
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Реконструкции растений
Вопросы реконструкции водорослей в литературе не освещены. При реконструкции известковых водорослей, изучаемых с помощью шлифов и пришлифовок, главная задача состоит в восстановлении общего вида слоевища по отдельным срезам. Методика последовательных серийных срезов с одного слоевища применяется у этих водорослей очень редко и только для крупных объектов. О несохраняющихся частях, таких, как спорангии и гаметангии, чаще всего приходится судить лишь косвенно, обращаясь к актуалистической модели. При исследовании динофлагеллят приходится изучать отдельно цисты и подвижные стадии (если они вообще известны). Сопоставить те и другие обычно не удается. У кокколитофорид чаще всего приходится иметь дело с разрозненными кокколитами, соединение которых в коккосферы известно лишь в редких случаях.
Способам реконструкции высших растений посвящены как специальные разделы в статьях и монографиях [520, 1341, 1707, 1836 и др.], так и специальные статьи [516 и др.]. Для растений в большей степени, чем для животных, свойственно прижизненное отделение и повторное развитие частей (листьев, генеративных органов). Лишь немногие ископаемые остатки растений, попавшие в захоронения, могут быть сопоставлены по сохранению прижизненного сочетания частей с такими остатками животных, как колонии мшанок и кораллов, раковины фораминифер, моллюсков и брахиопод, скелеты радиолярий и тетрапод, панцири трилобитов и морских ежей. Основная масса палеоботанического материала - это разрозненные листья, куски стеблей, оболочки миоспор, остатки фруктификаций. Понятно поэтому, что для палеоботаники реконструкция прижизненного сочетания разрозненных в захоронении частей - гораздо более сложная и несравненно чаще встающая проблема, чем для палеозоологии,
В целом для реконструкции растений используют все рассмотренные принципы реконструкций, но по сравнению с животными значение некоторых из них больше или меньше. Различие в значении отдельных принципов связано прежде всего с тем, что у растений степень корреляции между разными типами органов часто слабее или неопределеннее. Поэтому внешне очень сходные части могут принадлежать растениям, далеко отстоящим друг от друга в системе, и, следовательно, могут сочетаться с принципиально разными частями. Например, папоротниковидные вайи со строением перышек, свойственным форм-роду Pecopteris, известны у папоротников порядков Zigopteridales, Botryopteridales, Marattiales, Osmundales и Polypodiales (s. 1.), а также у голосеменных порядков Callistophytales и Peltaspermales. Соответственно, найдя листву такого типа и не имея данных о том, с какими органами размножения сочетались вайи обнаруженного вида Pecopteris, мы не сможем реконструировать недостающие органы растения. Эта особенность растений усложняет не только процедуру их реконструкций, но и таксономическо-номенклатурные процедуры (см. 4.1).
Слабость корреляции частей и опасность крупных ошибок при реконструкции растений с помощью актуалистической модели и корреляции были осознаны в палеоботанике не сразу. Более того, это осознание - определенный этап в профессиональном становлении каждого палеоботаника. То же можно сказать и о самом стремлении и умении реконструировать общий облик растений. То и другое появилось в палеоботанике не на заре ее развития, как не сразу появляется и в деятельности палеоботаника, если только его этому не учили специально. Можно назвать целые палеоботанические школы, где издавна практикуется широкое обращение ко всем доступным способам реконструкции изучаемых растений, и школы, где эти реконструкции выполняются редко и лишь тогда, когда имеющийся материал буквально "напрашивается" на эту работу.
Вследствие этого данные о реконструированных растениях и прижизненном сочетании отдельных частей чрезвычайно неравномерно распределены по флорам прошлого во времени и в пространстве, а также по разным систематическим группам. Например, в отношении папоротников существует устойчивая и давняя традиция устанавливать прижизненное сочетание вегетативных и репродуктивных вай. В отношении голосеменных такая традиция существует при изучении, например, юрских флор Европы и лишь в последние десятилетия стала достаточно сильной при изучении триасовых европейских флор. Очень мало реконструированных растений для позднего палеозоя и мезозоя Катазии. Работы по реконструкции позднепалезойских растений Ангариды развернулись лишь в последние годы. Попытки реконструировать прижизненные сочетания органов у покрытосеменных буквально единичны. Неслучайно до сих пор установлены лишь единичные надродовые таксоны вымерших покрытосеменных, хотя сейчас уже ясно, что древнейшие покрытосеменные принадлежали вымершим семействам, порядкам и, может быть, даже классам.
Реконструкции высших растений - важная часть таксономической работы палеоботаника, поскольку, не установив прижизненной связи разных органов, невозможно ввести ископаемые растения в одну систему таксонов с современными растениями. Палеоботаники часто нарушали это общее правило, результатом чего были многочисленные таксономические ошибки, причем некоторые из них повлекли за собой дальнейшие весьма важные ошибочные заключения. Именно так произошло с остатками древнейших покрытосеменных. В домеловых отложениях описывались якобы принадлежащие покрытосеменным листья и пыльца. В меловых отложениях указывались находки листьев современных родов и семейств. Эти заключения не подкреплялись данными о прижизненном сочетании органов. В результате вся проблема происхождения и ранней эволюции покрытосеменных была представлена неправильно, что затормозило и ее решение с помощью палеоботанических данных.
То, что реконструкция растений важна и для решения конкретных стратиграфических задач, не в полной мере осознается до сих пор. Чаще всего при сопоставлении флористических комплексов сравнивается их списочный состав без учета того, что под одинаковыми названиями могут выступать сходные части существенно разных растений. Учет этого обстоятельства особенно важен при сопоставлении разнопровинциальных комплексов растений, когда присутствие общего форм-рода может вообще ничего не значить для стратиграфии. Например, присутствие сходных облиственных побегов хвойных (типа Pseudovoltzla и Ulmannia) в верхней перми (цехштейне) Западной Европы и в казанском ярусе Русской платформы не свидетельствует об одновозрастности вмещающих отложений, поскольку ассоциирующие фруктификации тут и там совершенно разные (более примитивные в казанских отложениях).
В подобных случаях стратиграфические сравнения должны выполняться по растениям, для которых получены хотя бы частичные реконструкции. Правда, здесь мы сталкиваемся с номенклатурной проблемой. В палеоботанике не существует узаконенной процедуры названия реконструированных растений. В. А. Красилов [516] предлагал опираться на название одного из таксонов, входящих в реконструированное сочетание таксонов, и сопровождать это название словом "restitutio" (сокращенно "rest."). Например, Nilssonia Brongniart, 1825 rest. Harris, 1941 - реставрированное растение, соответствующее родам Nilssonia (листья), Beania (женские фруктификации) и Androstrobus (микростробилы). Этот прием пока не получил распространения в литературе. Он неприменим в тех случаях, когда в реконструкцию входит лишь часть каждого из включаемых в нее таксонов. Например, с вайями типа Callipteris ассоциируют женские фруктификации типа Pettaspermum [662] и Autunia [1830]. Сочетание с пельтоидами Peltaspermum известно также у Tatarina и Lepidopteris. Вполне возможно, что в перми США с Autunia сочетались также листья Gigantopteris. Поэтому лучше обозначать реконструируемое растение всем комплексом таксонов, вошедших в реконструкцию, например, Callipteris conferta (Sternb.). - Autunia sp. - Pterispermostrobus gimmianus W. Remy - Cordaicarpus ovoides Weiss - Vesicaspora sp. Для краткости можно брать лишь часть таксонов, если это не чревато путаницей. Скажем, для многих целей приведенное реконструированное растение может быть обозначено без риска путаницы как Callipteris conferta-Autunia - Pterispermostrobus. Тот же способ применим к родам. Например, в перми и триасе встречаются растения, которые можно обозначить как Glossophyllum - Stiphorus, Glossophyllum - Leuthardtia и Kirjamkenia - Stiphorus [662]. Эти комплексные названия обозначают таксономические единицы, которые можно назвать ансамбль-видами и ансамбль-родами.
Идеалом для палеоботаники была бы такая ситуация, когда все таксономические, филогенетические, стратиграфические, фитогеографические и иные исследования опирались бы на ансамбль-роды и ансамбль-виды. Поэтому получение реконструкций для всех ископаемых растений - непреходящая задача палеоботаники, имеющая большое практическое значение.
Как известно, принципы реконструкции растений и животных совпадают, но значение каждого из принципов различно. Чаще всего палеоботаник обращается к совместному захоронению частей и ищет на разных частях общие маркеры (рис. 26), имея в виду какие-то актуалистические или ископаемые модели (рис. 27), которые могут подсказывать определенные корреляции. Иными словами, представления о моделях и корреляциях служат скорее общим теоретическим фоном для заключения, чем ведущим способом работы. Практически процедура реконструкции в палеоботанике варьирует в зависимости от имеющегося материала, который может различаться полнотой сохранности остатков, полнотой представительства разных частей, количеством разных растений, попавших в захоронения. Многое зависит также от технической оснащенности исследования, от ограничений на время его выполнения, от полноты коллекций и просто от квалификации палеоботаника.
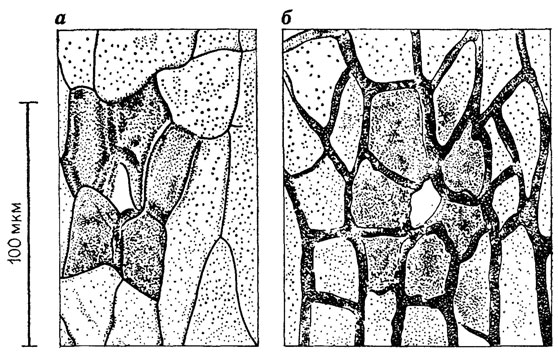
Рис. 26. Однотипные устьица на семеносной капсуле Leptostrobus laxiflora Heer (а) и листе Czekanowskia rigida Heer (б), принадлежащих одним и тем же растениям, верхняя юра Буреинского бассейна. По В. Л. Вахрамееву, М. П. Долуденко
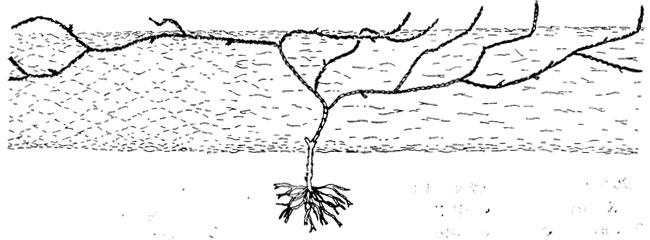
Рис. 27. Реконструкция среднекаменноугольной Selaginella fraipontii (Lecl.) Schlank. et Leism., показанной на фоне ствола лепидодендрона. По К. М. Шланкеру, Г. Э. Лейстману. Моделью реконструкции служили современные виды Selaginella
Разбирать детально специфику работы для всевозможных сочетаний перечисленных условий здесь невозможно. Поэтому ограничимся некоторыми показательными примерами, взяв как удачные и впоследствии подтвердившиеся, так и оказавшиеся ошибочными реконструкции.
Одной из реконструкций, оказавших немалое влияние на палеоботанику в целом, было установление прижизненной связи между семеносными купулами Lagenostoma, папоротниковидными вайями типа Sphenopteris hoeninghausii Brongn. и стеблями типа Lyginopteris oldhamia Binney. Благодаря этому открытию утвердилось представление о птеридоспермах как о растениях с семенами и папоротниковидными вайями. Для выполненной реконструкции решающую роль сыграло совместное нахождение разных частей и наличие маркеров, в качестве которых выступили характерные булавовидные железки на купулах и вегетативных органах. Соединение листьев и стеблей подкреплялось присутствием у тех и других своеобразной коры Dictyoxylon - типа. Придаточные корни тех же растений, ранее описанные как Kaloxylon hookeri Will., были найдены в прикреплении. В результате проведенных исследований удалось дать весьма правдоподобное изображение общего облика всего растения [2255]. Последующие исследования только дополняли реконструкцию, но не изменили ее в существенных чертах.
Показательно, что эта, ставшая классической, реконструкция выполнялась фактически без обращения к какой-либо актуалистической модели, а принцип корреляции использовался в ней только в том отношении, что была постулирована высокая корреляция маркера (булавовидных железок) с разными частями растения. Фактически же реконструкция опиралась преимущественно на маркер и сонахождение частей, обнаружение прижизненной связи некоторых частей играло второстепенную роль.
Сочетание совместного захоронения и маркера широко использовалось и при изготовлении других реконструкций. С. Леклерк и X. П. Бэнкс [1870] реконструировали кладоксилеевое Pseudosporochnus nodosus Lecl. et Banks, опираясь на сонахождение разных частей, несущих одинаковые склеренхимные гнезда в коре. Установление прижизненной связи стеблей Callistophyton, листьев и семян Callospermarion и синангиев Idanothekion (или Callandrium) основано на одинаковых секреторных органах и сонахождении всех этих частей [2180]. В качестве маркеров не обязательно выступают микроструктурные признаки. В. А Красилов [520], например, присутствие одинаковых катафиллов (чешуевидных листьев) у побегов Pseudotorellia angustifolia Dolud. и проблематических фруктификаций Umaltolepis rarinervis Krassll. считал доказательством их прижизненной связи.
Т. М. Гаррис в ряде работ, посвященных реконструкциям, приводил специальное обоснование используемых приемов. Так, он сформулировал следующее важное методическое положение: если мы встречаем в захоронении репродуктивные органы каких-либо растений, то вместе с ними должны быть захоронены и вегетативные части тех же растений [1707]. Исключения из этого правила очень редки (например, когда остатки испытали сильный перенос и сортировку по размерам и устойчивости частей). В таких случаях приходится сталкиваться со скоплениями семян, шишек, плодов без сопутствующих листьев или с такими их фрагментами, которые не поддаются идентификации. Следовательно, для Т. М. Гарриса сонахождение было ведущим критерием установления прижизненной связи дисперсных частей. Иногда, если это допускали общие морфологические предпосылки, он заключал о связи частей только по этому признаку. При анализе захоронения Т. М. Гаррис отмечал те компоненты, для которых прижизненное сочетание частей было установлено ранее; оставшиеся компоненты иногда удавалось воссоединять, уже опираясь на метод исключения. Кроме того, Т. М. Гаррис широко использовал в качестве маркеров эпидермально-кутикулярные признаки, сходные смоляные тельца в листьях, стеблях и фруктификациях, однотипную пыльцу в микроспорангиях и в микропиле семян. Он впервые обратил внимание на то, что сонахождение остатков листьев, семян и пыльцы в копролитах также может указывать на прижизненное сочетание. Этот прием позже позволил подкрепить соображения о прижизненной связи синангиев Pterispermostrobus gimmlatius (с пыльцой Vesicaspora) и листьев Callipteris conferta [662], а также синангиев Permotheca (с пыльцой Protohaploxypinus) и листьев Tatarina [659].
Остановимся теперь на некоторых поучительных неудавшихся реконструкциях. В литературе более столетия фигурировала и стала канонической реконструкция Psilophyton princeps Daws., которая, как оказалось [1760], объединила части трех совершенно разных растений. Ничего, кроме сонахождения этих растений, не было принято во внимание при изготовлении этой реконструкции.
Широко вошла в литературу также реконструкция Williamsonia sewardiana Sahni из мезозоя Индии. На реконструкции изображено растение, сходное с Cycas и имеющее листья типа Ptilophyllum cutchense Morris. Сейчас эта реконструкция отвергнута [1359]. Оказалось, что выбранная модель не подкрепляется данными о размерах стеблей. Листья P. cutchense не встречены в одном захоронении с W. sewardiana и имеют различные эпидермально-кутикулярные признаки. Достоверных данных оказалось недостаточно для новой реконструкции взамен отвергнутой.
В приведенных и других случаях ошибочных реконструкций палеоботаниками не были приняты во внимание тот или иной принципы. При реконструкции Psilophyton учитывалось только сонахождение остатков. Во втором примере решающую роль сыграла неверная выборка модели. Серьезные ошибки делаются и в тех случаях, когда палеоботаники вовсе не пытаются делать реконструкций, не используют имеющиеся сведения о прижизненном сочетании частей, а судят об их систематической принадлежности порознь, по отдельным наблюдаемым признакам. Именно так случилось с древнейшими покрытосеменными в разобранном примере.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'