
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
1.3. Морфофункциональный анализ спирально свернутой раковины головоногих моллюсков (И. С. Барсков)
Спиральная форма - наиболее распространенный способ реализации морфологии природных объектов (в живой природе в особенности). Универсальность спирального строения, характеризующего широкий круг объектов и процессов от молекул до галактик, общеизвестна. Такая универсальность, несомненно, является отражением общих закономерностей развития и самоорганизации материи на разных уровнях, что свидетельствует об огромной "поливалентности" спирального строения. Эта универсальность может трактоваться как неспецифичность спиральной морфологии, способной обеспечить широкий спектр функций и потому функционально необусловленной.
Применительно к биологическим объектам функциональная неспецифичность спирального строения как будто бы должна вызывать сомнения в возможности адаптивного объяснения такой морфологии. И действительно, правильность спирального строения раковины головоногих моллюсков, свойственная весьма различным таксонам, приводилась в качестве доказательства не только неадаптивности морфологии раковины, но и неадаптивного характера самого эволюционного процесса [308].
Головоногие моллюски в целом используют в строении раковины практически все возможные геометрические варианты плоской логарифмической спирали независимо от предполагающихся различий в образе жизни и от внутренней морфологии раковины. Можно привести многие примеры тождественности наружной морфологии раковин, принадлежащих заведомо различным таксонам головоногих моллюсков [1173].
Более того, тождественные геометрические варианты раковины встречаются, хотя и с разной частотой, среди головоногих и фораминифер. Отсюда следуют два заключения.
1. Тождественность спирального строения не имеет приспособительного значения.
2. Тождественность спирального строения является отражением того, что именно таким способом (простейшим в энергетическом и геометрическом симметрийном смысле) обеспечивались разные адаптивные потребности. Спиральная морфология действительно наиболее древний и экономичный способ морфогенеза.
Таким образом, вопрос состоит не в том, адаптивна или нет сама спиральная морфология как таковая (она может обслуживать разные функции), но адаптивны или нет те различия в геометрии спирали, которые могут быть зафиксированы среди форм, у которых спиральная раковина выполняет принципиально одинаковую функцию. Следует найти соответствие частоты тех или других вариаций степени, интенсивности, успешности выполнения этой функции и таким образом адаптивно объяснить эти вариации.
Как показал анализ выполненный на массовом материале, включавшем около 80% всех известных родов цефалопод с плоскоспиральной раковиной, такая попытка имеет смысл и дает определенные биологически значимые результаты.
Раковина у цефалопод исходно представляла собой коническую прямую или согнутую трубку, в широкой части которой (в жилой камере) располагается мягкое тело, а задняя часть (фрагмокон) выполняет функцию гидростатистического аппарата. Приобретение планоспиральной свернутости само по себе имеет отчетливый адаптивный смысл. Это дает возможность расположить центры тяжести и плавучести на одной вертикали, или даже в одной точке (у Nautilus несовпадение этих центров составляет около 2 мм). Это дает возможность животному постоянно находиться в состоянии безразличного равновесия, что обеспечивает минимум усилий для приобретения любого необходимого положения в воде. Прямая или согнутая раковина без дополнительных механизмов регулирования плавучести обусловливает лишь гипостомное положение в воде.
Цефалоподы со свернутой раковиной, впервые появившиеся в раннем ордовике (отряд Tarphycerida), в течение долгого времени не были многочисленными и лишь с девона (с появлением отряда Nautilida и аммоноидей) получили преобладающее распространение. Этот тип раковины возник независимо по меньшей мере в трех самостоятельных крупных филогенетических ветвях.
Уже первые исследователи геометрии спирально свернутых раковин [2398] указывали, что в подавляющем большинстве случаев она может быть описана логарифмической спиралью. Основными характеристиками геометрической формы раковины являются параметры спирали и форма поперечного сечения. Количественное описание формы может быть ограничено этими характеристиками. Их числовые параметры выражаются формулой логарифмической спирали или, что постоянно применяется при традиционных описаниях, в виде так называемых стандартных измерений (рис. 14, 1).
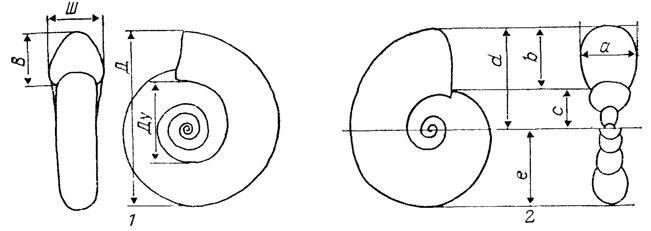
Рис. 14. Схемы измерений свернутых раковин цефалопод. 1 - схема стандартных 'измерений': В - высота оборота, Ш - ширина оборота, Д - диаметр раковины, Ду - диаметр умбилика; 2 - схема измерений, по Д. Раупу: S= b/a - форма поперечного сечения; D=c/d - форма образующей кривой (степень объемлемости); W = (d)2e - скорость расширения оборотов
В то же время рассмотренный способ представления данных затрудняет сравнение различных форм, ибо исследователь оперирует только числами.
Д. Рауп предложил оптимальную систему измерений и сравнения количественных описаний геометрии спиральных раковин [2123, 2124]. Три параметра, рассчитанные по пяти простым измерениям (см. рис. 14, 2), которые легко можно сделать по изображениям раковин (и даже более точно, чем по реальным образцам), дают достаточно полное представление о геометрической форме всех классов спиральных раковин. И. А. Ванчуров показал полную математическую адекватность всех указанных выше численных методов характеристики формы плоскоспиральной раковины [139]. Методика Д. Раупа позволяет моделировать форму с помощью аналоговой ЭВМ и наглядно сравнивать результаты измерений, нанесенные на скаттердиаграммы.
Уже первая попытка такого сравнения, проведенная на анализе распределения форм раковин 405 родов аммоноидей [2124], показала специфичность "аммонитовой" области среди всех возможных форм свернутых раковин и поставила ряд интересных проблем. Основные выводы Д. Раупа сводятся к следующему.
1. Раковины аммонитов по скорости расширения оборотов D и степени объемлемости оборотов W занимают довольно обширную область (W 1,2-3,5; D 0-0,65), которая, однако, почти не выходит за линию, отмечающую формы с несоприкасающимися оборотами: W=1/D (рис. 15).
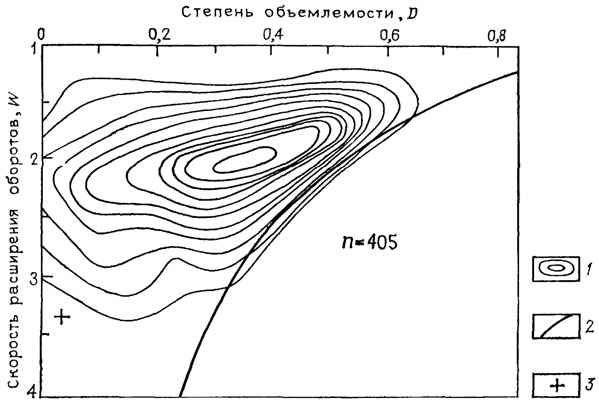
Рис. 15. Схема распределения по форме раковин аммоноидей. По Д. Раупу. 1 - изолинии плотности точек, полученных в результате измерения голотипов типовых видов или экземпляров другого вида данного рода, 2 - линия, ограничивающая поле раковин с несоприкасающимися (развернутыми) оборотами (справа), 3 - положение раковины современного Nautilus pompilius. Здесь и далее до рис. 25 n - число измеренных раковин
2. Общее распределение измеренных раковин внутри выделенной области одномодально: вокруг одного пика группируется максимальное количество родов аммоноидей.
3. Отдельные таксоны (это показано для Goniatitida и Litoceratina) распределены внутри общеаммонитовой области различно, группируясь вокруг разных морфологических пиков.
4. Эти различия связаны, вероятно, с различиями в образе жизни и местообитании.
5. Современный Nautilus выпадает по форме раковины из "аммонитовой" области. Другие Nautiloidea не изучались "и остается только гадать, случайно ли это различие, или же оно отражает тот факт, что Nautilus, а может быть, и все наутилоидеи в ходе эволюции приспособились к другому адаптивному пику" [2124].
Эти выводы явились лишь постановкой ряда вопросов. Анализ более обширного материала позволяет дать более определенные ответы на некоторые из них и провести возможную адаптивную интерпретацию особенностей основных геометрических типов свернутой раковины. Было измерено и проанализировано более 2070 экземпляров спиральных раковин, принадлежащих к 1200 видам 787 родов, 9 отрядов цефалопод. Установлены четыре наиболее "популярных" морфологических типа свернутых раковин, вокруг которых группировалось наибольшее число родов в разных таксонах цефалопод. В разные геологические эпохи среди определенных групп преобладали представители разных морфотипов.
Ключом к выделению основных типов послужило распределение наутилоидеи (отряды Tarphycerida и Nautilida, рис. 16). В отличие от схемы "общеаммонитового" распределения Д. Раупа распределение наутилоидеи (100 родов) оказалось более равномерным. На фоне этой равномерности выделились четыре морфологических пика, четыре области сгущения точек, соответствующих определенным параметрам формы раковины: морфологическая область А (модальные значения: D~0,5; W<2) - малобъемлющие медленно расширяющиеся раковины: морфологическая область Б (D 0,2-0,3; №~2,2) - среднеобъемлющие раковины с умеренно расширяющимися оборотами; морфологическая область В (D~0,1; W~2) - сильнообъемлющие раковины с медленно расширяющимися оборотами; морфологическая область Г (D<0,1; W>3,3) - сильнообъемлющие раковины с быстро расширяющимися оборотами.
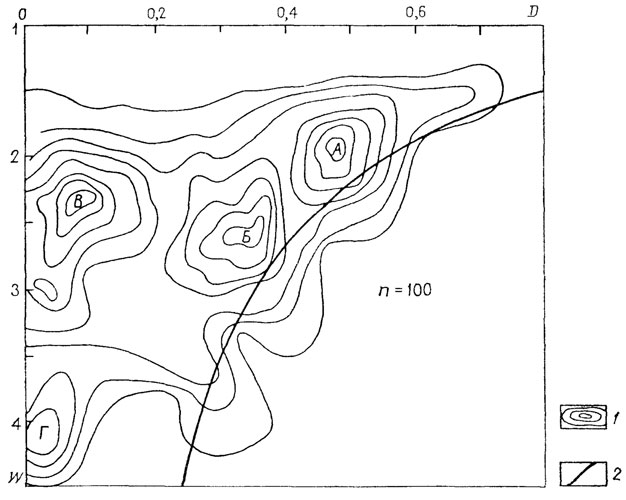
Рис. 16. Схема распределения по форме плоскоспиральных раковин Nautiloidea (Tarphycerida и Nautilida). Отчетливо видны четыре морфологические области, отвечающие выделенным морфогруппам раковин - Л, Б, В, Г; 1 - изолинии плотности точек; 2 - линия, ограничивающая поле раковин с несоприкасающимися (развернутыми) оборотами справа
Весьма показательно изменение распределения формы раковины наутилоидеи во времени (рис. 17). Раннепалеозойские Tarphycerida практически не заходили в область сильнообъемлющих (инволютных) раковин, наибольшее число родов было сосредоточено в области А.
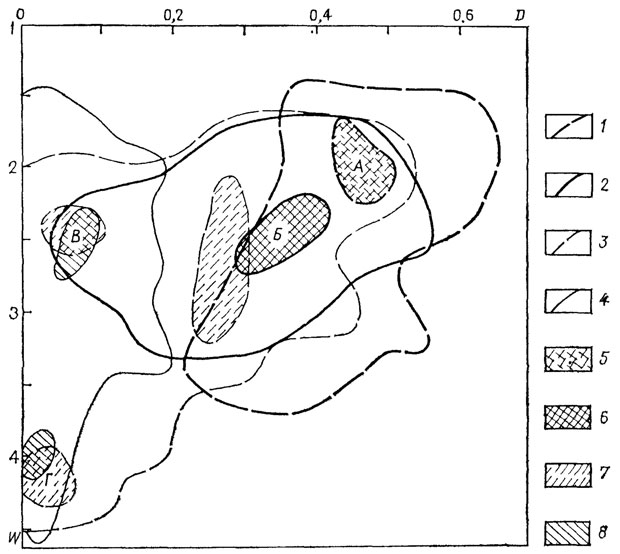
Рис. 17. Схема распределения по форме раковин Nautiloidea во времени. 1-4 - морфологические области: 1 - ордовикских и силурийских Tarphycerida, 2 - каменноугольных Nautilida, 3 - триасовых Nautilida, 4 - меловых и кайнозойских Nautilida; 5-8 - морфологические пики форм: 5 - ордовикских и силурийских, 6 - каменноугольных, 7 - триасовых, 8 - меловых и кайнозойских, Л, Б, В, Г - морфологические группы свернутых раковин
Каменноугольные Nautilida обладали большим количеством представителей с объемлющими оборотами: область морфологического пика отчетливо сдвинута в сторону более инволютных и быстрее расширяющихся раковин. Пик располагается в морфологической области Б вблизи "общеаммонитового" (и "общецефалоподового") пика. Среди триасовых наутилид были представители с разнообразной раковиной, причем появились морфологические типы, характерные для областей В и Г. Эти два типа строения свернутых цефалопод - единственные среди меловых и кайнозойских наутилид.
По данным П. Уарда [2454, фиг. 3], те же морфологические области занимали и юрские наутилиды. Современный Nautilus по форме раковины располагается в области Г. Полученные данные с полной определенностью показывают, что эволюция формы раковины среди наутилоидеи проходила по пути приобретения более инволютной и быстро расширяющейся раковины. После позднетриасового кризиса в развитии отряда Nautilida среднеинволютные и эволютные формы среди них вообще не возникали. Форма раковины современного Nautilus - не случайное уклонение, а результат длительного эволюционного процесса.
Основываясь на общих соображениях и экспериментальных данных Р. А. Реймента [2147], полученных на моделях раковин, обладающих разной скоростью расширения оборотов (рис. 18), можно сделать заключения о функциональном значении отмеченных тенденций. Увеличение скорости расширения оборотов уменьшает потенциальную плавучесть раковины, т. е. для достижения нейтральной плавучести быстрорасширяющимся формам (Nautilus в приведенном примере) требовалось приложить меньше усилий, чем медленно расширяющимся. Увеличение степени инволютности оборотов (уменьшение расстояния образующей спирали от центра свертывания) способствовало сближению центра тяжести и центра плавучести, что облегчало достижение безразличного равновесия, упрощало управление плавучестью и ориентированным положением в воде. Этими качествами обладают представители морфологических областей В и Г, различаясь скоростью расширения оборотов.
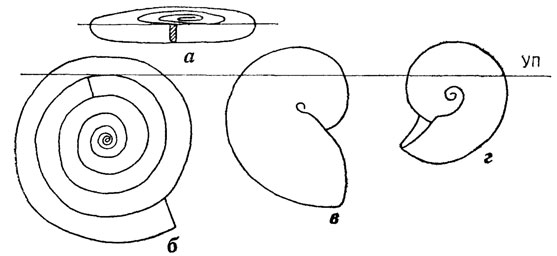
Рис. 18. Плавучесть и устойчивость спиральных раковин с различной скоростью расширения оборотов и степенью их объемлемости. По Р. Л. Рейменту. а, б - медленно расширяющиеся эволютные раковины (Dactylioceras): а - очень высокая плавучесть, полная неустойчивость положения, б - положение в воде такой же раковины, если размер жилой камеры более 1,5 оборотов, низкая плавучесть и относительная устойчивость; в - быстро расширяющаяся инволютная раковина (Nautilus): максимальная устойчивость, плавучесть малая, но больше, чем в случае б, г - умеренно расширяющаяся, среднеинволютная раковина (Ceratites), максимальная плавучесть, умеренная устойчивость; УП - уровень погружения раковины в воду при пустых камерах фрагмокона и заполненной водой жилой камере
Формы области Г, к которым принадлежит современный Nautilus, исходя из теоретических соображений, должны быть придонными обитателями (низкая плавучесть), обладающими хорошей маневренностью и способностью принимать любое положение в воде (легкость достижения безразличного равновесия), не способными, судя по форме поперечного сечения раковины, к длительному активному плаванию, но с вероятной возможностью к кратковременным быстрым перемещениям (гладкая обтекаемая раковина) с малой инерционностью. Эта теоретическая модель хорошо согласуется с данными о бентопелагическом образе жизни Nautilus, полученными в последние годы. Вероятно, представители морфологической области Г принадлежали к той же адаптивной зоне.
Исходя из общих соображений, наиболее выигрышной, казалось бы, должна быть форма раковины из морфологической области В. Эти представители сочетают высокую потенциальную плавучесть медленно расширяющихся форм и преимущество в достижении безразличного равновесия сильнообъемлющих форм. Они были распространены среди палеозойских аммоноидей, но редки среди наутилоидей (триасовые Gonionautilus, Gryponautilus, Callaionautilus). Однако такая форма раковины неизбежно влекла за собой очень сильное изменение формы мягкого тела животного, в частности мантийной полости. Из более или менее цилиндрического (как у форм из области А) или конического (как у форм из областей Б и Г) мягкое тело должно было приобрести сплющенную до листовидной и латерально изогнутую до серповидной в сечении форму. Ясно, что уменьшение объема и усложнение формы мантийной полости приводило к резкому ослаблению органа активного движения - воронки, к ограничению подвижности. Пожалуй, единственный возможный образ жизни для представителей морфологической группы В - планктонный, а большой запас плавучести и простота достижения безразличного равновесия заставляют предполагать их эпипланктонное существование.
В значительной степени сходными адаптивными характеристиками должны обладать медленно расширяющиеся эволютные формы, которые типифицируются морфологическим пиком в области А. Как показано в экспериментах Р. А. Реймента [2147] на примере Dactylioceras, такие формы при длине жилой камеры до 1 оборота имеют настолько высокую плавучесть и настолько малую устойчивость, что при отсутствии механизмов, утяжеляющих раковину, занимают почти горизонтальное положение на поверхности воды. Для достижения равновесного вертикального положения в воде для них необходимо было иметь жилую камеру, занятую мягким телом с плотностью, близкой к плотности воды, не менее чем в 1,5 оборота. Такие формы реально существовали.
Теоретически необязательно иметь жилую камеру указанной длины. Камера может быть короче, но тогда необходимо, чтобы последние камеры фрагмокона были бы заполнены жидкостью, так чтобы суммарно объем утяжеленной части раковины составлял не менее полутора оборотов. При этом оказывается, что вся система "мягкое тело + заполненные камеры фрагмокона + "пустые" камеры" имеет плавучесть, меньшую чем Nautilus - представитель морфологической области Г (см. рис. 18). Таким образом, основными гидростатическими характеристиками раковины представителей области А являются малая устойчивость, высокая потенциальная плавучесть (но при относительно устойчивом положении реальна низкая плавучесть), наличие длинного субцилиндрического до червеобразного мягкого тела, форма которого диктуется геометрией раковины.
Последний признак в адаптивной интерпретации указывает на то, что у таких животных мантийная полость не могла обеспечить сильного пропульсивного импульса, что исключает возможности быстрого плавания [1996] и заставляет считать их принадлежащими планктону. Высокая потенциальная плавучесть позволяет предполагать, что они могли совершать значительные вертикальные миграции. Вероятно, близким адаптивным аналогом этих форм является современная Spirula со сходным геометрическим типом раковины. Хотя раковина спикулы внутренняя, это с функциональной точки зрения не имеет значения.
Представители со спиральной раковиной из области Б ("общеаммонитовый" морфологический пик) обладают во всех отношениях средними гидростатическими и гидродинамическими характеристиками, которые в принципе обеспечивают максимальные экологическую пластичность и адаптивную широту. Поэтому неслучайно именно здесь располагается "общеаммонитовый" (и "общецефалоподовый") морфологический пик. Эта морфологическая область была наиболее популярной. В разные геологические эпохи такой морфологией обладали представители различных систематических групп. В некоторых же группах вообще не было форм этого типа.
Можно предположить следующую адаптивную интерпретацию особенностей строения раковин из морфологической области Б. Сравнительно короткая жилая камера и умеренно перекрывающиеся обороты свидетельствуют о функционально развитой мантийной полости и, следовательно, о способности животного к передвижению с помощью воронки, существенно более активному, чем позволяло строение раковин областей А и В. Таким образом, для них необязателен планктонный образ жизни. Тафономические данные о преимущественно раздельном захоронении наутилоидей с раковиной типа Г и аммонитов с раковиной типа Б (как в палеозое, так и в особенности в мезозое) указывают на их обитание в существенно различных адаптивных зонах. Это не позволяет относить аммонитов к бентопелагической жизненной форме. Исключая представителей с раковиной морфологического типа Б из планктонной и бентопелагической жизненных форм и учитывая их способность к более активному способу передвижения, мы должны прийти к заключению о их принадлежности к нектонной или нектобентосной адаптивной зоне.
В самом определении нектона как адаптивной жизненной формы существуют различные аспекты. Вопрос об отнесении тех или иных конкретных представителей к нектону зависит от того, каким из критериев придавать основное значение. В качестве определяющих критериев можно предположить пелагическое существование и способность к активному плаванию как возможность активно преодолевать расстояния на какой-то порядок больше, чем длина собственного тела, как абсолютная скорость передвижения, как независимость собственного движения от течений, как активный поиск пищи, а также способность размножаться в толще воды. Совершенно очевидно, что все эти критерии в значительной степени относительны и взаимодополняемы.
Морфологические параметры, выраженные в коэффициентах обтекаемости, определяемых экспериментально [1421], и числа Рейнольдса не позволяют точно определять нектонные формы даже среди ныне живущих нераковинных форм, хотя они и позволяют выявить потенциальную "степень" нектонизации. "Нектонные" морфологические параметры могут оставаться и нереализованными в действительном образе жизни животного. Наглядным примером этого могут служить данные Ю. А. Алеева [11]. По гидродинамическим показателям (характер поперечного сечения, обтекаемость, числа Рейнольдса) отмечается значительное перекрытие полей, характеризующих нектонные и планктонные формы. По своим параметрам цефалоподы со свернутой наружной раковиной, несомненно, попадут, судя по экспериментам с моделями, в это обширное поле перекрытия.
Приведенные выше соображения о степени развития аппарата активного движения и форме мягкого тела, обусловленные морфологией представителей типов А и В, позволяют отделить планктонные формы, даже если по гидродинамическим параметрам они попадают в поле перекрытия, но разделение нектонной и нектобентосной жизненных форм представляется весьма сложным. Достоверно этого можно достигнуть лишь прямым наблюдением за образом жизни, что невозможно.
Существуют представления о том, что нектонные цефалоподы появились вообще геологически поздно, на протяжении позднего кайнозоя, что к собственно нектерам можно относить лишь кальмаров открытого океана [742]. Вполне вероятно, что среди древних цефалопод со свернутой наружной раковиной не было аналогов этой нектонной группы. Это, однако, не означает, что из нектона нужно исключать современных неритических кальмаров и те формы ископаемых, наружная свернутая раковина которых обладает параметрами, максимально удовлетворяющими по соотношениям с другими типами свернутых раковин гидродинамическим требованиям нектонной жизненной формы. При этом, конечно, сохраняется относительность выводов (что, впрочем, характерно для большинства морфофункциональных и экологических заключений, сделанных на палеонтологическом материале). Она обусловлена как неполнотой палеонтологической летописи (сама же полнота относительна, и в связи с применением новых методов исследований может существенно изменяться), так и невозможностью прямого наблюдения.
В качестве особенностей раковины морфологического типа Б, свидетельствующих о большей нектонизации, можно рассматривать следующие. Форма поперечного профиля раковины: дисковидные со сжатым дорсовентрально сечением раковины рассматриваются потенциально более нектонизированными; наличие скульптурных особенностей, способствующих устойчивому положению раковины при активном движении, также считается приспособлением к нектонному образу жизни. В качестве таковых можно рассматривать присутствие нескольких килей на наружной стороне оборота, уплощенность или вдавленность края и вообще наличие продольных скульптурных элементов. Сильно уплощенные дисковидные формы с острым килем на наружной стороне, по-видимому, обладали скорее хорошей маневренностью, чем высокой скоростью движения, на что указывал Б. П. Марковский [622], хотя сечение раковины и допускает последнее.
Остаются слабо исследованными в функциональном и адаптивном отношении особенности поперечной скульптуры, столь разнообразно развитой у раковин морфологического типа Б. Рассматривать их просто как украшения, не контролируемые адаптивно, по-видимому, нельзя. Само присутствие поперечной скульптуры жестко скоррелировано со степенью инволютности раковины. Твердо установлено, что при инволютности D<0,2 поперечно скульптированные формы возникали как редкое исключение. Среди крайне эволютных форм (морфологическая область А) более 80% представителей несут поперечную скульптуру. Исключение среди них составляют литоцератиды, однако многие из них имеют сильна развитые воротники, игравшие явно стабилизирующую функцию, что однозначно решает вопрос о планктонном образе жизни этих представителей. Учитывая малую устойчивость раковин морфологического типа А, более вероятно, что основная функция поперечной скульптуры - повышение устойчивого положения в толще воды. Некоторое значение, по-видимому, небольшое, поперечная скульптура могла иметь и для повышения прочности раковины, особенно если принять положение о значительных вертикальных миграциях, свойственных этим формам.
Для представителей морфологического типа Б поперечная скульптура точно так же должна была иметь функцию сохранения устойчивого положения животного, но не в статическом, как в предыдущем случае, а в динамическом (из-за несомненно большей активности) режиме. Поперечные ребра могли выполнять роль своеобразных стабилизаторов при движении.
Нельзя также не отметить одной особенности в развитии поперечных ребер, отличающих свернутые раковины аммоноидей и наутилоидей. У наутилоидей поперечные ребра при переходе через наружную сторону оборота изгибаются выпуклостью назад, к начальной камере, т. е. следуют в своих очертаниях линиям нарастания и вырезу гипономического синуса на устьевом крае. У аммоноидей ребра переходят через наружную сторону оборота либо прямо поперечно, либо с отчетливым, иногда значительным изгибом выпуклостью к жилой камере. Ни по сохраняющимся очертаниям устьевого края, ни по линиям нарастания у аммоноидей не отмечается вентрального выреза устья, соответствующего гипономическому синусу.
Кажется несомненным, что эти различия в морфологии отражают весьма существенные различия в строении воронки - аппарата активного движения. У современного наутилуса воронка представляет собой мускулистое образование в виде листка, свернутого конусом, с несросшимися краями. Такое строение позволяет менять направление и силу реактивной струи. Подвижность воронки дает возможность животному изменять направление движения, вплоть до плавания головой вперед. Возможно, что наличие гипономического синуса на устьевом крае раковины отражает подвижность воронки. Вместе с тем двулепестковое строение, несомненно, ограничивает устойчивое "тяговое" усилие этого реактивного движителя. Современный наутилус способен развить скорость в 25 см/с лишь на протяжении 2-3 с.
Устойчивое и равномерное движение активно плавающих современных внутреннераковинных цефалопод обеспечивается воронкой, представляющей собой сплошную коническую трубку, не способную изгибаться назад и сообщить животному "попятное" движение. Правда, у некоторых кальмаров имеются внутри воронки специальные приспособления, позволяющие осуществлять резкое торможение за счет изменения направления выбрасываемой из воронки струи.
Поскольку в настоящее время мнение о близости аммоноидей и внутреннераковинных по строению мягкого тела общепризнанно, есть основания полагать, что строение воронки у них было сходным. Вероятно, воронка у аммоноидей обеспечивала преимущественно поступательное движение, а маневрирование осуществлялось, как и у внутреннераковинных, с помощью щупалец-рук. Высказанные соображения вполне объясняют отсутствие гипономического синуса на раковинах аммоноидей и особенности их поперечной скульптуры.
Таким образом, можно сделать вывод, что выделенные четыре основные морфологические группы свернутых раковин, которые типифицируются определенными модальными значениями параметров геометрической формы, по Д. Раупу (для группы А: D~0,5, W<2; для группы Б: D~0,2-0,3; W~2,2; для группы В: D~0,1, W~2; для группы Г: D<0,1, W>3,3), обладают существенно различными гидростатическими и гидродинамическими качествами, отражающими обитание их хозяев в адаптивных зонах моря и свидетельствующими о их принадлежности к разным жизненным формам.
Представители морфологических групп А и В могут рассматриваться как планктонные; группы Б - как нектонные (неритические) и нектобентосные; группы Г - как бентопелагические жизненные формы. Эти оценки являются относительными, между морфологическими группами (как, впрочем, и между адаптивными типами, которые они отражают) имеются переходы. Однако прослеживание этих переходов и изменений в геометрической форме раковины на протяжении исторического развития групп позволяет вынести определенные суждения об адаптивных сдвигах в их эволюции и о взаимных конкурентных отношениях. Выше были кратко рассмотрены эти изменения у наутилоидей. Далее они будут показаны для основных групп палеозойских и мезозойских аммонитов.
Отряд Agoniatetida. Распределение по скорости расширения оборотов и степени объемлемости раковины 66 родов этого отряда резко неравномерное (рис. 19). Выделяются две отчетливые морфологические области: эволютных, медленно расширяющихся, и инволютных раковин. Среди последних, в отличие от наутилоидей (см. рис. 16), области медленно расширяющихся (группа В) и быстрорасширяющихся раковин (группа Г) не разделены: имеется только один пик, занимающий промежуточное положение (D~0,02-0,1; W~2,5-3); эти формы можно классифицировать как инволютные умереннорасширяющиеся. К ним принадлежало большинство родов надсемейства Medlicottiacea. В адаптивной интерпретации их можно классифицировать как нектонные или нектобентосные формы и переходные к бентопелагическим. Отдельные крупные таксоны среди агониатитов характеризуются специфичной формой раковины. Наиболее разнообразен подотряд Cephroceratina, для него невозможно указать наиболее "типичную" форму раковины. Для подотряда Anarcestina характерны большие вариации степени объемлемости оборотов (D~0,5-0,01), но в целом малая скорость расширения оборотов (W<2). Представители подотряда принадлежали в основном планктону.
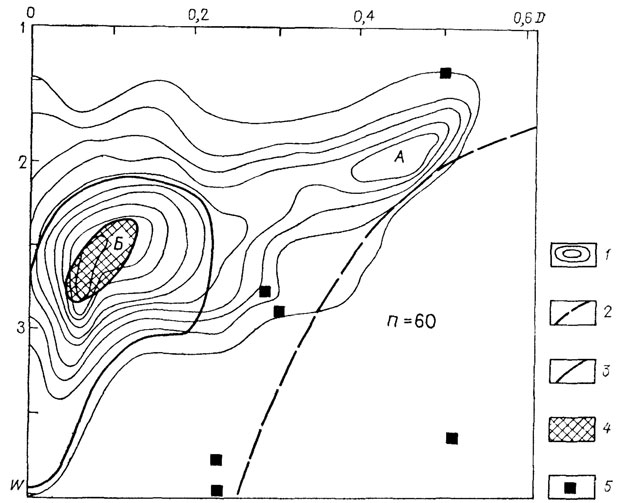
Рис. 19. Схема распределения по форме раковин Agoniatitida. 1 - изолинии плотности точек; 2 - линия, ограничивающая поле раковин с несоприкасающимися (развернутыми) оборотами (справа); 3 - граница морфологической области надсемейства Medlicottiaceae; 4 - морфологический пик Medlicottiaceae; 5 - положение родов надсемейства Mimogomatitacea, исходного для отряда Agoniatitida. Для агониатитов в целом характерны два морфологических пика (морфогруппы А и Б), для медликоттий - один (морфогруппа Б)
Среди Prolecanitina выделяются две морфологически и адаптивно раздельные группы. Роды надсемейств Medlicottiacea и Sageceratacea занимали отмеченную выше промежуточную морфологическую область, тогда как представители Prolecanitacea располагались в нетипичной для всего отряда области (D~0,2-0,3; W~2,5; морфологическая область Б).
Как отмечалось выше, представители этой морфологической области обладают наибольшими экологическими потенциями и поэтому не случайно, что у происходящих от них морфологически специализированных Medlicottiacea и Sageceratacea прослеживается и адаптивная специализация в сторону бентопелагических форм. В составе Prolecanitacea имеются два рода с эволютной раковиной, что в адаптивном плане указывает на их планктонизацию. Прослеживание изменений формы раковины агониатитов во времени показывает, что предковое для отряда надсемейство Mimoceratacea характеризуется чрезвычайно большим разбросом скорости расширения оборотов. Все их представители располагаются вблизи линии, отделяющей свернутые формы от форм с несоприкасающимися оборотами. Ранне- и среднедевонские агониатиты быстро освоили практически все характерные для цефалопод типы свернутой раковины. Морфологическая область позднедевонских представителей отряда заметно сужается; каменноугольные агониатиты занимали еще более ограниченную область с преобладанием почти инволютных (D менее 0,15) умеренно расширяющихся (W = 2,2-2,7) форм. Область морфологических типов раковины позднейших - пермских и триасовых представителей - располагается целиком в области инволютных раковин со скоростью расширения, близкой к таковой Nautilus. Таким образом, в историческом развитии агониатитов преобладало то же направление в сторону доминирования инволютных быстрорасширяющихся форм, которое было свойственно и наутилоидеям, проделавшим этот путь на несколько миллионов лет раньше.
Отряд Goniatitida. Распределение преобладающей формы раковины у представителей этого отряда (рис. 20) имеет целый ряд существенных отличий. Отмечается сдвиг преобладающих значений скорости расширения оборотов в сторону меньших значений, т. е. к преобладанию медленно расширяющихся раковин (более потенциально плавучих и адаптивно более планктонизированных форм). Основной "водораздел" на схеме изолинии плотности проходит по значениям весьма малой скорости расширения (W<2) в сравнении с агонититами (W~2,5). Менее 15% родов имеют скорость расширения большую, чем W~2,5, и лишь у трех эти значения превосходят W = 3.
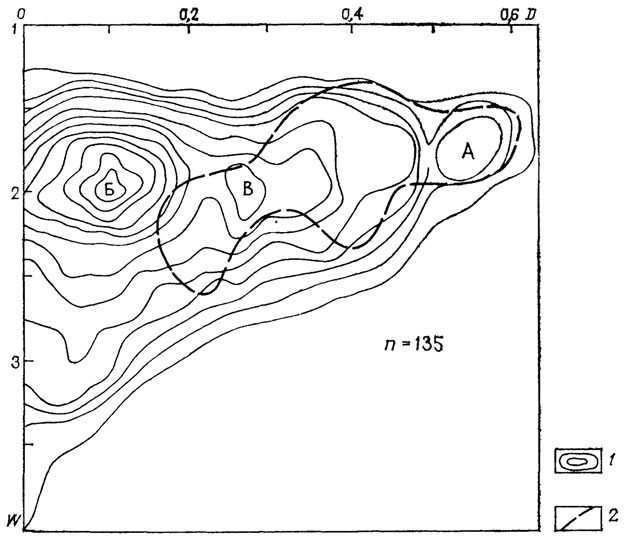
Рис. 20. Схема распределения по форме раковин Goniatitida. 1 - изолинии плотности точек; 2 - морфологическая область надсемейства Gastrlocerataсеае. Выделяются три морфологических пика (морфогруппы А, Б, В). Представители надсемейства Gastrtocerataceae входили в морфогруппы А и Б
Выделяются три морфологических пика, отвечающие областям А, Б и В. В отличие от агониатитов и наутилоидей существенно большее число родов занимает морфологическую область Б, хотя она сильно сдвинута в сторону форм с меньшей скоростью расширения оборотов, как и морфологическая область В инволютных раковин, в которой располагалось большинство представителей всех крупных таксонов отряда Agoniatitida. Исключение составляет лишь надсемейство Gastrioceratacea, в котором практически: отсутствуют инволютные формы, тогда как морфологическая область А - максимально эволютных форм занята почти исключительно представителями этого надсемейства.
В историческом развитии гониатитов проявлялись тенденции изменения преобладающей формы раковины, прямо противоположные тем, которые отмечены для агониатитов и наутилоидей. Если у последних со временем морфологическое разнообразие сужалось и все большее развитие получали инволютные формы (морфологические группы В и Г), то у гониатитов разнообразие геометрических типов форм возрастало и появлялись более эволютные и медленно расширяющиеся раковины. Поскольку различия в форме раковины отражают различия в экологии и местообитаниях, то, вероятно, экологическое разнообразие гониатитов в процессе эволюции возрастало. Однако это отражало скорее не занятие крупных: адаптивных зон, а приспособление к сравнительно ограниченным экологическим нишам в пределах планктонной адаптивной зоны.
Таким образом, представители наутилоидей, агониатитов и гониатитов обладали в целом раковинами одной и той же геометрической формы. Чрезвычайное морфологическое сходство между отдельными представителями этих крупных групп известно уже давно. Интересно сравнить распределение формы раковин у представителей, существовавших одновременно. На схеме (рис. 21) отчетливо видно несовпадение областей наиболее распространенной формы раковины, свойственной названным таксонам. Следовательно, представители наутилид, агониатитов и гониатитов в каменноугольное время обладали существенно различной морфологией раковины и обитали часто в разных адаптивных зонах [52]. Гониатиты составляли подавляющее большинство в планктоне; наутилоидей были преимущественно нектобентосными; агониатиты, обладавшие более инволютными раковинами, принадлежали частично к нектону и частично, вероятно, переходили к бентопелагическому образу жизни.
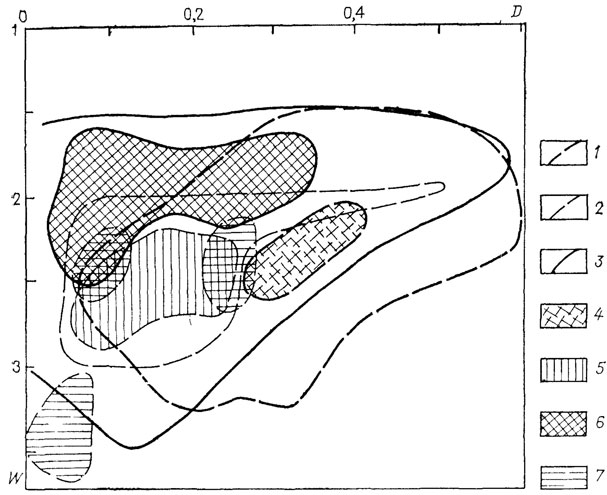
Рис. 21. Схема распределения по форме раковин одновременно существовавших каменноугольных представителей отрядов Nautilida, Agoniatitida и Goniatitida. 1-3 - морфологические области: 1 - Nautilida, 2 - Goniatitida, 3 - Agoniatitida; 4-7 - морфологические пики: 4 - Nautilida, 5 - Goniatitida 6 - Agoniatitida, 7 - триасовых Nautilida
На той же схеме показаны области, которые заняли наутилиды в конце палеозоя и в триасовом периоде. Можно полагать, что морфологический сдвиг наутилид в область инволютных и быстрорасширяющихся раковин обусловлен вымиранием занимавших эту область аммоноидей и адаптацией к новым условиям в освобождавшейся зоне. Возможно также, что здесь они оказались более конкурентоспособными и вытеснили аммоноидей. В триасовом периоде область, отвечающая морфологическому пику наутилид, была занята "новыми" аммонитами - цератитами. Таким образом, наутилиды в триасе оказались под тройным конкурентным давлением: цератиты вытесняли их из нектобентосной и нектонной адаптивной зоны; в планктоне преобладали гониатиты и цератиты; бентопелагическая зона была занята агониатитами. Не этим ли объясняются необычные морфологические "эксперименты" триасовых наутилид (возникновение у них сложно складчатой перегородки - Clydonautilidae, Siberionautilidae, антицератитовой перегородочной линии - Yakutionautilus [28]) и, наконец, почти полное вымирание в конце триаса (известный позднетриасовый кризис, когда длительное время существовал лишь один род наутилид)? Наутилиды выжили, окончательно перейдя к бентопелагическому существованию, в адаптивную зону, которую они занимают по настоящее время, выдержав здесь конкуренцию со стороны юрских и триасовых аммоноидей. Некоторые попытки адаптивной радиации предпринимались наутилидами в меловое время, когда они были существенно разнообразнее.
В развитии мезозойских аммоноидей не наблюдается таких резких морфологических сдвигов, как у палеозойских представителей данного класса. Схема распределения формы раковины в отряде Ceratitida значительно отличается от рассмотренных выше. Хотя общее разнообразие морфологических типов цератитов то же самое, однако наибольшее число родов сосредоточено в морфологической области Б (D~0,3-0,4; W~2,25). Подобного единого морфологического пика не имел ни один из рассмотренных выше отрядов.
Как уже указывалось, такая морфология наружной раковины характеризуется некоторым "средним", оптимальным соотношением гидростатических и гидродинамических качеств и наибольшей адаптивной широтой. По-видимому, именно за счет этого в сочетании с некоторыми, пока неясными преимуществами в строении мягкого тела и способе управления плавучестью, которые выражались в известном усложнении лопастной линии, позволили этому отряду вытеснить ранее существовавших примитивных аммоноидей, что, возможно, привело к их вымиранию. Однако и сами цератиты уступили пришедшим им на смену новым группам юрских и меловых аммоноидей. В морфологическом и, по-видимому, экологическом смысле последние буквально пришли "на их место".
Общая схема распределения родов юрских и меловых аммоноидей по морфологическим параметрам практически совпадает с цератитовой схемой (рис. 22) с тем же единым морфологическим пиком, приходящимся на область "оптимального" строения. Однако в отличие от цератитов среди них проявилась отчетливая морфологическая дифференциация трех групп отрядного ранга. Если подавляющее большинство представителей отряда Ammonitida оставалось в области морфологического пика, то Phyllocerida занимали совершенно отличную область, совпадающую с той, которая была характерна для гониатитов в девоне и для агониатитов - в перми. Морфологически они были совершенно обособлены от одновременно существовавших Lytocerida.
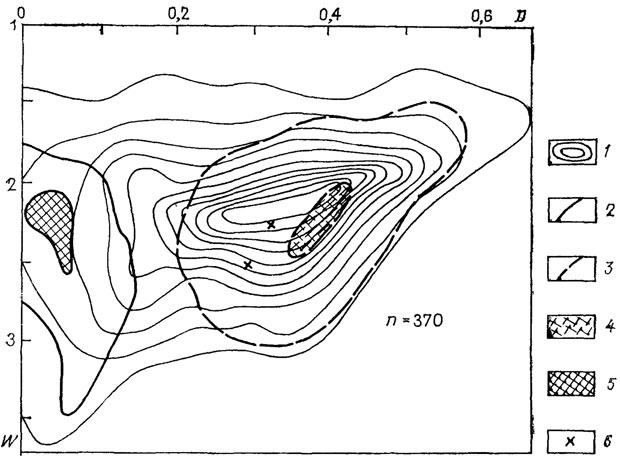
Рис. 22. Схема распределения по форме раковин мезозойских аммоноидей. 1 - изолинии плотности точек; 2, 3 - морфологические области отрядов: 2 - Pbylloterida, 3 - Lytocerida; 4, 5 - морфологические пики: 4 - Lytocerida, 5 - Phylloeerida: 6 - представители Phylloeerida, не попавшие в морфологическую область этого отряда
Морфологическая область и пик Lytocerida сдвинут в область эволютных раковин почти вплотную к линии, отделяющей формы с несоприкасающимися оборотами. В адаптивной интерпретации такое распределение раковины означает следующее: большинства литоцерид тяготело к планктонной адаптивной зоне, филлоцератид - к нектонной и бентопелагической, аммонитиды принадлежали преимущественно нектобентосу.
Среди наиболее многочисленного в юре и в мелу отряда аммонитид, несомненно, существовали морфологически и экологически разнообразные формы. Тем не менее даже при самом общем рассмотрении заметно весьма существенное изменение преобладающей формы раковины от ранней юры к позднему мелу. В ранней юре при значительном разнообразии преобладали более медленно расширяющиеся и эволютные формы (D~0,4-0,5; W~2), к мелу прослеживается сдвиг общего морфологического пика в сторону среднеинволютных умеренно расширяющихся раковин (D~0,3; W~2,5). Такая тенденция в изменении преобладающей формы раковины характерна для эволюции всех рассмотренных крупных таксонов, за исключением гониатитов.
Рассмотренные схемы, на которых каждый род представлен одной, в значительной мере случайной точкой (ибо нет никакой гарантии и в том, что типовой вид рода обладает "типичной" для рода морфологией), несомненно, дают представление о степени распространенности определенного морфологического типа в составе крупного таксона. Такой подход позволяет интерпретировать основные морфологические группы свернутых раковин как наиболее вероятные адаптивные типы, показать, что возникновение и развитие таксонов высшего ранга связано с проникновением в новую адаптивную зону, что сами эти таксоны могут рассматриваться не только как систематические, но и как экологические группировки. В ряде случаев на этом основании удается проследить основные черты экологической эволюции групп, их взаимные конкурентные отношения. Тем не менее все это позволило пока выяснить только самую общую картину.
Предлагаемый морфофункциональный и как его следствие экологические подходы могут, по-видимому, оказаться полезными и на более низком систематическом уровне, вплоть до видового. С их помощью можно подойти к определению видов как биологической реальности в функциональном и экологическом аспектах (вид как обитатель единичной экологической ниши, сколько видов- столько экологических ниш).
Возможность и правомерность таких исследований может быть проиллюстрирована результатами изучения по той же методике некоторых произвольно выбранных таксонов в пределах отряда Ammonitida. Единственным мотивом для выбора материала была возможность проведения измерений на достаточно большом количестве родов в составе семейства, подродов и видов в составе рода, а также экземпляров одного вида.
Поле морфологического разнообразия семейства Simbirskitidae (рис. 23) достаточно велико, положение морфологического пика обусловлено главным образом представителями двух из трех родов семейства, тогда как представители рода Craspedodiscus весьма сильно отличаются морфологически от двух других родов. Следующие схемы (рис. 24) иллюстрируют специфичность морфологии шести подродов рода Aulacostephanus. На приводимой схеме (см. рис. 24) показано распределение морфологических особенностей для трех из них. Несмотря на некоторое перекрытие, морфологические области для каждого из этих подродов существенно различаются. Согласно Б. Циглеру [2517], они развивались по независимым линиям от подродов рода Rasenia: Aulacostephanoides - Aulacostephanus и Aulacostephanites - Aulacostephanoceras. В первой линии (см. рис. 24) заметно ясное изменение в морфологии, выражающееся в приобретении более инволютной раковины. Во второй линии (рис. 25) при переходе от Aulacostephanites к Aulacostephanoceras несколько увеличивается морфологическое разнообразие, но морфологический пик остается на том же месте, несколько увеличивается число видов с более эволютной раковиной. Аналогичную картину, доказывающую специфичность морфологической области на видовом уровне, иллюстрирует схема для видов рода Deshaesites из другого надсемейства аммонитид.
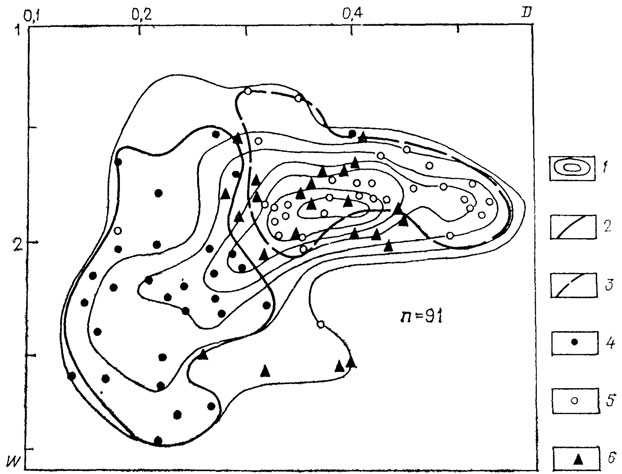
Рис. 23. Схема распределения по форме раковин родов семейства Simbirskitidae (Perisphinctaceae, Ammonitida). 1 - изолинии плотности точек; 2, 3 - морфологические поля родов: 2 - Craspedodiscus; 3 - Speetoniceras; 4 - род Craspedodiscus (7 видов, 29 экз.); 5 - род Speetoniceras (12 видов, 22 экз.); 6 - род Simbirskites (5 видов, 30 экз.)
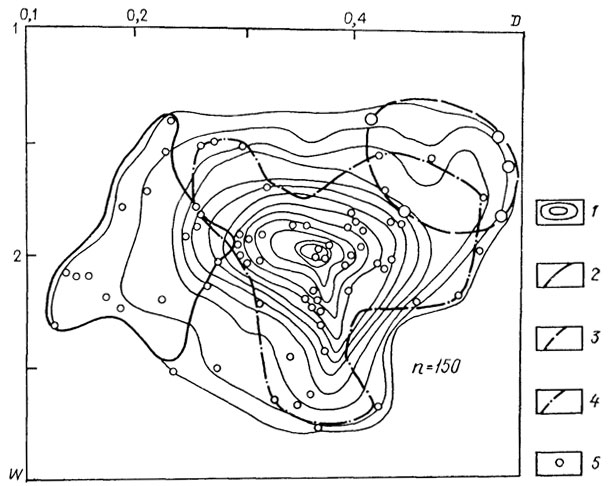
Рис. 24. Схема распределения по форме раковин подродов рода Aulacostephanus. 1 - изолинии плотности около 150 точек, принадлежащих 34 видам 6 подродов; морфологические поля подродов; 2 - Aulacostephanus; 3 - Xenostephanus; 4 - Aulacostephanoceras; 5 - точки результатов измерений
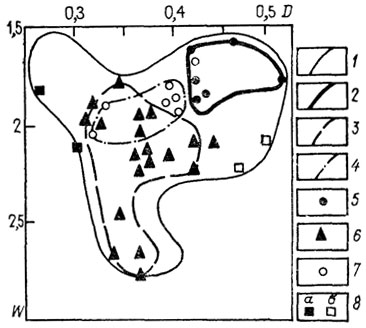
Рис. 25. Схема распределения по форме раковин видов подрода Aulacostephanus (Aulacostephanoceras): 1 - морфологическое поле названного подрода; 2-4 - морфологические поля: 2 - A. (A.) autissiodorensis, 3 - A. (A.) eudoxus, 4 - A. (A.) volgensis; 5-8 - точки результатов измерений раковин видов: 5 - A. (A.) autissiodorensis, 6 - A. (A.) eudoxus, 7 - А. (А) volgensis, 8 - A. (A.) undorae (a) и A. (A.) rigilis (6)
Приведенные данные в целом подтверждают хорошо известный факт: морфологические параметры и, в частности, выбранный метод их математического и графического выражения хорошо отражают специфичность и морфологическую определенность видов. Столь же отчетлива и морфологическая специфичность подродов, родов, семейств и более крупных таксонов. Если считать, что в этих особенностях морфологии заключен функциональный и приспособительный смысл, то, признавая реальность видовой систематической категории, мы логически должны прийти к заключению о не меньшей степени реальности и высших таксономических категорий.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'