
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
1.6. Палеофизиологические исследования (М. А. Шишкин, В. П. Макридин)
Представления о физиологии вымерших организмов могут быть получены многими путями, среди которых важная роль принадлежит анализу морфологических особенностей ископаемых организмов. Существенными источниками палеофизиологической информации служат также данные о микроскопическом строении и вещественном составе исследуемых скелетных остатков и условиях их захоронения (см. 2.1-2.6, 3.1-3.4).
Первоначально термин "палеофизиология" был введен Я. В. Самойловым [2199] для обозначения области исследований, охватывающей прежде всего биохимическую эволюцию организмов (см. 2.6, 2.7). Стремясь выяснить роль живых существ в осадочном накоплении химических элементов, участвующих в метаболических процессах, Я. В. Самойлов одним из первых указал [906, 907], что физиологическая деятельность вымерших форм должна была отличаться от современной. С этой точки зрения он пытался объяснить, например, накопление меди в пермских отложениях распространением организмов, у которых перенос кислорода кровью осуществлялся с помощью медистых соединений - гемоцианинов. Я. В. Самойловым был поставлен вопрос об эволюции скелетообразования, в которой первым этапом, по его мнению, было появление кремниевых скелетов.
Другим основателем палеофизиологии считают венгерского палеонтолога Ф. Нопшу [2017, 2018], который предполагал связь ряда эволюционных изменений в скелете позвоночных (гигантизм у динозавров, явления пахиостоза у мезозойских морских рептилий и т. д.) с нарушениями функции гипофиза. Подобные идеи выдвигаются и в наше время, например, для объяснения эволюции зубной ткани у млекопитающих [2486]. К этому направлению примыкают и многочисленные гипотезы, связывающие вымирание тех или иных групп организмов с глобальным воздействием абиотических факторов, нарушающим физиологическую норму: гибель динозавров от изменения уровня радиации или состава атмосферы, похолодания, отравления растительными алкалоидами; вымирание крупных млекопитающих от дефицита кобальта в пище, морских беспозвоночных - от изменения солености и т. д. [299]. Но палеофизиология имеет и более реальные достижения, связанные с изучением жизнедеятельности вымерших организмов, основанном на анализе их остатков и условий захоронения.
В целом предмет палеофизиологии как раздела физиологии [2168] составляют различные нормальные проявления жизненной активности, запечатленные в ископаемом материале, в первую очередь - различные реакции органов и тканей в процессе их развития и функционирования. Сюда же естественно отнести и физиологически обусловленные формы поведения, такие, как биология размножения и забота о потомстве, а также специфические требования организма к физико-химическим параметрам среды, изучаемые палеоэкологией (см. 3.1). В палеонтологии трудно разграничить область изучения физиологических отправлений и функциональной деятельности вообще. Последняя наиболее широко изучается в отношении механических процессов, в особенности связанных с передвижением и питанием. Это направление исследований составляет главный предмет морфофункционального анализа (см. 1.1-1.3).
Информацию о физиологии вымерших форм получают главным образом методом аналогий. Если ископаемый организм принадлежит к ныне существующей группе, то тем самым некоторые общие особенности его биологической организации становятся ясными сразу же. Например, установив принадлежность кости к млекопитающему, мы определяем и ее принадлежность теплокровному животному. Если мы относим строматолитную постройку к деятельности сине-зеленых водорослей, то тем самым подразумеваем и способность создавших ее организмов к фотосинтезу. Естественно, что для видов, близких или тождественных современным, степень физиологической аналогии максимальна. Поэтому находки таких форм обычно служат важными индикаторами среды обитания. Так обстоит дело, например, с неогеновыми моллюсками и рыбами или кайнофитными насекомыми.
Чаще, однако, возникает задача биологического истолкования более частных особенностей, и в этом случае используют сопоставление с гомологичными или по крайней мере морфологически сходными структурами у рецентных организмов. Объектами сравнения могут служить также минеральный и химический состав остатков, характер изменчивости, следы жизнедеятельности, особенности стадий развития и другие факторы. Все это, конечно, дает результаты лишь при условии, что нам известны корреляции этих признаков с физиологическими показателями у ныне живущих форм. В тех случаях, когда исследуемый орган не имеет достаточно близких рецентных аналогий, может быть использован метод построения парадигм, т. е. абстрактных функциональных моделей [2190]. Из серии таких моделей, соответствующих разным путям истолкования органа, наиболее правдоподобна та модель, которая позволяет правильно предсказать его особенности, не учтенные в модели, а также его изменения в тех или иных реально встречающихся вариантах (см. 1.1).
Метод парадигм пригоден там, где истолкование функции органа позволяет дать количественные оценки его возможных модификаций. Нередко он сочетается с методом аналогий. В целом, однако, последний остается главным инструментом исследования.
Морфология скелетных остатков. Морфологические сопоставления - наиболее обширный источник физиологической информации, особенно для организмов, у которых скелет в большой мере отражает строение мягких органов, а морфофизиологические корреляции достаточно хорошо изучены (позвоночные, членистоногие). Так, следы легочных мешков у девонских Placodermi ясно указывают на способность этих примитивных челюстноротых к воздушному дыханию [1495]. Отпечатки сейсмосенсорных органов на костях многих древних амфибий - лабиринтодонтов и лепоспондилов - говорят об ориентации в водной среде с помощью восприятия низкочастотных колебаний, а следы жабр и бранхиальных зубчиков у некоторых их личинок указывают на развитие через стадию жаберного дыхания и преимущественно фильтрационного питания.
У зверообразных рептилий переход к диафрагмальному типу дыхания, свойственному млекопитающим, может доказываться обособлением поясничного отдела позвоночника, а следы развития мягких губ у высших форм свидетельствуют о первых шагах в развитии маммального способа вскармливания детенышей [1033]. Строение слуховой косточки у древних амфибий и рептилий позволяет определенно различать две системы восприятия звуков - из воздуха (через барабанную перепонку) и из субстрата (через челюстной сустав). У палеоген-неогеновых китообразных строение звуковой улитки отражает разную степень приспособления к высокочастотной эхолокации [1584]. Вынашивание молоди или яиц в сумке матери весьма вероятно для меловых мультитуберкулят и плацентарных млекопитающих и даже для высших триасовых зверообразных рептилий - тритилодонтов, у которых найдена сумчатая кость, характерная для современных Marsupialia и однопроходных [1599, 1831]. Живорождение в разных группах позвоночных доказывается на ископаемом материале непосредственными находками молоди в полости тела у отдельных палеонискоидных рыб (Birgeria) и морских рептилий [1322, 1984]. Обнаружение молоди раннепермского лабиринтодонта Trimerorhachis в жаберных мешках взрослых особей [2028] имеет близкие аналогии у некоторых современных лягушек, вынашивающих личинки в голосовых мешках.
Для позвоночных животных, особенно млекопитающих, одним из важных источников данных, касающихся их поведения и взаимоотношений со средой обитания, является изучение мозговых структур по отпечаткам полости черепа, основанное на сравнении с близкими рецентными формами. Это направление получило наименование палеоневрологии. Примером таких исследований может служить работа Ю. А. Орлова [786], посвященная изучению слепка мозговой полости гигантской позднемиоценовой куницы Perunium. Преобладающее развитие височных и затылочных долей большого мозга позволило сделать вывод об опережающей дифференциации. У этой формы слуха и зрения по сравнению с функциями лобных долей, тогда как сильное развитие полушарий мозжечка заставляет предполагать сложную работу конечностей, которые, по-видимому, могли использоваться для лазания по деревьям (см. также 1.1).
Среди морфофизиологических процессов, непосредственно отражаемых строением ископаемых беспозвоночных организмов, особенно велик круг явлений, связанных с размножением и развитием: клеточные деления у простейших, почкование у колониальных организмов, линька у членистоногих и т. д. Многие другие особенности также становятся понятными благодаря рецентным аналогиям. Таким путем, например, выявлено наличие у ископаемых мшанок системы экстразооидальных водных потоков, обеспечивающих поступление пищи, выведение отфильтрованных вод, а также спермы и личинок [2367]. На этой же основе проведен анализ водно-сосудистой системы у ископаемых иглокожих [2005]. У трилобитов сопоставление с современными членистоногими позволяет говорить о респираторной функции щечной цеки у уплощенных форм [1800].
Во многих случаях, однако, близкие аналогии неизвестны, и остается лишь возможность моделирования предполагаемой функции. Так, трубчатые выросты края раковины у некоторых мезозойских брахиопод истолковываются как вместилища чувствительных краев мантии, действовавших подобно антеннам и подававших сигнал к закрытию створок при приближении опасности [2191]. Хорошо обоснованные парадигмы построены также для сенсорных органов камератных криноидей. Предполагается, что у этих реофильных животных поры теки и связанные с ними пиннулы представляли собой систему, чувствительную к горизонтальным течениям и помогающую выборочно извлекать пищевые частицы из водного потока [1711]. Широкие возможности для моделирования функции предоставляют глаза трилобитов, поскольку они могут сохранять свою структуру довольно полно. Анализ шизохроических глаз как оптической системы позволяет говорить об их способности к целостному восприятию изображений и устранению сферической аберрации [1340, 1429].
Нередко выбор правильной интерпретации затруднителен - например, в отношении дополнительной функции анального мешка у каменноугольной криноидей Aesiocrinus, снабженного порами и каналами. Предполагают, что он либо выполнял респираторную функцию для задней кишки, либо играл роль гипертрофированной мадрепоритной системы [1862]. Также неясно, назначение срединного бугорка глабели у трилобитов, который мог быть либо глазом, либо органом осязания и регистрации давления [1594].
В случаях, когда для современных организмов известны внешние причины определенных морфологических модификаций, возможно объяснить соответствующие изменения и у близких ископаемых форм. Например, у некоторых двустворок (Cardiidae) уменьшение размеров раковины и числа ребер связано с понижением солености водной среды. Размеры личинки и, следовательно, продиссоконха у двустворок часто обратно пропорциональны температуре воды [1908]. У остракод температура воды влияет на степень скульптированности раковины, а содержание кислорода в воде - на ширину внутренней пластинки и глубину вестибюля [1406, 2082].
Морфофизиологические интерпретации, основанные на аналогиях, используются и в палеоботанике. Митотические деления описаны у докембрийских одноклеточных водорослей [240]. У палеозойских птеридоспермов предполагают секретирование семезачатком капли жидкости для улавливания пыльцы, как у многих современных голосеменных. Это обосновывается находкой семезачатка с неклеточным образованием снаружи, содержащим пыльцу [2180].
Микроскопическое (гистологическое) строение. Изучение микроструктуры остатков беспозвоночных в сечениях и на естественных поверхностях дает обширную информацию об изменениях секреторной деятельности скелетообразующих тканей - циклических или же связанных с образованием различных структур скелета. Так, у кораллов пояски прироста могут соответствовать суточным, месячным и годичным ритмам, что позволяет судить о длительности астрономического года и лунных циклов в прошлые геологические эпохи [2257, 2477]. Пять различных ритмов (от суточного до годичного) установлены по структурным различиям в линиях нарастания у двустворчатых моллюсков [1307]. При этом скорость роста преимущественно зависит от внешних факторов, например у кораллов, - от температуры воды и освещенности [1295], хотя известную роль играют и особенности самих организмов [551, 552]. У литоральных моллюсков отложение карбонатных слоев раковины соответствует приливам, а органического - отливам [2067]. Образование слоев наружной раковины отражает различные физиологические состояния секретирующей ткани (у моллюсков и брахиопод - эпителия мантии) или деятельность различных ее участков. Некоторые секреторные процессы, такие, как образование третичного слоя раковины у брахиопод, зависят от температуры и других условий среды [481, 1911].
У наземных позвоночных одним из важнейших физиологических показателей, получаемых путем изучения микроструктуры скелета, является характер терморегуляции. У современных эктотермных животных периостальные ткани трубчатых костей относятся к пластинчато-зональному типу, характеризующемуся бедной васкуляризацией и наличием годичных колец. Напротив, у эндотермных групп (птиц, млекопитающих) эти ткани имеют волокнисто-пластинчатое строение, не содержат отчетливо выраженных годичных колец и снабжены сильно развитой сосудистой системой. Наличие тканей второго типа у терапсид, динозавров и птерозавров дает основание ряду авторов считать этих рептилий теплокровными [1297, 1298, 2161, 2164, 2167]. В отношении динозавров имеются, правда, противоречащие данные о наличии годичных колец в микроструктуре зубов [1804]. У лабиринтодонтов, способных подобно современным амфибиям к кожному газообмену, его интенсивность отражается в степени развития сосудистой сети в покровных черепных костях [1395]; (рис. 47).
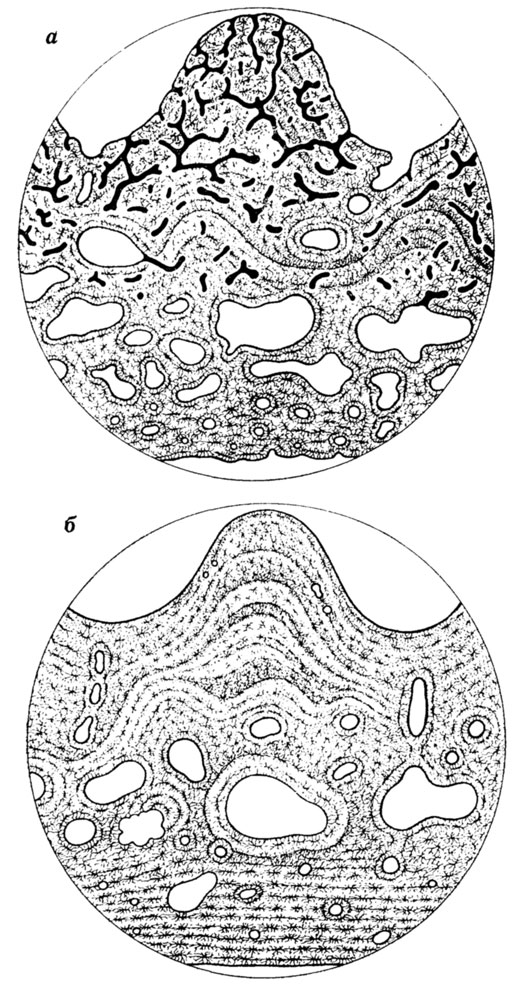
Рис. 47. Микроструктура покровных костей черепа (в вертикальных срезах) у пермских и триасовых амфибий - лабиринтодонтов как показатель интенсивности кожного газообмена. По А. П. Быстрову. а - 'гидрофильный' тип с хорошо развитой сетью сосудистых кожных капилляров у Benthosichus; б - 'ксерофильный' тип, не имеющий капилляров, у Platyoposaurus
Исследования скорлупы яиц верхнемеловых динозавров - завропод, протоцератопса [2280] - показало, что газо- и влагообмен осуществлялись у них путем диффузии сквозь многочисленные поры. Сопоставление же с яйцами современных рептилий и птиц привело к предположению, что вылупившиеся детеныши этих динозавров имели малые размеры, а по уровню метаболизма были близки к тем ныне живущим рептилиям и птицам, которые строят гнезда на земле в условиях высокой влажности и воздушной среды, обедненной кислородом при повышенном содержании углекислого газа (что обычно имеет место при подземной инкубации или искусственном зарывании кладок).
Содержимое пищевых путей и непереваренные остатки пищи. О характере питания и пищеварения часто можно судить по содержимому желудка и кишечника. У животных-илоедов (трилобитов, некоторых моллюсков, червей и др.) пищевые пути благодаря заполнению субстратом могут фоссилизоваться. Например, у раннеюрской двустворки Nuculana известен внутренний слепок кишечника с продольными бороздками (отпечатками стенок), аналогичными бороздкам на фекальных выбросах у современного близкого рода Nucula [1458]. Нередко сохраняются твердые части желудочного содержимого. Так, в кишечнике насекомых обнаружена пыльца растений [528, 2254], в желудке аммонитов - остатки различных мелких беспозвоночных [1872], у юрских акул - непереваренные ростры белемнитов и раковины аммонитов, у позднемелового динозавра Anatosaurus - побеги и плоды хвойных растений [1984]. У ихтиозавров в содержимом желудка, а иногда и кишечника встречаются остатки мягкотелых головоногих, рыб и планктонных организмов, в том числе мелких рачков. Иногда возможность питания ихтиозавров головоногими оспаривается [1823]. Другие исследователи [2273] приводят данные о находках в желудке ихтиозавров крючков (онихид) от рук белемнитов и подчеркивают, что ростры в желудочно-кишечном тракте этих организмов не встречаются.
Это дает им повод считать, что ихтиозавры питались мягкими тканями белемнитов. Позволительно, однако, предполагать и то, что ростр у белемнитов, как и у некоторых современных кальмаров, был эластичным [454].
Иногда прослеживается характер переваривания пищи. У олигоценового представителя скумбриевых из семейства Cybiidae в желудке были найдены мелкие рыбы, из которых наиболее близкие к кишечнику (ранее заглоченные) максимально размацерированы [2465]. Особенность пищеварения дневных хищных птиц - полное растворение костей добычи и характерная эрозия непереваренных зубов; поэтому остатки погадок этих птиц в четвертичных отложениях достаточно уверенно опознаются [1932]. О механизме размельчения пищи говорят находки гастролитов, известные у ископаемых птиц и морских рептилий. Важным источником сведений о характере питания служат копролиты [204], хотя их принадлежность не всегда удается идентифицировать.
Химический состав. Анализ количественных соотношений элементов в составе известкового скелета у водных беспозвоночных дает возможность судить о температурном и солевом режимах среды обитания по аналогии с современными формами. Методическая сторона этих исследований освещена в специальных разделах (см. 2.6, 3.1).
Отметим, что распределение магния и других элементов может быть неравномерным, отражая цикличность секреторной деятельности организма [1871] или ее закономерное изменение с возрастом, при переходе в иные условия обитания [1527]. Имеются, впрочем, данные о том, что и накопление магния в целом может в большой степени физиологически контролироваться самим организмом, в зависимости, например, от того, секретирует ли он кальцит или арагонит [1358, 2457]. Изотопный состав может отражать помимо температуры и солености водной среды также изменения скорости роста [1813, 2061].
Сопоставление возрастных стадий. Этот метод дает много сведений о физиологии развития и размножения. Количественный анализ массового материала (например, по раковинам моллюсков) позволяет проследить закономерные изменения в характере смертности в течение жизненного цикла, которые могут быть разными у близких форм одного биотопа [1743]. В случае, когда у колониальных организмов наблюдается постепенное изменение в завершенности морфогенеза очередных генераций зооидов, можно с большой уверенностью говорить о физиологических градиентах, закономерных изменениях концентрации стимуляторов или ингибиторов, управляющих развитием колонии. Это, в частности, имеет место при образовании субколоний мшанок из монтикулярных структур [1278]. Наличие в одновременном захоронении новорожденной и трехмесячной особей пещерного медведя позволяет судить о продолжительности брачного периода, который был длиннее, чем у современного бурого медведя [1544]. Ход личиночного развития у палеозойских лабиринтодонтов доказывает отсутствие у них резкого (катастрофического) метаморфоза, свойственного их потомкам - бесхвостым амфибиям.
Межвидовые численные соотношения. Относительная численность организмов внутри биоценозов может служить их важной биологической характеристикой. Например, количественное отношение "хищник - жертва" среди млекопитающих составляет обычно не более 4%, тогда как у эктотермных позвоночных оно во много раз выше. Поэтому определение на массовом материале по меловым динозаврам очень низкого соответствующего показателя (3%), косвенно свидетельствует о теплокровности этих животных [1297].
Особенности захоронения и литология вмещающих пород. Связь этих показателей с физиологией захороненного организма может быть очень тесной. В общих чертах она обычно определяется уже тем, с какой из двух главных обстановок захоронения - морской или континентальной - мы имеем дело. На этом фоне выявляются более частные зависимости. Так, первичная пиритизация глубоководных протерозойских сланцев Северной Австралии, указывающая на восстановительную среду их накопления, позволяет говорить об анаэробности найденных здесь бактерий и водорослей [2023]. Биостратономический анализ захоронений некоторых неогеновых гастропод Паратетиса указывает на их переход от легочного дыхания к жаберному [288]. Находки в пермокарбоновых континентальных красноцветах Техаса характерных нор с остатками лепоспондильных амфибий Lysorophus свидетельствуют о существовании фазы спячки в образе жизни этих животных, что подтверждается и находками здесь аналогичных нор двоякодышащих рыб [2029]. Представители двух групп триасовых лабиринтодонтов - трематозавры и плагиозавры - часто встречаются как в континентальных, так и прибрежно-морских фациях, что указывает на их способность (подобно многим современным лягушкам) переносить довольно высокую соленость, оставаясь в норме пресноводными животными.
Палеофизиологические данные имеют огромное значение для изучения эволюции биосферы (см. 8.4). Они позволяют судить о физико-химических условиях среды обитания и тем самым об условиях осадконакопления в геологическом прошлом. Продукты жизнедеятельности организмов, как и их непосредственные остатки, могут быть материалом породообразования (например, накопление копролитовых горизонтов). Изучение ритмичности скелетообразования у беспозвоночных открывает в перспективе возможности для нового пути определения возраста их остатков (если будет достаточно точно рассчитано изменение числа дней в году для прошлых эпох). Изучение ископаемого материала позволяет также изменить представления о многообразии ранее существовавших физиологических типов. Функциональные сочетания, не имеющие аналогий в современном органическом мире (например, сохранение рудиментов амфибийной сейсмосенсорной системы на рептилийном уровне организации), нередко оказываются реальностью при изучении древних организмов.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'