
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Реконструкции беспозвоночных организмов
К настоящему времени накопился весьма интересный и разнообразный материал по реконструкциям различных вымерших беспозвоночных организмов: общего облика животных, отдельных их органов и систем, а также образа жизни.
Реконструкции беспозвоночных организмов основываются, естественно, на принципах, сформулированных в начале данной главы. Однако всякий раз дают о себе знать и специфические особенности изучаемых объектов и среди них прежде всего характер и степень сохранности остатков.
Наиболее распространенный прием реконструкций беспозвоночных - сравнение вымерших организмов с ныне живущими представителями той же самой или близкородственной (принцип гомологии), реже неродственной (принцип аналогии) группы. Для тех организмов, современные представители которых не установлены, на первое место выходит один из более сложных приемов морфофункционального анализа, так называемый парадигматический подход [864]. В этом случае для определенной структуры или органа подбираются наиболее вероятные функции и для проверки каждой из них строятся гипотетические модели - парадигмы.
Исключительно ценными являются как прямые (нахождение фоссилий в прижизненной ориентировке), так и косвенные палеоэкологические данные - характер расположения и ориентировки симбионтов, поселявшихся прижизненно на поверхности тела или внутри покровов организма-хозяина, т. п. В последнем случае нередко удается расшифровать не только положение организма при жизни, но и функции его органов и систем, в первую очередь механизма поступления внутрь животного воды и пищевых частиц. Независимо от выбора способа реконструкция достоверна только в том случае, когда исследуется окаменелость хорошей сохранности. Очевидно, что при реконструкциях организмов с внутренним скелетом немаловажную, а иногда и первостепенную роль играют отпечатки мягкого тела.
Остановимся на некоторых примерах реконструкций различных, но конечно же далеко не всех беспозвоночных организмов, иллюстрирующих высказанные общие положения.
Весьма наглядно значение актуалистического подхода к реконструкциям беспозвоночных может быть показано на примере моноплакофор. Их одностворчатые, обычно мелкие колпачковидные или блюдцеобразные раковинки с почти центральной или эксцентричной вершиной и округлым или овальным устьем известны из нижне- и среднепалеозойских отложений многих геологических регионов. На внутренней поверхности многих раковин различимы от пяти до восьми пар преимущественно овальных лункообразных углублений, расположенных по кругу или подковообразно между центром раковины и ее краями, которые рассматривались как мускульные отпечатки. На основании формы раковин и характера отпечатков моноплакофор относили первоначально к брюхоногим моллюскам, причем выделяли в качестве семейства, затем отряда и подкласса.
После обнаружения в 1952 г. в Тихом океане глубоководных моллюсков Neopilina galatheae появилась возможность осмысливания ископаемого материала под новым углом зрения и создания реконструкции тела и образа жизни некоторых древних родов, объединяемых теперь вместе с Neopilina в один класс Monoplacophora. На рис. 28 приведена реконструкция тела моноплакофоры Scenella из среднего кембрия Канады [2193], основывающаяся на изучении внутреннего ядра этой блюдцевидной раковины с шестью парами мелких, расположенных билатерально симметрично мускульных отпечатков. С учетом данных о строении мягкого тела Neopilina предполагают, что древние моноплакофоры имели широкую плоскую ногу, с помощью которой ползали по поверхности субстрата.
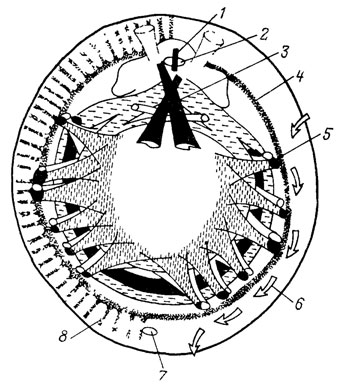
Рис. 28. Реконструкция среднекембрийского моллюска Scenella sp. (класс, Monoplacophora), Канада, Британская Колумбия. Ув. 4. (вид с нижней стороны тела). По Ф. Расетти из Б. Раннегара и Ж. Пожет. 1 - радула; 2 - ротовое отверстие; 3 - мускулы переднего отдела тела (первая пара гипотетична); 4 - мантийная линия; 5 - мускулы ноги; 6 - вероятное направление движения водного потока через мантийную полость; 7 - анальное отверстие; 8 - мантийные мускулы
Обратимся теперь к другому примеру реконструкций, когда из-за отсутствия среди современных беспозвоночных не только родственных, но даже отдаленно сходных форм сравнительно-актуалистический подход не мог быть использован. Показательными в этом отношении являются, в частности, рудисты, обладающие неповторимыми в других группах двустворчатых моллюсков формой раковин и внутренними структурами. Известно, что вследствие морфологического своеобразия некоторых их представителей первоначально относили к брахиоподам, кораллам, головоногим моллюскам и даже к усоногим ракам.
Особенно сложной для расшифровки функций внутренних скелетных элементов и реконструкции мягких тканей оказалась одна из наиболее распространенных групп рудистов - гиппуриты. Очень интересна их синтетическая реконструкция [2292], воспроизводящая функции элементов скелета, строение мягкого тела, механизм питания и газообмена (рис. 29).
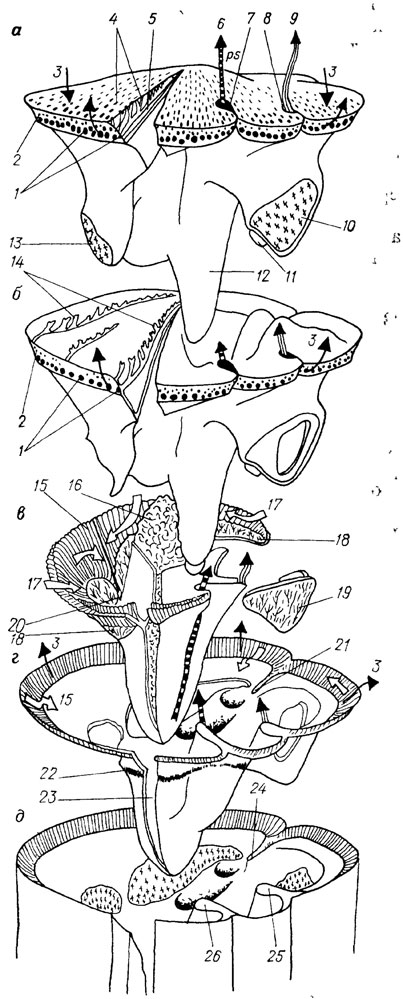
Рис. 29. Реконструкция позднемелового гиппурита (класс Bivalvia, отряд Hippuritida), Франция. По П. В. Скелтону с упрощениями. а - верхняя створка; б - верхняя или левая доля мантии; в - тело моллюска; г - нижняя или правая доля мантии; д - верхняя часть нижней створки; 1 - краевые устья радиальных каналов; 2 - краевые устья пор; 3 - направление водных потоков; 4 - вводные поры; 5 - радиальные каналы; 6 - поток псевдофекалий (ps); 7 - оскулюм S; 8 - оскулюм S; 9 - поток фекалий (f); 10 - задняя миофорная апофиза; 11 - задний зуб; 12 - передний зуб; 13 - передняя миофорная апофиза; 14 - мантийная выстилка внутри пор и радиальных каналов; 15 - жабры; 16 - висцеральная масса; 17 - направление движения частиц, перемещаемых ресничным эпителием мантии и жабр; 18 - передний аддуктор; 19 - задний аддуктор; 20 - ротовое отверстие; 21 - лигамент; 22 - мантийная линия; 23 - мантийная полость; 24 - лигаментный гребень 25 - пилляр S; 26 - пилляр Е
В основу реконструкции положен детальный морфофункциональный анализ раковин Hippurites radiosus, H. toucasi и Н. socialis.
Было установлено, что аддукторы у названных видов очень короткие. Например, у H. socialis при диаметре раковины 20 мм и при сомкнутых створках зазор для переднего мускула-замыкателя составлял всего 1 мм, а заднего - около 0,6 мм. Максимально возможное растяжение мускула могло обеспечить увеличение зияния вдоль краев створок всего лишь на 0,5-0,8 мм. Поэтому было высказано предположение, что гиппуриты не открывали и не закрывали створки раковины. Отверстие между краями створок было постоянным и составляло доли миллиметра. В этом случае лигамент мог полностью атрофироваться, так как его функцию стали выполнять аддукторы, действуя подобно тонким, но широким прокладкам липкого вещества. Водный поток, несущий пищевые частицы, поступал в организм через ирригационную систему - наружные поры и внутренние радиальные каналы в верхней створке.
Ток воды создавался движением ресничек мантийной выстилки на стенках пор и каналов. Так как система каналов неизменна и непрерывна по периферии створок, то вода из них поступала на всю поверхность мантийной лопасти нижней створки. Благодаря действию ресничного эпителия мантии из воды отфильтровывались пищевые частицы, которые направлялись вдоль желобка у внутреннего края лимба в передневентральную область раковины, а очищенная вода отводилась наружу через узкую щель между открытыми краями нижней и верхней лопастей мантии. Таким образом, у гиппуритов наружные мантийные лопасти стали выполнять роль аппарата улавливания пищевых частиц.
Однонаправленный поток пищевых частиц в передневентральной области нижней створки попадал на жабры и после очередной сортировки направлялся к ротовому отверстию, расположенному между двумя долями переднего аддуктора. Возможно, в процессе отбора пищевых частиц участвовали и ротовые лопасти. Нежелательные частицы оставались в мантийной полости и как псевдофекалии выводились по псевдосифону наружу через вентральный оскулюм. Анальное отверстие, через которое из организма выводились фекалии, находилось вблизи дорзального оскулюма. Газообмен осуществлялся жабрами; возможно, в этом процессе принимали участие и мантийные лопасти.
Реконструкции мягкого тела и функций отдельных систем и органов выполнены также и для других рудистов: Diceras [2293], Radiolites [2294] и др. На основании изучения микроструктуры стенки верхней створки рудистов Osculigera в сопоставлении с современными двустворками Corculum были получены доказательства симбиоза оскулигер с зелеными водорослями (2428].
Все сказанное о реконструкциях рудистов представляет интерес и для палеофизиологических исследований (см. 1.6).
Наглядный пример парадигматического подхода к реконструкции механизма питания пермских брахиопод Prorichthofenia приведен М. Радвиком [2189]. Весьма своеобразная раковина этих брахиопод состоит из двух неодинаковых по форме и размерам створок - конической нижней и очень маленькой плоской верхней. Согласно М. Радвику, верхняя створка была подвижной и могла открываться и закрываться. При этом возникал эффект насоса, благодаря которому к лофофорам поступал ток свежей воды с органическим детритом и удалялась вода, прошедшая через пищеварительный тракт организма. В процессе разработки реконструкции М. Радвик использовал моделирование движений верхней створки, помещенной в водную среду с суспензионными каплями масла, причем направление и циркуляция водного потока фиксировались путем киносъемки.
Надежным приемом является восстановление строения беспозвоночных организмов или их отдельных частей по фоссилизированным остаткам мягкого тела. Благодаря быстрому постмортальному замещению мягких тканей теми или иными фоссилизирующими минеральными соединениями иногда образуются биоморфозы по тонким внутренним структурам (например, кишечному тракту). Подобные реконструкции известны в ряде работ [687, 1413, 1458 и др.].
Оригинальный способ реконструкции наружного облика тела и прижизненного положения прямых ортоцератид на дне моря, основывающийся на изучении совместного нахождения раковин цефалопод и отпечатков их щупалец на поверхности грубозернистой породы из верхнего ордовика США, использован Р. Г. Флауэром [1587].
Ранее уже было сказано, что при создании реконструкций и их использовании в ходе дальнейшего изучения каждой группы ископаемых важное значение имеют морфологические особенности и характер сохранности исследуемых объектов.
Например, членистоногие заключенные в прочный хитиновый панцирь, который у морских форм еще часто обызвествляется, сохраняются весьма полно и для понимания строения животного обычно не требуется реконструкций. Лишь в отдельных случаях, как, например, с остатками из знаменитых кембрийских сланцев. Беджес в течение более чем 50 лет обсуждалось фактически не строение ископаемых, а их реконструкции, сделанные тем или иным исследователем в зависимости от его представлений о систематических связях ископаемых форм. В последнее время, наоборот, предприняты новые детальные исследования, которые показали необоснованность большинства этих реконструкций.
Особое положение среди членистоногих занимают насекомые. Они редко сохраняются полностью Обычно это только изолированные крылья или даже части крыльев. Поэтому суждения о систематическом положении насекомых неизбежно оказываются реконструкциями. Ископаемые остатки, как правило, принадлежат взрослым формам. Лишь для немногих водных насекомых известны и личинки. Личиночные стадии насекомых могут вести совершенно иной по сравнению со взрослыми особями образ жизни и иметь отличный от них внешний вид. Суждение о систематическом положении древних насекомых, таким образом, включает и определенные представления о их жизненном цикле и о строении личинок. В некоторых случаях жизненный цикл и строение разных стадий развития древних насекомых может быть реконструирован с большой полнотой. Так, для раннемеловых комаров рода Chironomaptera известны находки яиц, личинок, куколок и взрослых насекомых. При этом, правда, возникает проблема установления принадлежности разных стадий к единому циклу развития, к одному виду. К счастью, в местонахождении редко встречается более одного вида того же рода, представленного массовыми находками его остатков.
Изредка сохранность ископаемых остатков позволяет дать детальные реконструкции древних насекомых. Особенно подробные реконструкции могут быть созданы для некоторых водных форм, находки ископаемых остатков которых весьма многочисленны.
В качестве примера можно привести реконструкцию водного жука Coptoclava longipoda Ping [830], для которого известны к настоящему времени тысячи находок личинок и сотни взрослых жуков. Реконструирование этого насекомого тем более интересно, что и взрослый жук и особенно личинка совершенно непохожи на других водных жуков Личинка была обитателем вод больших мезозойских озер, на дне которых существовали заморные условия и отсутствовала прикрепленная водная растительность. В этих условиях личинка жука вела исключительно нектонный образ жизни, передние ноги превратились в хватательные, вооруженные шипами, средние и задние - в гребные с уплощенной и расширенной пластинкой (рис. 30). Внешний вид личинки настолько необычен, что сначала были сомнения в принадлежности ее остатков не только к жукам, но и насекомым вообще. Основные трахейные стволы личинки превращены в гидростатический аппарат с мощным спиральным утолщением. Изменяя объем заключенного в трахеи воздуха, личинка могла менять плотность и перемещаться в воде в вертикальном направлении.
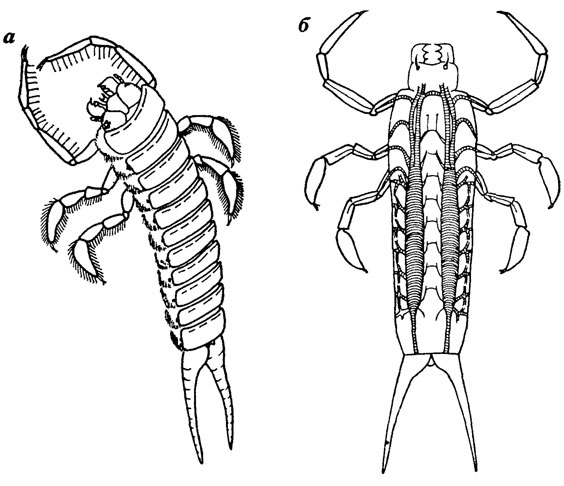
Рис. 30. Реконструкция личинки раннемелового жука Coptoclava longipoda Ping. По А. Г. Пономаренко. а - внешний вид б - трахейная система
Склеротизация трахей не уступает степени склеротизации наружных покровов, что позволило дать достаточно содержательную реконструкцию строения трахейной системы древнего насекомого. Хотя коптоклава принадлежит к дитискоидным жукам, в ее строении оказалось много черт, делающих ее похожей не на других дитискоидов, а на вертячек: разделенные на две пары - верхнюю и нижнюю - глаза, длинные хватательные передние и гребные уплощенные и расширенные средние и задние ноги (рис. 31).
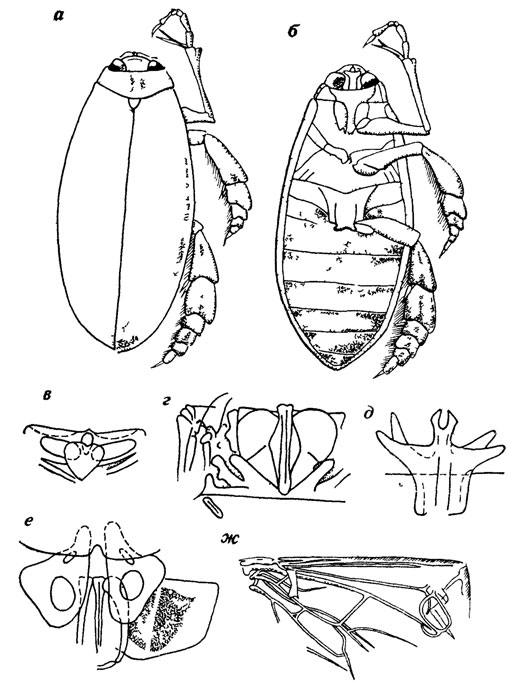
Рис. 31. Реконструкция взрослого жука Coptoclava longipoda Ping. По А. Г. Пономаренко. а - вид сверху, б - вид снизу, в - среднеспинка, г - заднеспинка, д - эндоскелет заднегруди, е - конец брюшка и генитальные придатки самки, ж - крыло
Вероятно, как и вертячки, этот жук обитал на поверхностной пленке воды, собирая падающих на нее насекомых или охотясь на выходящих из куколок комаров. Для подобного образа жизни жук имел большие размеры и поэтому покровы его были тонкие. Он оказался благодатным объектом для изучения и реконструкции внутреннего скелета и тех частей наружного, которые обычно у жука скрыты - крылья, нижняя сторона надкрылий и их запирательный аппарат, верхняя сторона груди и брюшка, генитальные придатки (см. рис. 31). В результате удалось восстановить строение жука почти полностью, с детальностью, известной лишь для немногих современных форм. Анализ строения позволил реконструировать и некоторые физиологические и поведенческие характеристики коптоклав - характер питания, плавания, вплоть до частоты гребных движений.
Реконструкция коптоклавы преимущественно направлена на выяснение как можно большего числа черт этого своеобразного вида, понимания его систематического положения и экологической роли.
Более распространенный тип реконструкций - иллюстративные, на них обычно изображаются представители полностью вымерших или, наоборот, древние представители существующих форм. В последнем случае главной целью реконструкции является стремление показать сохранение насекомыми сходного строения за десятки и сотни миллионов лет. Эти реконструкции часто бывают составными крылья от одного вида, тело, ноги, антенны часто от иных или синтетические, реконструируемые по признакам многих родственных форм, вымерших или современных. Эти реконструкции лучше смотрятся, если насекомому придается "оживленная" поза. Для этого лучше использовать фотографии близких современных форм, что позволяет несколько уменьшить схематизм реконструкции.
Иллюстративные реконструкции широко распространены, хотя часто бывают ошибочными. Так, вплоть до настоящего времени публикуются реконструкции каменноугольных палеодиктиоптер с грызущим ротовым аппаратом. Эти реконструкции делались в то время, когда была уверенность, что насекомое со столь примитивным строением крыльев могло иметь только неспециализированный грызущий ротовой аппарат. Реконструкции создавались и продолжают публиковаться, хотя все ископаемые остатки палеодиктиоптер с сохранившимися ротовыми органами имели сосущие хоботки. Иллюстративная реконструкция палеодиктиоптеры показана на рис. 32. В издании, откуда она воспроизведена [450], имеются и другие подобные реконструкции древних насекомых.
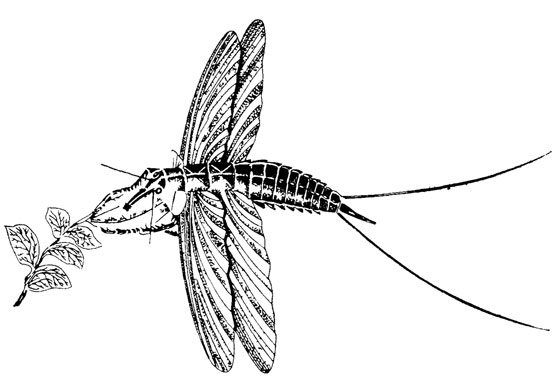
Рис. 32. Реконструкция каменноугольной палеодиктеоптеры Goldenbergia sp. По А. Г. Пономаренко
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'