению, представляли собой явление более прогрессивное, чем одиночные формы. И это преимущество проявлялось прежде всего в быстром и экономном росте популяций и в более эффективном противостоянии неблагоприятным условиям существования.
Стабильность колониальности на разных эволюционных уровнях была различной. Менее стабильно это явление у низкоорганизованных форм, имеющих, например, временные колонии, и более стабильно - у высокоорганизованных организмов.
Изучение явления колониальности имеет большое теоретическое и практическое значение. Оно обеспечивает лучшее понимание природы и закономерностей развития колониальных организмов, среди которых многие группы имеют важное породообразующее, биостратиграфическое и биологическое значение. Очевидно, что палеонтология может внести существенный вклад в выяснение путей формирования и общей направленности эволюции колониальных жизненных форм в истории жизни на Земле.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'