
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
2.5. Микроструктурные исследования в палеоботанике (С. В. Мейен)
Термин "гистология" почти не употребляется в ботанике и палеоботанике, хотя Э. Гофман [1746] и назвала свою книгу "Палеогистология растений". Гораздо употребительнее термин "анатомия растений", означающий любое микроструктурное исследование - от ультраструктуры клеточных стенок до расположения тканей в органе. В последние десятилетия отмечается все большее сближение анатомии и морфологии растений, хотя самостоятельность обеих дисциплин еще сохраняется в вузовских программах, библиографических рубрикаторах, названиях лабораторий и в других случаях. Показательно, что в учебниках и сводках по анатомии растений приводятся сведения об общем облике органов, т. е. об их морфологии, а в аналогичных изданиях по морфологии все больше места отводится изложению микроструктуры органов.
Прогрессирующее слияние анатомии и морфологии, наблюдаемое при изучении и современных, и ископаемых растений, отчасти связано с тем, что само разграничение макро- и микроструктурных признаков очень условно. Например, жилкование листа может интерпретироваться по-разному. Даже если считать общее расположение жилок макроморфологическим (эпиморфологическим) признаком, то все равно для изучения наиболее тонких жилок и поведения жилок у края надо прибегать к микроструктурным (микроморфологическим) исследованиям. У ископаемых растений о расположении жилок порой приходится судить по упорядоченности эпидермальных клеток. Если же мы хотим проследить ход жилок из стебля в лист, то это будет уже классический пример анатомического исследования.
Тем не менее можно привести аргументы и в пользу самостоятельности анатомии и морфологии растений в некоторых существенных отношениях. В теле высшего растения, как любого другого многоклеточного организма, с большей или меньшей четкостью намечается иерархизация структуры, образующая ряд "органы - ткани - клетки - внутриклеточные структуры (вплоть до молекулярных)". Соответственно уровням этой иерархии можно обособить и области исследования - морфологию (точнее, органологию), гистологию (анатомию), цитологию и молекулярную биологию. Иногда разграничение задач между этими дисциплинами вполне возможно. Нас может интересовать листорасположение, формулы которого устанавливаются безотносительно к анатомическим признакам листьев и стебля. Гистологическим (анатомическим) будет исследование признаков вторичной древесины, когда общая архитектоника ствола опускается. Цитологические исследования на палеоботаническом материале выполняются тогда, когда производится сравнение поровости трахеид у разных растений и прослеживается эволюция этого признака.
Самостоятельность макро- и микроморфологического исследования сохраняется в палеоботанике и по ряду других причин. Палеоботаник нередко вынужден ограничиться макроморфологическим исследованием, если микроструктура остатков не сохранилась. В микроструктурном исследовании иногда нет необходимости (если точное определение систематического положения остатка возможно и без того) или возможности (в условиях производственной определительской работы). Некоторые исследования, наоборот, не затрагивают область макроморфологии. Так, при изучении обломков вторичной древесины мы ограничиваемся микроструктурными признаками.
Разграничение макро- и микроморфологических признаков и соответственно морфологии и анатомии особенно условно при изучении микроскопических остатков или органов, имеющих микроскопические размеры. Так, споры и пыльца в приведенной выше иерархии структур могут рассматриваться и как органы (когда они уже высеялись из спорангиев или когда мы имеем в виду мегаспоровую мембрану семени), и как клетки (когда они находятся на стадии спороцитов). Заметим, что в отечественном реферативном журнале "Ботаника" принято помещать рефераты по строению спор и пыльцы в рубрику "Морфология". Расположение корневых органов вроде бы должно входить в предмет морфологии. Однако когда речь идет о микроскопических ризоидах моховидных, то их расположение описывается вместе с микроструктурными признаками, что определяется не какими-либо принципиальными соображениями, а просто применением микроскопической техники.
Из-за тесной связи морфологии и анатомии в ботанике нет смысла здесь рассматривать все аспекты анатомических исследований ископаемых растений, так как тогда пришлось бы повторять многое из того, о чем сказано в главе, посвященной морфологическим исследованиям. Поэтому ниже рассмотрены лишь вопросы, касающиеся собственно микроструктурных исследований в палеоботанике.
Ископаемые растения стали объектом анатомического исследования на заре развития палеоботаники. Когда Р. Гук пришел к выводу, что окаменелости, внешне сходные с древесиной, действительно являются таковой, то он опирался прежде всего на микроструктурные признаки. Клеточная структура ископаемых растений нередко хорошо видна просто в сильную лупу. Поэтому не удивительно, что анатомические признаки некоторых ископаемых растений отмечались в описаниях и включались в иллюстрации ранних палеоботанических работ. В публикациях 30-х годов прошлого века мы находим уже довольно много анатомических сведений. Специально анатомией ископаемых растений первым занялся Г. Уитхэм, применивший технику изготовления тонких шлифов. Соответствующие его публикации относятся к 1831 и 1833 гг. С тех пор в литературе регулярно появлялись статьи и даже монографии, специально посвященные той же теме. Уже тогда палеоботаникам стало ясно, насколько расширяется представление об ископаемых остатков растений, если удается изучить их анатомическую структуру. Анатомические признаки довольно быстро проникали в диагностику многих вымерших таксонов. Дальнейшая история анатомических палеоботанических исследований изложена в книгах Г. Н. Эндрьюса [1274] и Э. Гофман [1746]. Сейчас технические средства, позволяющие изучать микроструктуру растительных остатков, весьма обширны. Кроме традиционных, введенных еще в прошлом веке методов мацерации фитолейм и изготовления шлифов и пришлифовок, сейчас применяют разнообразные другие методы - приготовление пленочных оттисков и латексных слепков, разные способы травления остатков, электронную микроскопию и др. [683, 2370].
Поскольку литература по анатомии растительных остатков необозримо велика, мы остановимся только на наиболее общих вопросах - на смысле микроструктурных исследований, их возможностях и ограничениях. Общий смысл микроморфологических исследований можно свести к следующим основным положениям.
1. Многие типы органов могут быть установлены лишь путем анализа анатомических признаков. Так, без обращения к этим признакам невозможно различить у зигоптеридиевых папоротников стебли, филлофоры и черешки. Само по себе взаимное расположение этих частей в теле растения не позволяет их разграничить. Без проведения анатомических исследований легко совершить крупные морфологические ошибки. Спорофиллы плауновидных на отпечатках бывают очень сходными с семенными чешуями хвойных. Оба типа органов нередко смешивались. Пластинка спорофилла имитировала семенную чешую, спорангий - семя, и наоборот. Можно легко принять ложный ствол некоторых папоротников, сложенный компактно расположенными черешками и воздушными корнями (иногда те и другие заключены в паренхимную массу), за настоящий ствол. Случалось, что окатанные обломки таких ложных стволов принимались за стробилы. В этих случаях простейшее анатомическое исследование позволяет вскрыть или предотвратить ошибку.
2. Микроструктурные исследования резко расширяют круг признаков растительных остатков, делают наши знания несравненно более полными.
3. Перед палеоботаникой стоит непреходящая задача установления прижизненной связи частей, встречающихся в захоронениях дисперсно. Микроморфологические признаки широко используют как маркеры, указывающие на принадлежность разных частей (например, листьев и фруктификаций) одним и тем же растениям (см. 1.4.).
4. Поскольку изменения в морфологических интерпретациях и установление новых признаков (в том числе касающихся общего облика растений, реконструированного с помощью маркеров) так или иначе ведут к таксономическим следствиям, микроморфологические исследования стали непременным условием полноценного таксономического анализа. Многие группы ископаемых растений вообще не поддаются систематизации без привлечения микроморфологических признаков. Таковы моховидные, многие папоротники и голосеменные. Основные диагностические признаки родов ископаемых моховидных - это микроструктура листьев и талломов. Бывали случаи, когда остатки мхов относили по эпиморфологическим признакам к хвойным и плауновидным. Без изучения микроструктурных эпидермально-кутикулярных признаков листьев нельзя надежно определить многие роды цикадовых, гинкговых, хвойных и других групп. Точное определение и систематизация папоротников требует знания микроструктуры спорангиев и спор, а в некоторых случаях - черешков, филлофоров и стеблей. В палеоботанике от микроструктурных исследований зависит не только морфология, но и номенклатура растений. Остатки одних и тех же растений могут обозначаться разными родовыми и видовыми названиями в зависимости от того, известна или нет их микроструктура (см. 4.1).
5. Поскольку точная диагностика растительных остатков связана с микроморфологическими исследованиями, последние приобретают непосредственное практическое значение всюду, где необходима точная диагностика, т. е. в стратиграфии, палеобиогеографии, палеогеографии, палеоэкологии, тектонике и других дисциплинах. Нередко случается, что группы растений, имевшие незначительное стратиграфическое значение, приобрели статус важных руководящих ископаемых после того, как их остатки были изучены микроструктурно и систематика была соответственно существенно усовершенствована. Таковы листья верхнепалеозойских кордаитантовых Ангариды, мезозойских гинкговых чекановскиевых и хвойных.
Некоторые микроструктурные признаки могут служить хорошими индикаторами той среды, в которой произрастали растения Общеизвестно значение слоев (колец) прироста и особенно их отсутствия как показателя климатических условий (слои прироста отсутствуют только у растений несезонного экваториального климата). Многолетние маноксилические стволы известны только в безморозном климате. По воздушным полостям в корнях можно судить о сильном обводнении местообитаний. Перечисленные признаки относятся к микроструктурным. Можно еще упомянуть ксероморфные признаки кутикулы, которые не обязательно прямо указывают на сухость местообитаний и нуждаются для экологической интерпретации в учете дополнительных признаков, таких, как макроморфология листьев и побегов, структура захоронений и др.
7. Изучение микроструктуры растительных остатков имеет большое значение для выяснения материнского вещества и условий образования угольных пластов. В этих случаях изучают микроструктуру остатков в угольных почках и непосредственно в угле.
Естественно, что микроструктурные исследования возможны далеко не всегда. Нередко встречается растительный остаток настолько плохой сохранности, что никакими методами изучить его микроструктуру невозможно, особенно если это только отпечаток на грубой породе. Однако очень часто могут быть изучены хотя бы какие-то микроструктурные признаки, в особенности относящиеся к поверхности листа или иного органа. Оттиски клеток отпечатываются на породе, видны на угольной корочке (фитолейме) или покрывающей ее кутикуле Реже встречаются остатки с минерализованными внутренними тканями. Применяемая в этих случаях палеоботаническая техника заключается, во-первых, в специальной обработке растительных остатков для "проявления" скрытых структур (мацерация, изготовление срезов, травление и др.) и, во-вторых, в специальной технике наблюдения бывших изначально или выявленных в ходе обработки структур (световое или электронное микроскопирование, фотографирование в различных лучах и др.).
Сейчас нельзя назвать ни одной микроморфологической особенности растений, которая заведомо никогда не сохраняется и потому никогда не будет известной палеоботаникам. Степень полноты сохранности микроструктур бывает поразительной (рис. 80). Могут сохраняться даже такие внутриклеточные структуры, как тилакоиды хлоропластов и хромосомы [1368, 2012]. Кажется очевидным, что сохранение столь тонких структур возможно лишь в исключительно благоприятных тафономических условиях. Действительно, такие тонкие детали удавалось наблюдать лишь в очень немногих случаях. Однако вполне вероятно, что сохранение очень тонких структур более обычно, но мы просто еще не нашли средств эти структуры выявлять. На такую возможность указывают недавние исследования высокометаморфизованных углей с помощью метода ионного травления [485]. В углях, в которых обычными углепетрографическими методами не обнаруживается практически никаких первичных структур растительных остатков, ионное травление выявляет весьма тонкие детали. Это не только форма и расположение клеток, но и внутриклеточные органеллы, включая плазмодесмы между клетками. Вполне возможно, что в дальнейшем удастся применить ионное травление и для менее метаморфизованных углей, а также для изучения микроструктуры фитолейм на отпечатках, не поддающихся обычным микроструктурным методам изучения.
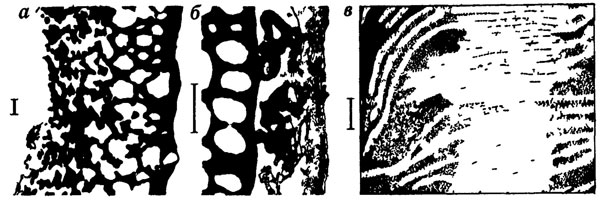
Рис. 80. Сохранность ультраструктуры растений в ископаемом состоянии. а - предпыльца Dolerotheca formosa Schopf (Trigonocarpales) поперечный разрез экзины с альвеолярной структурой, б - такая же предпыльца, показана внешняя часть экзины с сохранившимися остатками тапетальной мембраны, в - строение хлоропласта из миоценового листа Betula, сохранилась мембранная система, на а, б линейка 1 мкм, на в - 100 нм, рис. а, б по фотографиям из Т. И. Тейлора, в - из К. М. Никласа и Р. М. Брауна
Каким образом сохраняются наиболее тонкие структуры растений, пока неизвестно. Впрочем, переход в ископаемое состояние и более грубых структур во многом непонятен [2235]. Лучше всего казались изученными процессы преобразования клеток и тканей в углях, но, как показало применение ионного травления, в действительности эти процессы были поняты неправильно.
Серьезные ограничения в применении микроструктурных методов - деформация растительных остатков, их сильное сплющивание, нередко сопровождаемое боковым смещением, перекристаллизация внутренних частей растений, коррозия органического вещества и другие вторичные процессы Материал может пострадать и по вине людей от отсутствия должной осторожности при сборах и хранении образцов. Случается, что фитолейма или минерализованная корочка теряется, а остается только отпечаток растения.
Изучение микроструктуры растительных остатков связано и с трудностями в интерпретации наблюдений. Требуется разобраться в типах клеток и тканей, их взаимном положении, в изменении микроструктуры по всему остатку. Далее надо разобраться в таксономическом, экологическом или ином значении выявленных признаков. В общем, здесь палеоботаник сталкивается с теми же вопросами, что и при интерпретации макроморфологических признаков. Микроструктуры столь же подвержены сложной изменчивости, так же часто проявляют параллелизм и конвергенцию, что и макроструктуры. Конвергенция микроструктур может быть удивительно полной. Например, даже с помощью электронной микроскопии не удается выявить признаки, отличающие тонкую организацию (строение мешка, микрогаметофита и др.) пыльцевых зерен каллистофитовых (класс Ginkgoopsida) и некоторых хвойных (класс Pinopsida). Независимость возникновения столь сходной структуры совершенно очевидна, так как у общих предков обоих классов была несравненно более примитивная пыльца, неотличимая от спор споровых растений. С другой стороны, тонкие микроструктурные признаки могут обнаруживать непонятное разнообразие в некоторых таксонах, хотя в других таксонах те же признаки очень устойчивы. Так, распределение проводящих пучков очень изменчиво в семезачатках покрытосеменных и весьма устойчиво у семезачатков голосеменных. Некоторые эпидермальные признаки изменчивы у палеозойских и устойчивы у современных хвойных. Когда не удается прямо сопоставить микроструктурные особенности ископаемых и современных растений, палеоботаникам приходится вводить нейтральные описательные термины (например, "клетки с темным содержимым").
Одной из интересных проблем будущих исследований можно считать распределение микроструктурных признаков по всей системе растений и по геохронологической шкале. Пока такая работа выполнена лишь для немногих признаков [1301, 1419, 1746]. Тем не менее некоторые закономерности распределения микроструктурных признаков в геохронологической шкале уже выявлены. Оказалось, что большинство микроструктурных признаков, свойственных высшим растениям в целом, появляется еще в девоне. В более поздние периоды вновь появляются лишь сравнительно сильно специализированные признаки (например, торус в окаймленных порах трахеид вторичной древесины известен лишь с юры; бабочковидная кутинизация замыкающих клеток - с перми; протосаккатная организация пыльцы - с карбона, рис. 81).
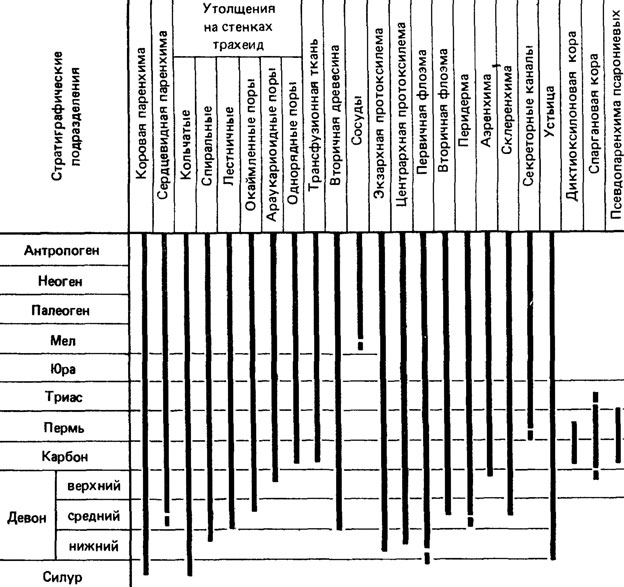
Рис. 81. Стратиграфическое распространение отдельных типов клеток и тканей высших растений. По Х. П. Бенке, У. Г. Челонер, И. Ширин, Э. Гофман
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'