
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Реконструкции позвоночных животных
Возможность восстановления облика вымерших позвоночных всегда интересовала палеонтологов, попытки дать изображения этих животных предпринимались с самого момента оформления палеонтологии как самостоятельной науки. Эти попытки имели не только чисто научное значение - сейчас мы не можем даже представить себе книгу об эволюции животного мира или музей естественной истории без художественных изображений животных, населявших нашу Землю в прошлом. И тем не менее, несмотря на огромное количество опубликованных реконструкций представителей большинства групп вымерших позвоночных, в современной палеонтологической литературе отсутствуют обобщающие работы, специально посвященные данной проблеме.
Рассматривая проблему реконструкций, прежде всего нужно помнить, что она имеет два самостоятельных аспекта. Первый аспект - реконструкция строения целого организма по тем ископаемым остаткам, которые попадают в руки палеонтологов; эта проблема - чисто научная. Другая сторона вопроса - реконструкция внешнего облика вымерших позвоночных животных и возможность их художественного изображения. Эта задача уже комплексная, требующая совместной работы специалиста-палеонтолога и художника-анималиста. Возможны, конечно, счастливые исключения, когда оба эти качества совмещаются в одном лице. Широко известны, например, реконструкции крупных советских палеонтологов А. П. Быстрова и К. К. Флерова, однако в большинстве случаев реконструкции становятся результатом творческого содружества ученых и художников. При этом, естественно, фантазия художника не должна выходить за рамки научной обоснованности.
Работа по восстановлению облика позвоночных животных чаще всего начинается с изучения палеонтологом строения скелета, поскольку в подавляющем большинстве случаев в руки ученых попадают именно костные остатки. Как указывал А. П. Быстров [125], правильно смонтированный скелет является достаточным каркасом для того, чтобы избежать особенно грубых ошибок при реконструкции общих очертаний тела. На основании строения скелета можно правильно представить себе пропорции основных частей тела животного, пределы их возможных движений. К сожалению, находки полных скелетов редки, чаще всего палеонтолог имеет дело с их фрагментами или даже с отдельными костями. Поэтому перед реставраторами обычно встает задача не реконструкции облика животного по скелету, а реконструкция самого скелета в целом по его частям.
Путей таких реконструкций может быть несколько. Прежде всего известны многочисленные случаи находок различных фрагментов скелетов разных экземпляров одного и того же вида, что дает возможность "собрать" полный или почти полный его скелет. Однако при этом для реконструкции естественных пропорций тела важно использовать кости животных одного размерного класса. Но даже и тогда, когда материал по отдельным видам ограничивается фрагментами скелета, палеонтолог иногда может реконструировать недостающие части. Определенной опорой здесь могут стать аналогии с другими известными представителями той же группы животных и обоснованные основоположником палеонтологии позвоночных Ж. Кювье принципы корреляций и соподчинения ("subordination") органов, согласно которым строение одних органов связано с определенными особенностями строения остальных.
В то же время, говоря о корреляциях в строении различных органов животных, мы не должны забывать, что они часто отражают не столько причинные связи, сколько эмпирически установленные соотношения (например, рогатые млекопитающие непременно должны быть копытными). Но мир животных чрезвычайно разнообразен, и реальные особенности строения отдельных его представителей могут иногда выходить за рамки привычных нам соотношений. Вопрос в том, насколько хорошо мы представляем все разнообразие строения животных различных таксонов, отдельных представителей которых мы реконструируем, и есть ли в пределах интересующих нас групп животных формы, уклоняющиеся по каким-либо признакам от характерного для этих групп общего типа строения. Например, своеобразный представитель непарнокопытных халикотерий имел на пальцах не копыта, а когти, хотя по всем остальным чертам своего строения он, несомненно, должен быть отнесен к копытным. По этой особенности строения конечностей халикотерий не имеет аналогов среди других копытных, и если бы скелет его конечностей не был найден, вряд ли мы могли бы даже вообразить такое строение его пальцев, явно противоречащее всем нашим привычным представлениям.
Другой иллюстрацией сказанного может служить история реконструкции Г. Ф. Осборном и А. А. Борисяком скелетов гигантских безрогих носорогов - белуджитерия и близкого к нему индрикатерия [109]. Зная первоначально один только череп белуджитерия, Г. Ф. Осборн при попытке реконструкции всего животного придал его телу вполне "носорожий" (похожий на современных носорогов) облик (рис. 33, а). Однако последующие находки показали, что тело белуджитерия было совсем не таким, как у других известных ранее носорогов. После того как Г. Ф. Осборн ознакомился со сделанным А. А. Борисяком описанием скелета конечностей очень близкого к этому виду индрикатерия, его реконструкция получила иной вид (см. рис. 33, б). Сам же А. А. Борисяк шел обратным путем. Не имея черепа, он в своей реконструкции индрикатерия придал слишком "носорожий" характер голове. И только когда он ознакомился со сделанным Г. Ф. Осборном описанием черепа белуджитерия, реконструкция скелетов этих носорогов стала вполне обоснованной (см. рис. 33, в).
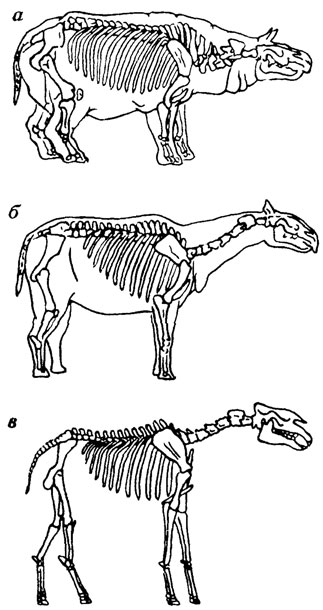
Рис. 33. Реконструкции скелета белуджитерия и индрикатерия: а - первая реконструкция белуджитерия (по Г. Осборну из работы А. Л. Борисяка); б - вторая реконструкция белуджитерия (по Г. Осборну из работы Л. А. Борисяка); в - реконструкция индрикатерия (по А. А. Борисяку)
Таким образом, возможность использования при реконструкциях аналогий и эмпирически выведенных корреляций определяется прежде всего полнотой нашего знания о строении животных, принадлежащих к отдельным группам. В тех случаях, когда среди материала по реконструируемым видам отсутствуют отдельные части их скелета, аналогии с уже известными науке животными не всегда могут служить гарантией правильности предлагаемых реконструкций, поскольку именно строение не представленных в находках частей может уклоняться от общего типа. В такой ситуации чаще всего целесообразно вообще отказаться от попыток реконструкции всего животного целиком и реконструировать только те части тела, по которым имеется скелетный материал, особенно если мы имеем дело с представителями групп, не имеющих близкого родства с ныне существующими животными [953].
Изучение строения скелета вымерших животных - начальный этап подготовки к реконструкции их внешнего облика, имеющий очень важное значение. Эта работа позволяет не только представить пропорции и самые общие контуры тела реконструируемых животных. Изучение костей дает палеонтологу информацию о строении имевшихся в скелете хрящевых образований, об их мускулатуре и о других мягких тканях, необходимую для более полного представления о внешнем виде ископаемых животных.
Основой реконструкции таких образований, как хрящевые элементы черепа и висцерального скелета, сосудистая и нервная системы, являются отпечатки хряща на поверхности костей, борозды на их поверхности, по которым проходят кровеносные сосуды и нервы, отверстия для сосудов и нервов в черепных костях [127, 1030 и др.], а о некоторых чертах строения головного мозга можно иногда получить представление по сохраняющимся естественным отливам мозговых капсул черепа [786, 787, 2116, 2117]. Реконструкция подобных структур представляет собой совершенно особую задачу, не всегда имеющую непосредственную связь с проблемой восстановления внешнего вида животных.
Основная задача подобных реконструкций - восстановление внутреннего строения животных, а это в первую очередь важно для решения проблемы филогенетических связей между различными группами [1030, 1176]. Так, именно на основе реконструкции хрящевых структур нейрокраниума, висцерального скелета и плечевого пояса палеозойских Ostracodermi шведский палеонтолог Э. Стенше установил их принадлежность к ветви древнейших бесчелюстных [106], а его соотечественник Г. Сёве-Сёдерберг [2203] на основе подобных реконструкций получил морфологические доказательства происхождения современных амфибий группы Anura от лабиринтодонтов.
Следует сказать, что успешная реконструкция хрящевого скелета, сосудистой и нервной систем может дать и определенную информацию о некоторых чертах внешнего облика вымерших животных. Например, на основе реконструкции сосудов и нервов головы зверозубых рептилий Л. П. Татаринов [1030] высказал предположение о наличии у некоторых из них мягких губ и осязательных придатков типа вибрисс. Иногда подобные указания могут быть более косвенными. Изучение отливов мозговых капсул ископаемых выдр позволило Л. Радинскому [2116] предположить, что у видов группы Lutra, живших около 10 миллионов лет назад, была хорошо развита тактильная чувствительность передней части головы, что должно было выразиться в сильном развитии вибрисс; напротив, у выдр группы Aonyx, существовавших в это же время, была, по-видимому, развита тактильная чувствительность передних конечностей. Такие соображения могут быть использованы при попытках изображения этих животных.
Очень большое значение для успешного восстановления внешнего облика вымерших животных имеет реконструкция мышечной системы, поскольку многие характерные черты их внешнего вида могут определяться рельефом мускулатуры. О степени развитости отдельных мышц можно судить прежде всего по наличию и размерам впадин, островков, бугров и гребней на костях, к которым эти мускулы крепятся. Так, характер сагиттального гребня и височных впадин на черепе млекопитающих связаны со степенью развития жевательной мускулатуры, строение невральных дуг и остистых отростков позвонков позволяют судить о мускулатуре спины и шеи, мозолистые бугры на костях конечностей свидетельствуют о развитости их сгибателей и разгибателей и т. д. Все это позволяет палеонтологу получить представление о рельефе мускулатуры животных, необходимое для реконструкции их внешнего вида в целом.
С другой стороны, при реконструкции мускулатуры, как и при восстановлении недостающих частей скелета, определенную роль могут играть аналогии с современными представителями тех групп, к которым относятся реконструируемые животные. При этом чем ближе родство ископаемых и современных животных, тем более точной может быть основанная на аналогиях реконструкция. Именно знание строения мускулатуры современных лошадей позволило В. И. Жегалло [365] осуществить детальную реконструкцию мускулатуры головы близкого к ним гиппариона. Для представителей же полностью вымерших групп животных возможны, как правило, лишь обобщенные реконструкции мускулатуры и только в отдельных случаях удается относительно детально восстановить строение отдельных систем мышц, например челюстной мускулатуры [2066, 2205].
Наиболее детальная реконструкция мягких тканей возможна в тех случаях, когда соотношение между рельефом скелета и толщиной его мягких покровов может быть подробно изучено на рецентном материале. Лучшим примером здесь могут служить работы антрополога М. М. Герасимова [239] по восстановлению индивидуальных черт лица человека и его ближайших предков по черепу. Метод М. М. Герасимова основан на корреляциях между толщиной мягких тканей и развитием определенных элементов черепа человека, анатомия которого известна до мельчайших подробностей.
На опыт работы М. М. Герасимова может иногда опираться и палеонтолог - в тех, например, случаях, когда реконструируемые вымершие виды имеют в современной фауне близкородственные формы, которые могут быть изучены с максимальной степенью детальности. Так, на изучении мягких тканей современных слонов базируются реконструкции головы мамонта и ряда других вымерших слонов, выполненные В. Е. Гаруттом [1626]. Основой этих реконструкций послужило определение толщины слоя мягких тканей на различных участках головы слонов, проведенное как путем препарирования трупного материала, так и методом фотографического совмещения изображений черепов слонов, павших в зоопарках, с их прижизненными фотографиями. Полученные таким путем показатели толщины слоя мягких тканей головы наносились затем на профильные изображения черепов ископаемых слонов, что давало возможность воспроизвести очертания их головы (рис. 34).
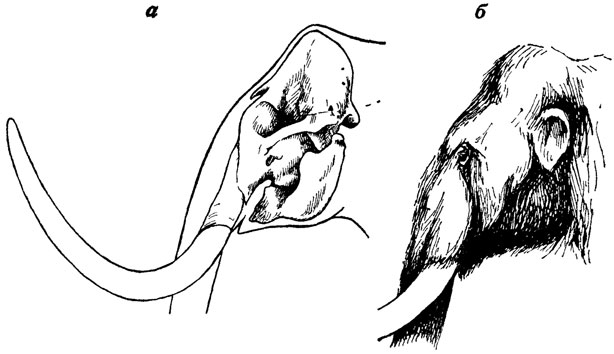
Рис. 34. Реконструкция головы мамонта. По В. Е. Гарутту. а - череп и контуры головы, б - внешний вид головы
Наружные покровы редко сохраняются в ископаемом состоянии. Идеальный для палеонтолога случай - находки в вечной мерзлоте целых туш зверей (например, мамонтов или шерстистых носорогов), что дает возможность не реконструировать, а документально описывать их покровы. Однако такие случаи редки, обычно при находках ископаемых остатков млекопитающих от их шерсти не остается никаких следов.
При реконструкции волосяного покрова можно опираться и на некоторые косвенные соображения. Так, у грызунов с роющим образом жизни (о чем можно судить по строению их конечностей и зубов) шерсть должна была быть плотной и гладкой; звери, обитавшие в условиях холодного климата, должны были бы иметь густую и длинную шерсть и т. д. С покровами позвоночных остальных классов дело иногда обстоит иначе Костная чешуя рыб может сохраняться достаточно хорошо. Известны случаи находок отпечатков покрытой чешуей кожи некоторых рептилий и оперения птиц - достаточно вспомнить отпечатки археоптерикса с прекрасно сохранившимися следами перьев, крыльев и хвоста. Иногда такие отпечатки дают довольно неожиданную картину. Так, А. Г. Шаровым [1154] в Каратау был найден отпечаток птерозавра, названного им Sordes pilosus, со следами покровов, напоминающих волосы млекопитающих. Но чаще всего палеонтолог может только выдвигать более или менее обоснованные предположения о характере наружных покровов вымерших животных.
Наконец, следует сказать несколько слов о рогах, клювах, хоботах, когтях, копытах и подобных образованиях. Рога оленей часто хорошо сохраняются, по найденным остаткам о них можно получить достаточно полное представление. Напротив, роговые чехлы полорогих не сохраняются, но некоторое представление об их размерах и форме дают сохраняющиеся их костные основания. О наличии или отсутствии рогов на морде вымерших носорогов можно судить по бугристым образованиям на поверхности костей черепа; известны и отдельные находки сохранившихся целых рогов плейстоценовых шерстистых носорогов. О наличии у зверей хобота мы можем судить по форме носовых костей, однако определенных указаний на длину этого хобота кости черепа не дают. О присутствии роговых клювов у некоторых ископаемых рептилий также можно судить по форме их челюстных костей и по остающимся на этих костях следам прикрепления роговых чехлов, о форме самих чехлов чаще всего можно только строить предположения. То же самое можно сказать и о когтях или копытах на ногах - хотя сами они, как правило, не сохраняются, об их форме и размерах можно судить по конечным фалангам пальцев. Иногда удается получить свидетельства о наличии плавательных перепонок между пальцами, в частности, о перепонках на ногах некоторых динозавров можно судить по известным находкам их следов [210].
Таким образом, палеонтолог может получить достаточно свидетельств о многих характерных чертах внешнего облика вымерших животных. Однако на этом подготовительная работа к созданию собственно реконструкции не заканчивается. Прежде чем приступать к попыткам дать научно обоснованное художественное изображение, необходимо провести по возможности полный (насколько это позволяет имеющийся материал) функциональный анализ строения животного, чтобы понять образ его жизни, тип движений, попытаться представить себе наиболее характерные лозы в различных жизненных ситуациях.
Основой функционального анализа строения ископаемых животных стали в первую очередь работы выдающихся палеонтологов прошлого века В. О. Ковалевского и Л. Долло. Как писал А. А. Борисяк [108], метод В. О. Ковалевского состоял в изучении ископаемого скелета в связи с образом жизни животного, которому он принадлежал, т. е. в выявлении экологического значения строения изучаемого скелета (таким путем получались данные для реконструкции животного) Развивая идеи В. О. Ковалевского, Л. Долло создал этологическое направление в палеонтологии, ставшее основой изучения морфологических приспособлений вымерших животных к определенному образу жизни. Следуя этому методу, сам Л. Долло впервые доказал двуногое хождение игуанодонов и обосновал правильный монтаж их скелетов.
Прекрасным примером использования этологического метода может служить проведенный А. А. Борисяком функциональный анализ строения халикотерия Phyllotillon betpakdalensis (принятое ныне название этого вида - Bonssiakia betpakdalens) из третичных отложений Голодной степи в Казахстане [107, 1356]. Исследуя строение зубов халикотерия, А. А. Борисяк пришел к вы воду, что основу питания этого животного составляли листья деревьев, строение его передних конечностей и лопаток свидетельствует, что, добираясь до листьев, халикотерий мог карабкаться передними ногами по стволам деревьев, используя для опоры имевшиеся на пальцах ногти. Такой способ питания делает понятной необычную для легкого в целом скелета относительную тяжеловесность задних конечностей, служивших опорой телу при его вертикальном положении. Следовательно, наиболее вероятной позой кормящегося халикотерия следует считать поставленное вертикально, опирающееся передними конечностями о ствол дерева тело с высоко поднятой вверх и слегка закинутой назад головой (рис. 35).
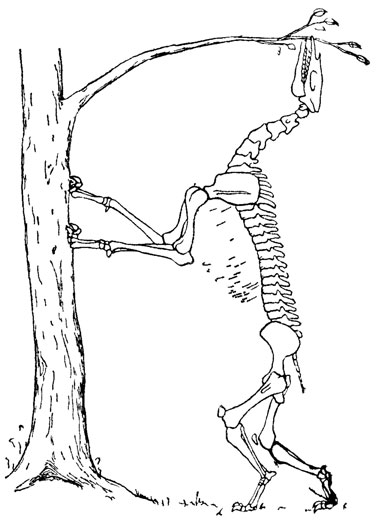
Рис. 35. Поза кормящегося халикотерия. По А. А. Борисяку
Изучение строения вымерших животных позволяет судить о том, какой образ жизни вели интересующие нас виды животных - наземный, водный или амфибиотический. Ископаемые остатки наземных животных могут дать достаточно оснований для суждения о механике их локомоции на определенных субстратах, что также должно учитываться при их изображении. Прямые указания на характер постановки конечностей при локомоции, определяющей возможные аллюры их движений, дают строение костей самих конечностей и их поясов, особенности строения суставов [215, 1801 и др.]. Можно получить и косвенные указания на характер использования различных субстратов вымершими животными. Так, сильное развитие полушарий мозжечка у неогеновой куницы Perutum, о котором можно судить по естественному отливу мозговой капсулы черепа, позволило Ю. А. Орлову [786] предположить, что это животное, несмотря на гигантские размеры, обладало ловкостью и проворством и, видимо, хорошо лазало по деревьям.
Строение парных и непарных плавников может говорить о способах плавания водных животных Исследование конечностей и хвостового плавника ихтиозавров показало, например, ошибочность представлений о том, что эти морские рептилии были хорошими ныряльщиками и могли, подобно дельфинам, выпрыгивать из воды при плавании [787]. Хвостовой плавнику ихтиозавров в отличие от китообразных был расположен в вертикальной плоскости и играл, как у рыб, роль аппарата, толкающего тело вперед, он не был способен в той мере, как у китов, изгибать заднюю часть тела вверх и вниз. Сочленение плечевых костей с поясом передних конечностей у ихтиозавров было таково, что ласты должны были постоянно сохранять перпендикулярное к продольной оси тела положение [787]. Поэтому передние ласты играли роль не гребного механизма, а рулей глубины и аппарата, поддерживающего тело в горизонтальном положении. Отсюда следует, что неверно рисовать ихтиозавров, как это сделали, например, З. Буриан и К. К. Флеров, выскакивающими из воды, с дугообразным изгибом тела; особенности строения непарного и парных плавников говорят скорее о том, что нормально ихтиозавры плавали в полупогруженном состоянии у поверхности моря (рис. 36).
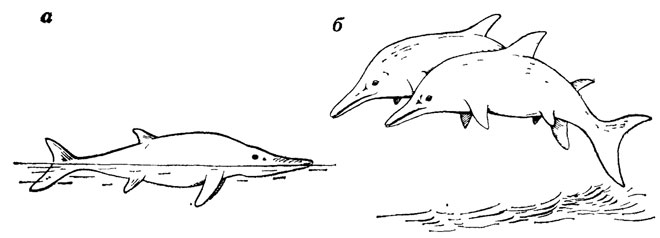
Рис. 36. Изображение раннеюрских ихтиозавров: а - Sfenopterygius (по З. Буриану), б - ихтиозавры из бассейна р. Сены (по Е. Эмихену)
Иллюстрацией того, до какой степени наши представления о внешнем виде и образе жизни вымерших животных могут видоизменяться в зависимости от предположений об их локомоторном аппарате и физиологии, могут служить различные точки зрения по поводу реконструкций некоторых динозавров. До недавнего времени палеонтологи считали большинство динозавров медлительными существами, наиболее крупные из которых вели преимущественно полуводный образ жизни (трудно представить, чтобы животные с огромной массой могли вести наземный образ жизни). Соответствующим образом - полупогруженными в воду обитателями мелководий - их обычно и рисовали. Однако недавно американский палеонтолог Р. Беккер [1296] предположил, что, будучи теплокровными животными, динозавры (включая и самых крупных) вели, подобно некоторым современным копытным, подвижный образ жизни на равнинных участках суши. На рисунках Р. Беккера динозавры приняли совершенно иной облик (рис. 37), несмотря на то что сами черты их строения остались по сути дела традиционными, за исключением более прямой постановки передних конечностей. Реконструкции Р. Беккера не получили широкого одобрения палеонтологов, но мы приводим этот пример для того, чтобы показать, как наше понимание палеонтологии вымерших животных может повлиять на попытки восстановить их внешний облик в условиях среды, которую они населяли.
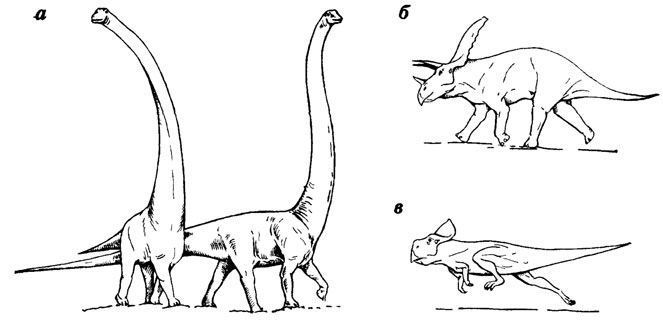
Рис. 37. Реконструкции динозавров. По К. Беккеру. а - Barosaurus; б - Torosaurus; в - Protoceratops
Хорошие примеры того, как по некоторым чертам строения даже отдельных частей тела животных можно воссоздать наиболее вероятные их позы в определенных жизненных ситуациях, дают нам различные группы современных и ископаемых копытных. Все копытные питаются главным образом вегетативными частями растений и совершают большие переходы в поисках новых мест кормежек. Пищевая специализация отдельных видов в конкретных условиях определенных ландшафтов сопряжена с определенными чертами их строения. Для обитающих на равнинах животных-грассифагов необходимо, чтобы опущенная голова свободно доставала до земли. Если травостой в местах обычной пастьбы высокий, то важно, чтобы удлиненной была сама голова, так как животное, опустив голову в траву, не должно лишаться возможности осматриваться. Поэтому если ископаемые копытные имеют длинную (с большим расстоянием от конца морды до глаза) голову, свободно достающую при опускании до земли, то мы можем с большой долей вероятности предположить, что имеем дело со специализированным грассифагом и соответствующим образом изображать его при реконструкции. Иной морфологический тип представляют собой скальные копытные - горал, снежная коза или такин. Все эти звери имеют короткую шею и голову и мощный плечевой пояс - скальный ландшафт дает возможность кормиться, не опуская низко голову, а сильные передние конечности облегчают передвижение по скалам.
Как подобные соображения могут быть учтены при изображении вымерших животных, видно на примере реконструкции шерстистого носорога, выполненной В. Е. Гаруттом [216]. Сравнивая строение шерстистого носорога со строением некоторых современных носорогов, В. Е. Гарутт отметил следующие особенности. Во-первых, шерстистый носорог имел относительно короткие конечности при длинном туловище; сходное строение имеет травоядный белый африканский носорог, в то время как черный африканский носорог, питающийся главным образом ветками кустарников, имеет относительно высокие конечности при более коротком туловище. Во-вторых, у шерстистого носорога плоскость затылка образует с линией основания черепа тупой угол, что определяет опущенное положение головы в спокойном состоянии; у лесных и лесостепных современных носорогов величина этого угла меньше, поэтому в спокойном состоянии их голова поднята выше. В-третьих, на сохранившихся мумифицированных головах шерстистого носорога видно, что верхняя губа у них была широкой, что вообще характерно для копытных, питающихся травянистой растительностью. Наконец, дошедшие до нас рога шерстистого носорога сильно стерты с передней стороны (возможно, в результате использования рогов для разрыхления снега в зимнее время).
Все это позволило предположить, что шерстистый носорог, подобно белому африканскому носорогу, был типично травоядным животным. Исследование сохранившихся пищевых остатков, взятых из ротовой полости сохранившейся мумифицированной головы, подтвердило обоснованность такого предположения. Поэтому изображать шерстистого носорога следует именно пасущимся на открытых равнинах (рис. 38), а не объедающим ветки на кустарниках.

Рис. 38. Пасущийся шерстистый носорог. По В. Е. Гарутту
В ряде случаев указания на образ жизни вымерших животных может давать и сам характер захоронения их скелетов. Так, известны находки скоплений скелетов триасовых капитозавроидных лабиринтодонтов, погибших при пересыхании мелководных водоемов и захороненных на месте их гибели [790]. Такие находки свидетельствуют, по мнению авторов, о том, что эти древние амфибии были чисто водными животными, неспособными покидать воду даже в критических ситуациях. Эти животные не могли, по-видимому, даже переползать из одного водоема в другой; при пересыхании водоемов они искали спасения в наиболее глубоких их участках, где, вероятно, и погибали от перегрева, недостатка кислорода и высыхания.
Однако описание строения вымерших животных, проведение функционального анализа их строения и восстановление характера их образа жизни еще не представляют собой собственно реконструкцию их внешнего облика - они дают только необходимую основу для такой реконструкции [107]. Само изображение внешнего вида создает художник или скульптор, перед которым встают новые проблемы и трудности.
Основная трудность реконструкции внешнего облика вымерших видов связана с тем, что даже в тех случаях, когда палеонтологи дают возможно полное описание строения животного, художник должен изобразить существо, которое он никогда не видел и в принципе увидеть не может. В этом смысле работа художника-реставратора напоминает творчество писателя-фантаста - впрочем, с одной весьма существенной разницей фантастика, даже если она называется "научной", остается фантастикой, а художник должен выполнить изображение, не противоречащее действительно научным представлениям об обитателях Земли далекого прошлого.
Проще всего поступить следующим образом. На основе знания строения скелета можно правильно представить основные пропорции тела животного: если материал дает возможность хоть в какой-то мере представить рельеф мускулатуры в целом (или даже на отдельных частях тела), то художник может с определенной степенью обоснованности "обрастить" скелет мягкими тканями, а затем изобразить получившееся животное, "обтянув" костно-мускульную основу наружными покровами и дополнив картину изображением некоторых деталей строения в соответствии с таксономической принадлежностью реконструируемого животного (зверя "одеть" шерстью, птицу - перьями и т. д.). В результате получается довольно условное изображение, обычно лишенное выразительных черт, свойственных практически всем животным в действительности.
Надо признать, что большинство палеонтологических реконструкций вымерших животных представляют собой схематические изображения костно-мускульной основы, обтянутой кожей и снабженной также весьма условными когтями, копытами, рогами, хоботами и т. д. Именно так, например, выглядят реконструкции некоторых представителей северодвинской фауны, выполненные А. П. Быстровым [125], изображения головы мастодонтов (рис. 39), приведенные в работе А. А. Борисяка [107], и многие другие, не исключая даже считающиеся классическими многочисленные реконструкции З. Буриана. Не меняют обычно сути дела и попытки художников изобразить животных в динамической позе и даже дать изображение "неравных кровавых схваток между хищными и травоядными рептилиями" [125]. При всех стараниях придать реконструкциям максимальную динамичность художнику не всегда удается преодолеть схематизм и условность изображения вымерших животных.
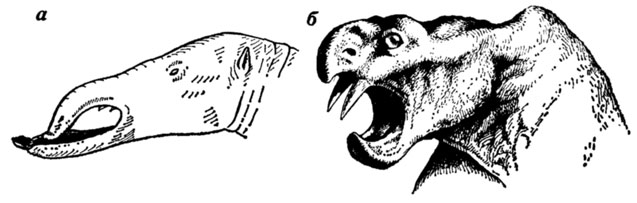
Рис. 39. Изображение головы вымерших животных: а - мастодонта Platybelodon - пример условного изображения (по А. А. Борисяку); б - днцинодонта - изображение по принципу 'обтягивания черепа кожным покровом' (по А. П. Быстрову)
Пытаясь избежать такого схематизма, художники часто уподобляют внешний вид реконструируемых форм облику какого-либо родственного и хорошо известного им современного животного (если это возможно). Именно так часто поступают при изображении вымерших млекопитающих - например, на многочисленных реставрациях вымерших зверей, выполненных зоологом и художником К. К. Флеровым. Флеров был прекрасным художником-анималистом, имевшим богатейший опыт рисования животных с натуры. Вымершие звери на его рисунках всегда изображены в динамичных позах, в жизненно достоверных ситуациях. Однако как это ни парадоксально, именно прекрасное знание современных млекопитающих стало, с нашей точки зрения, причиной того, что многие из изображенных им вымерших зверей имеют облик родственных им (или просто в чем-то похожих на них) современных животных. Так, рисуя миоценового хищника Amphicyon, К. К. Флеров изобразил его очень похожим на крупную массивную собаку (рис. 40, а). Действительно, череп Amphicyon имеет сходство с черепом современных псовых, но его конечности были гораздо более массивными и скорее походили на медвежьи, и он по всей вероятности был, подобно медведям, стопоходящим [1634]; однако последняя особенность строения Amphicyon не получила соответствующего отражения на реконструкции К. К. Флерова. Другой пример: голова протэмболотерия на рисунке К. К. Флерова очень напоминает голову черного африканского носорога (см. рис. 40, б), и только строение выростов на голове указывает на принадлежность нарисованного зверя к титанотериям. Но голова протэмболотерия отличалась от носорожьей не только характером выростов, имевших костную основу: вся она была короче и шире, чем у современных носорогов, ее затылочная часть - более мощной и высокой, а глаза должны были находиться ближе к концу морды.
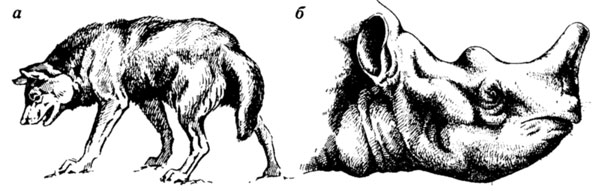
Рис. 40. Реконструкция млекопитающих. По К. К. Флерову из работы Ю. Л. Орлова. а - Amphicyon; б - Protembolo herium
Нам кажется, что выполненные К. К. Флеровым рисунки ископаемых зверей, несмотря на все их несомненные художественные достоинства, не всегда можно считать научно обоснованными реконструкциями в полном смысле этого слова, поскольку работа по восстановлению всех деталей внешнего облика здесь, по сути дела, подменяется подчеркиванием лишь некоторых из них, в то время как остальные "подгоняются" под облик другого животного.
Недостаточную надежность использования при реконструкциях аналогий с другими видами подтверждают и сопоставления облика некоторых близких друг другу современных форм. Даже родственные формы при сходном строении скелетов могут иметь существенные различия в некоторых характерных чертах внешности. Например, все тушканчики - ночные зверьки, у них хорошо развит слух. Соответственно у многих тушканчиков на черепе имеются относительно крупные слуховые барабаны. Но четкой зависимости между размерами слуховых барабанов на черепе и размерами наружного уха у тушканчиков нет. Такие виды, как гребнепалый тушканчик Paradipus ctenodactylus или длинноухий тушканчик Euchoretus naso, при крупных слуховых барабанах имеют и крупные ушные раковины, а у карликовых тушканчиков при очень крупных слуховых барабанах ушные раковины, наоборот, очень маленькие (рис. 41), что придает их голове совершенно иной облик.

Рис. 41. Величина и форма слуховых барабанов (показаны черным на контурах черепов) и наружного уха у различных тушканчиков Рис. В. М. Смирина. а - мохнатоногого Dipus sagitta, б - длинноухого Euchoretus naso в - гребнепалого Paradipus ctenodactylus, г - тушканчика Лихтенштейна Eremodipus lichtensteini, д - жирнохвостого карликового Salpingotus crassicauda
Другой пример - строение черепа и головы некоторых зубастых китов. Кости переднего отдела черепа этих животных образуют длинное рыло, которое в той или иной степени определяет и внешний вид головы большинства представителей. Но у кашалота в передней части головы имеется мощный спермацетовый мешок, выступающий за передний конец челюстей и придающий голове огромные размеры и слегка сплюснутую с боков бочкообразную форму без каких бы то ни было признаков удлиненного рыла (рис. 42). И хотя кости черепа образуют "ложе" для основания этого мешка, мы все же думаем, что человек, видевший только череп кашалота и знающий облик других зубастых китов, никогда бы не реконструировал спермацетовый мешок такого гигантского размера, какой он имеет в действительности.
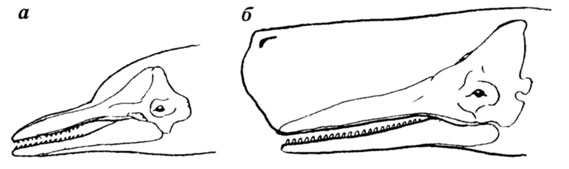
Рис. 42. Контуры черепа и головы зубастых китов Рис. В. М. Смирина. а - дельфина б - кашалота
Особые трудности могут возникать при попытках изображения вымерших видов, которые обладали очень своеобразными чертами строения, не имеющими прямых аналогий в современной фауне. Ни у кого из современных пресмыкающих нет, например, таких гигантских остистых отростков на позвонках, как у пермских эдафозавров; никто не имеет на спине рядов таких длинных чешуи, какие были у триасовой рептилии Podopteryx mirabilis, или костных щитов, как у стегозавров. У миоценовой свиньи Kubanochoerits лобные кости образовывали большой непарный вырост, лишенный, вероятно, рогового чехла [787]. По поводу функций таких образований иногда высказывают различные точки зрения. Вместе с тем, понимание их функций должно определить характер изображения животного, обладавшего столь необычным строением. Были ли гигантские остистые отростки и костные щиты у упомянутых рептилий погружены в мягкие ткани или же выходили за их пределы? Ответить на такие вопросы должны палеонтологи, а художники уже в соответствии с этими ответами должны дать обоснованные изображения внешнего вида животных.
Однако главная и едва ли вообще преодолимая трудность при создании образно выразительных реконструкций связана с тем, что в формировании внешнего облика животных очень часто решающую роль играют такие черты строения, которые вообще не оставляют никаких документальных свидетельств на ископаемом материале. Как справедливо указывает Дж. Г. Симпсон [953], о форме таких образований, как ушные раковины или хоботы у зверей, мы можем только гадать, на ископаемом материале очень редко сохраняются следы наружных покровов. Не всегда можно дать обоснованную реконструкцию таких образований, как когти, копыта, рога или клювы, от которых, как правило, сохраняется только внутренняя костная основа, а такие черты внешнего облика, как наличие гривы, окраска животного, присутствие в ней пятен или полос в большинстве случаев абсолютно невосстановимы [953]. У некоторых современных зверей (ежей, тенреков, дикобразов) на теле имеются иглы различной длины и окраски, можем ли мы с уверенностью судить о наличии таких образований у вымерших зверей? Не надо быть специалистом, чтобы оценить роль всех перечисленных черт строения в формировании внешнего облика животных - тигр без полос уже не будет тигром, лев без гривы - львом, а дикобраз без игл - дикобразом. Хотим мы того или не хотим, но мы должны признать, что единственными "ориентирами" при изображении некоторых деталей внешнего вида вымерших животных иногда оказываются фантазия художника, его художественный вкус и интуиция.
Не надо думать что попытки воссоздать характерные детали внешнего облика вымерших животных вообще лишены смысла. В каждом отдельном случае надо отчетливо понимать, что можно действительно восстановить, а о чем - только догадываться. При этом следует иметь в виду, что у представителей разных групп животных соотношение восстановимых и невосстановимых черт во внешнем облике могло быть неодинаковым. В наиболее трудном положении художник-реставратор часто оказывается при изображении зверей, большинство которых имеет сплошной волосяной покров, или покрытых перьями птиц. С другой стороны, покровные костные образования, развивающиеся на теле у некоторых низших позвоночных, могут облегчить работу художника, поскольку такие образования, в значительной степени определявшие внешние очертания этих животных, иногда хорошо сохраняются на ископаемом материале (наиболее типичный случай таких реконструкций - воссоздание по отпечаткам облика костистых рыб).
При достаточной полноте ископаемого материала палеонтолог может дать описание строения вымерших животных и провести функциональный анализ их организации. На основании этих данных художник имеет возможность, не рискуя впасть в серьезные ошибки, представить контуры и пропорции тела этих животных, пределы их возможных движений, некоторые характерные позы. В то же время он всегда должен помнить, что о некоторых чертах облика ископаемых животных никаких документальных свидетельств не остается и что именно такие черты во многом определяли выразительность образа этих животных. Поэтому в художественных реконструкциях вымерших видов неизбежно присутствует и известный элемент фантазии, и одна из главных задач авторов реконструкций - не допустить противоречия предлагаемого изображения с имеющимся научным представлением.
Мы считаем, что соотношение действительно восстановленных и только предполагаемых черт внешнего облика в изображениях несуществующих ныне животных может в известной мере меняться в зависимости от тех непосредственных задач, которые стоят перед авторами палеонтологических реконструкций. В научной иллюстрации элемент художественного вымысла должен быть, безусловно, сведен к возможному минимуму. Главная цель таких иллюстраций - дать наглядное представление о строении вымерших животных и об основных особенностях их образа жизни, и здесь, вероятно, наиболее уместны несколько упрощенные и условные изображения, показывающие лишь наиболее общие и характерные черты их организации и поведения. При этом желательно полностью отказаться от соблазна "оживить" изображения придуманными деталями внешнего облика. Удачным примером подобного подхода нам представляются иллюстрации художницы Д. Спенсер к монографии Дж. Г. Симпсона [953], выполненные в предельно условной манере.
В иллюстрациях научно-популярных работ и рисунках для музейных экспозиций, рассчитанных на широкий круг интересующихся естественной историей людей, художникам не следует пренебрегать попытками придать реконструкциям определенную образную выразительность. Однако при выполнении таких работ грамотность художника приобретает особое значение, поскольку именно в музеях и из популярной литературы неспециалисты узнают историю жизни на Земле. Излишняя свобода творческого воображения может стать здесь источником ошибочных представлений об эволюции животного мира.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'