
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
1.5. Изучение онтогенеза вымерших организмов (В. П. Макридин, М. А. Шишкин)
Рассматриваемую в данной главе область исследований мы обозначаем условным термином "палеоэмбриология", понимая под ним изучение индивидуального развития в той мере, в которой оно запечатлевается на палеонтологическом материале. Хотя изолированные остатки собственно эмбриональных стадий различных организмов или даже предшествующих им продуктов гаметогенеза (в особенности яйцевые оболочки животных, споры, пыльца и семена растений) встречаются весьма часто, сами по себе они лишь в небольшой степени используются для характеристики онтогенеза древних форм. Основная же роль в этом плане принадлежит остаткам, показывающим ход постэмбриональных скелетных преобразований, ведущих в итоге к становлению взрослой особи.
Уже с начальных этапов развития палеонтологии онтогенетические исследования стали ее неотъемлемой составной частью, приобретя важную роль при выяснении филетических отношений между формами, а также при анализе происхождения различных морфологических структур. Начиная с работ Л. Вюртембергера [2506] и А. Хайетта [1774, 1776], введших понятия акселерации и ретардации (соответственно для ускорения и замедления онтогенетического развития признаков), палеоэмбриология стала уникальным источником сведений о гетерохрониях (эволюционных изменениях темпов индивидуального развития) и рекапитуляциях (повторениях в онтогенезе потомка анцестральных черт), вскрывая пути их исторического становления.
Поскольку жизненные циклы организмов очень разнообразны и часто совершенно несравнимы, крайне трудно дать определение понятия индивидуального развития (онтогенеза) и установить границы его применения. Например, оспаривалось существование онтогенеза у простейших - в том смысле, в каком он существует у многоклеточных [926]. Как особый процесс выделяется развитие на основе вегетативного размножения - бластогенез, или гистерогенез [1738], с которым палеонтолог чаще всего сталкивается при изучении колониальных организмов (губок, кораллов, мшанок, граптолитов и др.); при этом формирование индивидуальной колонии обозначается как астогенез [1472]. Полный цикл развития, ведущий к повторению исходной организации, может быть связан также с чередованием полового и бесполого (бесполых) поколений (метагенез).
Однако в практике палеонтологических исследований эти разграничения обычно не связаны с какими-либо существенными различиями в подходе к изучению онтогенетических процессов. Далее, приложимость понятия онтогенеза к развитию простейших (включающему и такие явления, как, например, метаморфоз) сейчас не вызывает сомнений [428]. Кроме того, независимо от споров па этому вопросу, наличие онтогенетических изменений (прибавление новых камер) у таких скелетообразующих Protozoa, как фораминиферы, всегда было очевидным фактом. Сопоставление разных типов развития, по существу, проводится здесь так же, как, например, в отношении аммонитов, и аналогичным образом служит основой для выводов о гетерохрониях и рекапитуляциях. Не возникает также существенных различий в подходе к изучению развития половых и бесполых поколений (например, микро- и макросферических генераций у фораминифер, или прото- и гистерокораллитов у колониальных кораллов). Изучение астогенеза используется палеонтологами для выводов о модусах исторических преобразований развития в той же мере, что и развитие индивидуальных особей.
Этапность индивидуального развития также является относительным понятием. Ее невозможно свести к одной общей схеме даже для наиболее типичного случая, т. е. развития из оплодотворенной яйцеклетки, ведущего непосредственно к формированию половозрелого одиночного организма. Схема А. Н. Северцова [926], подразделявшего жизнь многоклеточных на периоды онтогенеза и зрелости, а период онтогенеза в свою очередь - на периоды морфогенеза и преимущественного роста, далеко не отражает всего многообразия существующих здесь соотношений. Процессы морфогенеза и роста, относительно хорошо разграниченные у позвоночных, во многих других группах протекают в значительной мере параллельно, а у насекомых с полным превращением период роста (личиночные стадии) заключен между покоящимися стадиями с интенсивным морфогенезом (яйцо и куколка). Изменения скелетных структур, происходящие в репродуктивном и даже старческом возрасте, могут иметь большое значение для понимания путей эволюции и родственных связей отдельных форм, и потому их изучение на ископаемом материале столь же важно, как и анализ более ранних стадий развития.
При изучении онтогенеза ископаемых беспозвоночных чаще всего используют расчленение этого процесса на стадии, выделенные А. Хайеттом для головоногих моллюсков [1774]: 1) эмбриональную, личиночную, или непионическую (от греч. nepion - младенец); 2) юношескую, или неаническую (от греч. neanis - юный); 3) взрослую, или эфебастическую (от греч. ephebus - зрелый); 4) старческую, или геронтическую (от греч. geros - старость). Варианты этой схемы используют также при описании развития ископаемых фораминифер, кораллов, брахиопод и др. Для отдельных групп применяют более специфические подразделения - например, протасписная, мерасписная и голасписная стадии (этапы формирования отделов тела) в постэмбриональном развитии трилобитов.
Основной объект палеонтологического изучения онтогенеза - постэмбриональное развитие. Как и у современных животных, оно могло быть прямым (когда организм, покидающий яйцо, отличается от взрослого в основном лишь размерами) или же протекать с метаморфозом, т. е. через ряд существенно различающихся личиночных состояний. Метаморфоз в свою очередь может быть эволютивным, т. е. основанным на относительно постепенных морфогенетических изменениях (например, развитие у большинства рыб), и катастрофическим, связанным с резкими преобразованиями - как у бесхвостых амфибий, насекомых с полным превращением или большинства сидячих бентосных беспозвоночных, имеющих расселительную личинку. Границы между этими категориями также достаточно условны, поскольку практически ни один тип постэмбрионального развития не может быть сведен исключительно к росту, и в нем всегда можно найти структурные преобразования, т. е. элементы "метаморфоза".
Резкие онтогенетические превращения, не имеющие промежуточных стадий, редко и очень неполно запечатлеваются в геологической летописи, будучи связаны обычно с ранними фазами развития, предшествующими скелетообразованию. Поэтому основным предметом онтогенетических исследований на ископаемом материале являются, по существу, процессы эволютивного метаморфоза скелетных структур. Это остается в общем справедливым, независимо от того, изучаем ли мы изменения, основанные на аккреционном росте (раковинные организмы, кораллы и т. п.), прибавления новых элементов (иглокожие), линьке (трилобиты, остракоды) или морфологически более сложных преобразованиях. Резкий (катастрофический) метаморфоз чаще всего бывает доступен для изучения у насекомых (см., например [458, 459], где у ископаемых комаров описаны стадии развития - яйцо, личинка, куколка и имаго обоих полов). Личинки в сочетании с имаго известны у многих поденок, веснянок, жуков, вислокрылых и т. д. [830, 831].
Поскольку возможности палеонтологического изучения индивидуального развития ограничены лишь морфогенезом скелета, то иногда полагают, что вообще неправомерно говорить об изучении онтогенеза [2227]. Это не вполне верно. Во-первых, анализ скелетных возрастных изменений может давать информацию об одновременных морфологических и функциональных преобразованиях различных "мягких" структур вымершего организма. Например, микроструктура стенки протоконха аммонитов указывает на отсутствие (или неактивное состояние) мантии в фазе развития [53]; в развитии древних амфибий-лабиринтодонтов строение жаберно-челюстного аппарата позволяет проследить становление полости среднего уха и ряда кровеносных сосудов [1176, 1392]. С другой стороны, любое исследование по развитию рецентных форм точно так же ограничивается анализом изменений конкретных структур или их комплексов, как и исследование ископаемого материала. Возможности охвата различных аспектов развития здесь, конечно, неизмеримо больше, но это отличие не носит принципиального характера. В конечном счете наше знание целостного онтогенеза складывается из изучения морфогенезов отдельных органов и признаков [423].
Источники онтогенетической информации разнообразны и могут быть подразделены на несколько групп.
Яйцевые оболочки. В последние годы благодаря совершенствованию методики дезинтеграции пород, содержащих фоссильные микроскопические остатки организмов, значительно возросло числа находок ископаемых яиц беспозвоночных. Так, описаны известковые яйца наземной улитки Wallonia из плиоцена Канзаса (США), сходные с кальцитизированными яйцами различных Stylomatophora, ранее обнаруженными в олигоцене Англии и миоцене Уганды [2389]; скопления яиц морских гастропод найдены на отпечатке внутренней поверхности створки раковины Pseudopecten из нижней юры ФРГ [1812]. Следует заметить, что и современные гастроподы нередко используют пустые раковины пектинид для откладывания яиц. Известны находки яиц раннепенсильванской бластоидеи Pentremites rusticus, представляющие собой мелкие (0,12-0,20 мм) полупрозрачные сферы, в большом количестве расположенные в пределах задних (анальных) гидроспир, которые, по-видимому, были приспособлены для вынашивания яиц вместо обычной респираторной функции [1816]. Кроме того, была обнаружена "сумка с яйцами" - предположительно яичники самок макроконхов среднетриасового головоногого моллюска Ceratites evolutus parabolicus и раннетоарского Eleganticeras elegantulum [1986].
Многочисленны также находки ископаемых яиц членистоногих: щитней, конхострак, насекомых и др.
Значительно более широкий и лучше изученный фактический материал накоплен по яйцевым структурам позвоночных животных [203, 209, 1986]. Среди хрящевых рыб описаны ископаемые яйцевые капсулы эласмобранхий и химер. Первые известны с девона [1984, 2325], а вторые - с триаса [175]. Остатки яйцевых капсул химер встречены в юрских, верхнемеловых и олигоценовых отложениях Западной Европы, Советского Союза и Северной Америки [763].
Древнейшая находка яйца рептилии приурочена к нижней перми США [2175], но ее природа дискуссионна [1740]. Гораздо чаще остатки яиц рептилий, принадлежащие в основном динозаврам и черепахам, встречаются в мезозойских отложениях, где они представлены фрагментами скорлупы, отдельными целыми яйцами и их кладками. Яйца черепах встречены начиная с альба [1956, 1250, 1418], но наиболее широко их остатки, как и остатки яиц динозавров, распространены в верхнемеловых отложениях МНР [1003, 1250, 1275], СССР, КНР, Франции, США, Бразилии и других стран. Иногда наряду с известковой скорлупой яйца сохраняется и подстилающая мембрана, состоящая из устойчивых протеиновых волокон [1004]. Особый интерес представляют единичные находки яиц с остатками эмбрионов. Так, яйцо из верхнего мела МНР, принадлежащее, по-видимому, рогатому динозавру, содержит остатки скелета, степень окостенения которого примерно соответствует последней стадии инкубационного периода у современных птиц [1005]. Точная идентификация остатков яиц (отнесение их к конкретным формам) обычно затрудняется редкостью совместного захоронения. К подобным редким исключениям относится находка кладки в верхнемеловых отложениях шт. Монтана (США), содержащей наряду со скорлупой яиц остатки ювенильных особей гадрозавра Maisaura peeblesorum разных возрастных стадий [1755].
Наиболее древние ископаемые яйца птиц известны в нижнем эоцене Франции. Их остатки довольно часто встречаются в различных вышележащих горизонтах кайнозоя СССР (Туркмении, Молдавии, Забайкалья), МНР и КНР. Преобладают яйца страусов и других крупных бескилевых форм, но описаны также находки, относящиеся к летающим птицам, включая и гнездовые кладки [203, 1545].
Изучение яйцевых оболочек позвоночных пока слабо связано с собственно онтогенетическими исследованиями и в основном служит для выделения различных типов гистологической структуры, имеющих систематическое и стратиграфическое значение [502, 901, 1003, 1004, 1508].
С этой категорией остатков можно сблизить и изолированные чехлы ранних личиночных стадий, существовавшие у отдельных организмов. Например, в полостях гелиолитоидей из силура Подолии встречены известковые образования, напоминающие капсулы личинок (планул) современных кораллов [274].
Онтогенетические серии. В тех случаях, когда онтогенетические изменения скелета не сводятся к простому приращению новых слоев или добавлению элементов, охватывающих или надстраивающих его прежнюю структуру, ход индивидуального развития не может быть восстановлен по строению одной особи - для этого требуется подбор возрастных серий. Так обстоит дело, например, с развитием хитинового скелета членистоногих, обновляющегося путем многократных линек, с целостными преобразованиями черепа позвоночных, или же с развитием ручного аппарата брахиопод (хотя их раковина изменяется путем аккреции).
Сама процедура подбора онтогенетических серий имеет свои трудности, поскольку в качестве возрастных особенностей могут быть ошибочно интерпретированы отличия другой природы, например систематические или половые. Поэтому всегда предпочтительно наличие массового материала по отдельным стадиям, позволяющего выявить типичную усредненную картину развития. Возможность ошибок, связанных со смешением таксонов, уменьшается в случае совместных находок зрелых экземпляров вида и его предполагаемых ювенильных стадий; см., например [2064].
Изучение онтогенеза по возрастным сериям при наличии достаточного материала применяют в отношении самых различных групп организмов, независимо от характера роста их скелета. В частности, им широко затрагиваются и формы с аккреционным или аддитивным ростом (например, моллюски, брахиоподы и иглокожие). Один из типичных объектов таких исследований среди беспозвоночных - организмы с периодической сменой наружного скелета (членистоногие). Это в особенности относится к изучению трилобитов, у которых анализ возрастных серий позволяет выявить их характерные онтогенетические стадии (протасписную - мерасписную - голасписную) совместно с более дробными подразделениями [1424, 1758, 1759, 1906, 2487]; иногда сохраняется и более ранняя стадия личиночного развития - фазелус, соответствующая переходу от науплиуса к зоэа у высших ракообразных [1595].
Рассматриваемый метод является также основным для изучения развития панциря остракод [496, 1321, 1722] и скелета насекомых, у которых преобладающим объектом онтогенетических исследований служат различные нимфальные и субимагинальные стадии форм с неполным превращением [686, 955, 956, 1140, 1852, 2503]. Среди других групп беспозвоночных метод возрастных серий особенно широко применяют при исследованиях ручного аппарата брахиопод [306, 967, 969 и др.].
Изучение относительно ранних морфогенетических процессов у позвоночных с помощью возрастных серий возможно на примерах рыб и амфибий, развитие которых включает личиночные стадии. Особенно важны в этом отношении данные по позднекаменноугольным и раннепермским амфибиям - рахитомным лабиринтодонтам и батрахозаврам, у которых доступные изучению онтогенетические ряды иногда начинаются со стадий, предшествующих первым проявлениям окостенения [434, 1200, 1367, 1463, 1735]. На этом материале удается проследить многие аспекты развития: порядок появления отдельных окостенений, преобразования жаберного аппарата, покровного орнамента и т. д. Аналогичные исследования проводились на палеогеновых и неогеновых бесхвостых амфибиях; в ряде случаев удалось достаточно полно проследить процесс метаморфоза [2310, 2311].
У амниот, обладающих прямым развитием, эмбриональные окостенения, сохранившиеся внутри яиц, описаны лишь в немногих случаях - например, у меловых динозавров [1005] и птиц [1552]. Здесь первоочередной проблемой является не изучение развития, а идентификация самих остатков. Большинство же данных по онтогенезу ископаемых амниот касается позднего развития, когда основная часть морфогенетических преобразований уже завершена и дальнейшие изменения связаны преимущественно с аллометрическим ростом и усилением окостенения. Примерами в данном случае могут служить развитие позднемеловых динозавров Protoceratops, для которых известен весьма полный ряд ювенильных стадий [1456], юрских птерозавров - птеродактилей и рамфоринхов [2474, 2475] и водных ринхозавров - плеврозаврид [1568].
Кроме непосредственного изучения морфогенеза конкретных форм с помощью возрастных стадий у позвоночных возможно и косвенное моделирование обобщенных типов развития для более крупных групп на основе сопоставления близких взрослых форм с различной степенью дефинитивного недоразвития (фетализации). Применение этого метода к изучению пермских и триасовых амфибий-лабиринтодонтов позволило восстановить в общих чертах картину онтогенеза покровного орнамента, эндокрания, среднего уха (включая и эволюцию его развития), а также некоторых элементов сосудистой системы [1176]. Недостатком данного метода, помимо обобщенности построений, являются определенные трудности в различении генерализованных особенностей развития группы и частных черт, свойственных лишь отдельным ее формам. Преимущество же заключается в том, что метод позволяет в ряде случаев анализировать развитие таких структур, которые на остатках настоящих личинок либо не окостеневают, либо не оставляют следов (например, хрящевые связи эндокрания и висцерального скелета, челюстные сосуды и т. д.).
Детали строения, развитие которых может исследоваться на ископаемом материале с помощью возрастных стадий, охватывают широкий круг признаков. У беспозвоночных, помимо черт, названных выше, а также изменений формы, размеров, наружной скульптуры и других, следует отметить такие исследуемые в динамике внутренние структуры, как замок раковин двустворчатых моллюсков [64, 730, 731 и др.], отпечатки аддуктора на раковинах остракод [268], замочный отросток брахиопод [558, 605, 913 и др.] и т. п. Важную роль в этих исследованиях играет сравнение с аналогичными преобразованиями у рецентных форм.
Организмы с аккреционным ростом скелета. В случае когда скелет изменяется в онтогенезе за счет приращения новых частей без резорбации старых, анализ его последовательных генераций у одной особи позволяет судить о ходе ее развития. Такой тип скелетообразования в той или иной мере свойствен большинству групп беспозвоночных, остатки которых широко известны в ископаемом состоянии - фораминиферам, археоциатам, кораллам, моллюскам, брахиоподам, а в отдельных случаях - членистоногим (конхостракам) и др. Для многих из этих групп, таких, как археоциаты и кораллы, где изучение отдельных видов ведется с помощью поперечных срезов скелета на разных уровнях, оно практически равнозначно изучению их онтогенеза (рис. 44).
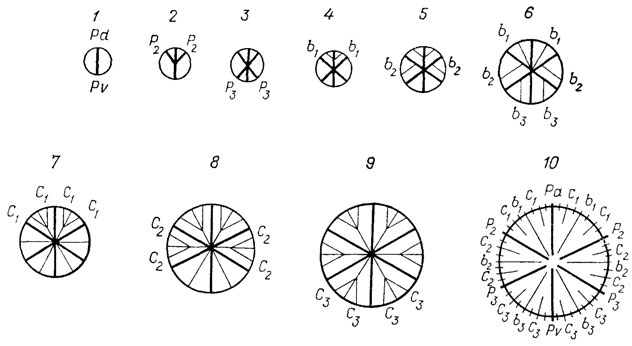
Рис. 44. Онтогенез септального аппарата у склерактинии Turbinolia sulcata Larru Франция, Парижский басе, ср. эоцен. По Ж. Аллуато. 1 - первоначально возникает одна осевая (Pd-Pv) дорзо-вентральная протосепта, определяющая двустороннюю симметрию полипа, 2 - далее появляются две дорзальные протосепты (Р2) и 3 - две вентральные протосепты (Р3), 4 - появляются две дорзальные септы второго порядка (b1), 5 - еще две септы второго порядка (b2) и 6 - последние две септы второго порядка (b3), 7 - появляются четыре септы третьего порядка на стороне дорзальных камер (C1), 8 - еще четыре септы третьего порядка, со стороны боковых камер (С2) и 9 - последние четыре септы третьего порядка, 10 - септы третьего порядка во всех секторах отходят от септ второго порядка и выпрямляются, образуя 24 одинаковых сектора, одновременно возникают еще 24 слабо развитых (рудиментарных) септы четвертого порядка, расположенные равномерно во всех секторах
Онтогенетический метод в палеонтологии возник именно в связи с анализом аккреционных изменений скелета - в первую очередь у головоногих моллюсков (рис. 45). В то же время наряду с исследованием онтогенеза скелета по характеру его нарастания для большинства подобных групп широко практикуется и исследование возрастных стадий.
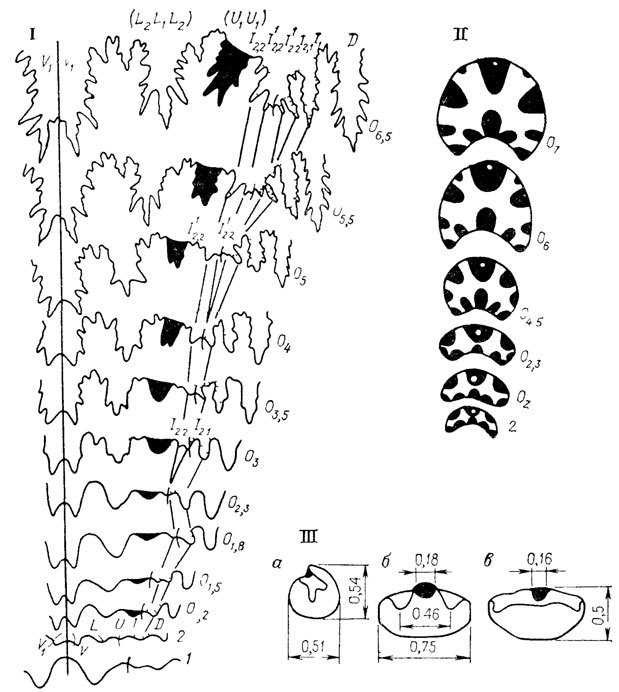
Рис. 45. Онтогенез раковины аммонита Pavlovia iatriensis Ilov из волжского яруса Приполярного Урала. По Т. Л. Ломинадзе, И. В. Кванталиани, М. З. Шарикадзе I - лопастная линия, 1, 2 - 1-я и 2-я лопастные линии, О1,2, О1,5, О2, О6,5 - положение лопастной линии или сечения на обороте раковины L - боковая (латеральная), U - пупковая (умбиликальная), V - брюшная (вентральная), D - спинная (дорсальная), I - внутренняя (внутренняя боковая), II - форма поперечного сечения оборотов раковины, III - протоконх, а - вид сбоку, ув 14, б - вид сверху, в - вид со стороны просепты, ув 20
Количество работ о процессе преобразования скелета у форм с аккреционным ростом очень велико: по фораминиферам - [59, 260, 262, 626-628]; по археоциатам - [371, 873] - по одиночным кораллам-ругозам - [177, 431, 438, 439, 440, 917, 2004, 2103]; по одиночным склерактиниям - [71, 546, 1314, 1315, 1471]; по аммоноидеям - [90, 91, 131, 348, 405, 534, 671, 696, 699, 700, 702, 704, 716, 890, 891, 1160, 1161, 2150, 2512] и многие другие.
Скелетные чехлы, растущие аккреционным путем, часто сохраняют в своей вершинной (начальной) части личиночные или эмбриональные структуры: известковую капсулу личинки (планулы), с которой начинается рост протокораллита у различных кораллов - гелиолитид, ругоз, склерактиний; начальную раковину моллюсков-протоконх гастропод и цефалопод или продиссоконх двустворок; аналогичное образование (протегул) у брахиопод и т. д. У большинства моллюсков эта раковина соответствует стадии свободной личинки - велигера; у головоногих моллюсков ее роль служит предметом многих дискуссий. Все современные головоногие характеризуются прямым развитием, но в отношении ряда ископаемых форм допускают существование вторичной или первичной личиночной стадии - в частности, для ряда наутилоидей, бактритоидей и аммонитов [53, 893, 1167, 1537, 1558]. Так, у бактритов и аммонитов, где вздутый протоконх представляет собой гидростатическую камеру раковинки юной особи, которая вела планктонный образ жизни, последнюю иногда считают вторичной личинкой [53, 1853]. При решении подобных вопросов большую роль играет метод электронной микроскопии, позволяющий по гистологическому строению первичной раковины установить характер изменения ее секреции при переходе к дальнейшим стадиям развития [345, 349, 350, 705, 1338].
Явления аккреционного роста делают возможным широкое изучение онтогенетических изменений поверхностных структур скелета, маркируемых линиями нарастания. Таким путем исследуются, например, возрастные изменения размеров и формы раковин моллюсков и брахиопод [605, 1459, 1765]. Изучение вариксов на раковинах многих видов гастропод позволяет судить о динамике роста этих моллюсков [1886]. Развитие их шло очень быстро в периоды, соответствующие промежуткам между вариксами, хотя в это время и происходили более мелкие колебания скорости роста раковин.
Многочисленные исследования проведены на эпитеке кораллов (ругоз и склерактиний), состоящей из ритмично чередующихся горизонтальных нитевидных колец различной толщины, образование которых связано с неравномерным поглощением извести из морской воды, обусловленным различными (суточными, сезонными и др.) биологическими ритмами [551, 552, 1657, 1658, 2257]. Это позволяет определить по характерным линиям прирастания на эпитеке (ругам) темпы роста кораллов. Радиографически показано [1589], что зоны роста на эпитеке представлены парами тонких слоев - темного и светлого, различающихся по плотности скелетного вещества и указывающих на соответствующие сезонные изменения в развитии кораллов. Экспериментально доказано, что темные слои соответствуют летним, а светлые - зимним периодам [827]. Определенные аналогии в ритмическом развитии эпитеки одиночных кораллов и чередовании темных и светлых полос известны в астогенезе колониальных форм [551, 552, 1589].
Близок к аккреции рост скелета путем добавления однородных частей, сохраняющих самостоятельность. Такой тип роста, который может быть назван аддитивным, свойствен, например, иглокожим. У морских ежей индивидуальное развитие на постларвальной стадии связано с постепенным увеличением числа табличек в амбулакральных и интерамбулакральных полях за счет появления новых элементов у амбулакрального поля и последующего отодвигания их вниз. При этом сами таблички растут аккреционно.
У позвоночных животных рост скелетной ткани путем периодического приращения новых слоев часто сопровождается более или менее интенсивной резорбцией и перестройкой старых генераций. Тем не менее во многих случаях последовательность наслоений может быть с успехом использована для определения индивидуального возраста особи и анализа динамики ее роста. У костистых рыб для этих целей используются кольца нарастания чешуи, а также структура отолитов, позвонков и покровных костей [766]. Закономерности роста и дифференциации скелетных тканей ископаемых позвоночных (гистогенеза), обычно изучают на срезах. Эти исследования составляют предмет палеогистологии (см. 2.4). Иногда, однако, линии прирастания хорошо видны на поверхности костей - например, на чешуях костистых рыб или на щитках панциря некоторых бесчелюстных из группы Heterostaci [766], что позволяет изучать возрастные изменения формы этих элементов. Еще одним примером подобных изменений, наблюдаемых на поверхности костей, является развитие ячей или бугорков наружного орнамента по периферии центров окостенения покровных элементов у ископаемых амфибий по мере их роста [1176, 2204].
Колонии. Как более сложный вариант аккреции можно рассматривать рост колониальных организмов (кораллов, мшанок, граптолитов и др.). В этом случае речь идет о приращении скелета не одной особи (хотя оно также имеет место), а у целых их генераций, образующих интегрированное надиндивидуальное объединение, имеющее свой собственный жизненный цикл - астогенез (см. 1.2). Развитие одной вегетативной особи в колонии, как уже было сказано, называют бластогенезом, или гистерогенезом. Его отличие от типичного онтогенеза заключается в начальной стадии, представляющей собой не яйцеклетку, а группу разнородных дифференцированных клеток материнской особи [429].
Изучение бластогенеза у колониальных организмов с прирастающим скелетом, например у кораллов [98, 101, 547, 1076, 1807, 2187, 2320 и др.], принципиально не отличается от изучения индивидуального развития особи-основательницы колонии, возникшей половым путем, а также одиночных форм с аналогичным типом скелетообразования (рис. 46).
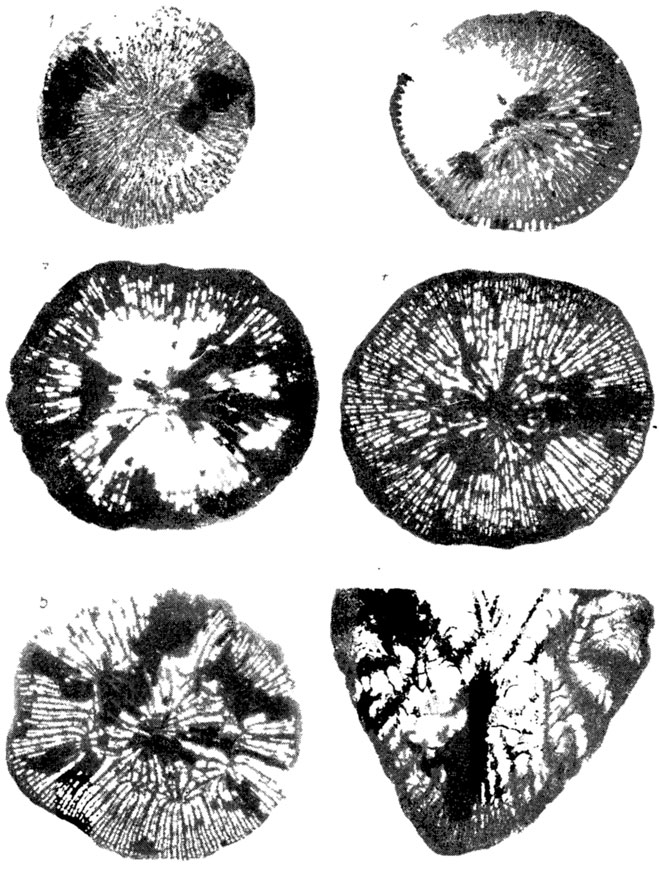
Рис. 46. Астогенез колонии склерактинии Monocyclastraea alpina (Koby). Крымская область, Белогорский р-н, с. Балки; нижний берриас. По Е. И. Кузьмичевой. 1 - стадия одиночного кораллита, ув. 4; 2 - в 10 мм от центра появляются три дочерние почки, две из них по одну сторону чашечной ложбины и одна - по другую, ув. 4; 3 - ранее появившиеся кораллиты отчетливо обособлены друг от друга, прослеживаются разрывы септ и возникновение новых дочерних почек, ув. 2; 4 - видны шесть четко обособленных дочерних кораллитов и появление по обеим сторонам материнской чашки еще двух новых почек, ув. 2; 5 - вокруг материнского кораллита расположены девять сформировавшихся дочерних кораллитов, ув. 2; 6 - продольное сечение взрослой колонии, ув. 5
Астогенез представляет собой более своеобразный процесс. Его изучение началось давно [1579, 2024], и нередко его описывали под обобщенным названием "онтогенез" [321, 409, 981, 1007]. Более направленно астогенез исследуют у кораллов и мшанок в последние десятилетия [98, 101, 102, 176, 711, 918, 1001, 1075, 1076, 1345, 1528, 1575, 1939, 1940, 2077, 2095 и др.].
Астогенез не есть просто сумма индивидуальных циклов особей, составляющих колонию; он имеет более общие свойства. Начало астогенеза - появление и развитие особи-основательницы: у кораллов - протокоралла (или протокораллита); у мшанок - анцеструлы; у граптолитов - сикулы и т. д. Эту стадию астогенеза называют филастической [1472]. Далее у кораллов следуют брефастическая (детская), неанастическая (юношеская), эфебастическая (зрелая) и, наконец, геронтастическая (старческая) стадии. Брефастическая стадия характеризуется бесполым размножением протокораллита, в неанастической стадии происходит размножение бесполым путем следующих генераций с эфебастической стадией связано появление полового размножения.
Общим для астогенеза кораллов (ругоз, гелиолитид и склерактиний) является неравномерность темпов развития более быстрый рост колонии и интенсивное вегетативное размножение на ранних стадиях сменяются нарастанием дифференциации кораллитов, вплоть до полиморфизма к концу развития, наличие на эфебастической стадии так называемого цикломорфоза, т. е. чередования фаз полового и бесполого размножения, которым соответствуют в колонии смены темных и светлых зон и ритмические изменения морфометрических показателей [102]. Помимо сезонного чередования типов размножения [840, 1310 и др.] причинами цикломорфоза могут быть и другие физиологические особенности колоний. Вместе с тем ритмические изменения, аналогичные цикломорфозу, отмечены и в бластогенезе [98].
Астогенез сходен с индивидуальным развитием тем, что заключает в себе рекапитуляции прежних состояний колонии, соответствующие более ранним этапа эволюции данной группы. Это было показано как для современных кишечнополостных [728], так и для ископаемых [92, 97, 101, 105, 200, 548, 550, 744 и др.].
Естественно, что изучение астогенеза желательно проводить на одной колонии. Но практически это не всегда возможно, и возникает необходимость в подборе серии колоний, позволяющей получить представление о всей последовательности возрастных изменений данного вида.
Сравнительно-астогенетических наблюдений пока немного. Одним из наиболее отчетливых изменений, установленных в развитии колоний у кораллов, являются различия в степени интеграции составляющих их особей, например, они сильно выражены в эволюции склерактиний [552]. У палеозойских граптолитов отмечена сильная зависимость нормального хода астогенеза от жизнедеятельности начальной особи [2404].
В целом ископаемые остатки, относящиеся к различным фазам индивидуального развития и их сериям, могут содержать информацию о любых аспектах жизни и эволюции древних организмов, а не только об их онтогенезе как таковом. Например, остатки и яйцевые оболочки позвоночных наряду с их использованием для целей стратиграфии, систематики и выяснения родственных связей отдельных групп [2514] могут также быть индикаторами условий инкубации или особенностей климата, выявляемых по микроструктуре и изотопному составу [1742, 2280]. Находки молодых экземпляров или яиц в полостях тела взрослых особей указывают на различные особенности их биологии - например, на каннибализм или живорождение у ряда палеонискоидных и ганоидных лучеперых рыб [1322, 1546], наличие полового диморфизма у аммонитов [1986], эволюцию строительного поведения у личинок ручейников [1022]. Для реконструкции палеоклиматических обстановок и свойственных им сезонных или других периодических изменений огромную роль играет изучение у различных беспозвоночных ритмов онтогенетического роста скелета, морфометрических особенностей личиночных раковин и других черт развития (см. 1.6).
Однако главной практической целью изучения онтогенеза и его стадий у ископаемых форм остается выяснение родственных отношений между различными организмами и их группами. Как можно видеть из многочисленных работ, посвященных этим вопросам, в основе всех выводов о родственных связях, полученных с помощью сравнительно-онтогенетического метода, лежит использование двух главных принципов - закона зародышевого сходства К. М. Бэра и биогенетического закона Э. Геккеля, указывающего на сохранение (рекапитуляцию) черт взрослых предков в онтогенезе потомка*. Наличие у двух сходных форм, например родов аммонитов, существенных различных черт в раннем онтогенезе, соответствующих ювенильным или взрослым особенностям других далеких друг от друга форм или групп, служит для палеонтолога ясным указанием на отсутствие близкого родства между сравниваемыми родами; и наоборот, сходство ранних стадий у дефинитивно различающихся форм доказывает их родство; см., например [131, 330].
* (О сущности этих двух законов и их взаимоотношениях см. подробнее в 7.4.)
На подобных принципах строятся и все заключения о происхождении различных вымерших групп, основанные на сравнительном изучении их онтогенезов, например, о возникновении одиночных склерактиний из ругоз-полицилин [438, 439, 2221], о филогенезе фораминифер-глоботрунканид [627, 628], различных групп аммоноидей [91, 305, 701, 702, 887, 2286] и т. д. То же касается и изучения онто- и филогенетической эволюции отдельных структур - ручного аппарата у ряда групп брахиопод [306, 969], септальных трубок у аммоноидей [343], зубного аппарата у двустворчатых моллюсков [1288], отпечатков аддукторов у остракод [266] и др.
Палеоэмбриологическими исследованиями накоплен огромный фактический материал, давший эволюционизму наиболее убедительные доказательства реальности гетерохронии, рекапитуляции, консерватизма ранних стадий (= зародышевого сходства) и других феноменов, отражающих закономерности морфологической эволюции онтогенеза. Все они подтверждают то положение, что эволюция взрослой стадии начинается всегда с нее самой и что именно этот процесс определяет ход исторических преобразований всего индивидуального развития [1178]. Однако приходится признать, что на практике попытки реконструкции эволюционных событий на основании онтогенетических изменений, наблюдаемых на ископаемом материале, все еще в большей мере строятся на неприемлемых теоретических представлениях, отождествляющих ход исторического процесса с его итоговым отражением в индивидуальном развитии (подробнее об этом см. в 7.4).
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'