
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
2.6. Палеобиогеохимические и палеобиохимические исследования (И. С. Барсков)
Начало исследованиям химического состава организмов и их скелетных частей положено В. И. Вернадским в связи со сформулированными им общими положениями биогеохимии о роли живого вещества в эволюции материи. Основополагающее значение имели труды А. П. Виноградова [171, 172], посвященные изучению химического состава морских организмов, работы Я. В. Самойлова [906], Ф. У. Кларка и Уиллера [1428], обосновавших применение биогеохимической методики к изучению палеонтологических объектов. Палеонтологические проблемы геохимического изучения растительных остатков, выдвинутые С. А. Ивановым [416] и В. А. Успенским [1085], получили в дальнейшем существенное развитие [1385, 2011], хотя более интенсивно это направление прогрессировало в рамках геохимии органического вещества (см. 2.7).
Несмотря на более чем полувековую историю, палеобиогеохимия все еще переживает трудный и противоречивый период становления. Несомненно, остаются справедливыми основные общетеоретические постулаты биогеохимии: 1) каждый биологический таксон обладает специфичностью своего химического состава; 2) в химическом составе каждого организма находит свое отражение геохимическая обстановка среды его обитания.
Вместе с тем необходимо признать, что в настоящее время отсутствуют теоретические научные обоснования, которые позволяли бы объяснить конкретные пути формирования специфичности химического состава организмов, уровень специфичности содержания отдельных элементов и соединений, биологические и физико-химические механизмы влияния среды на формирование химического состава различных частей и тканей организмов. Эти вопросы оказались значительно более сложными, чем представлялось вначале, и эмпирические материалы, накопленные к настоящему времени, пока не дали на них определенных ответов. Исследования химического состава ископаемых остатков организмов призваны учитывать влияние еще одного фактора - процессов фоссилизации, геохимические закономерности которых также известны мало.
В соответствии с тремя названными факторами, обусловливающими состав анализируемых ископаемых скелетных остатков (влияние физиологии организмов, влияние среды и влияние диагенеза в широком смысле), палеобиогеохимия формулирует свои задачи: исследование геохимической эволюции организмов, геохимических параметров среды прошлого и геохимических особенностей постседиментационных процессов. Недооценка хотя бы одного из этих факторов или переоценка любого из них, даже приводя на первых порах к видимому успеху, оборачиваются в конечном итоге ничтожной научной отдачей и могут привести к дискредитации всего научного направления.
Сложность интерпретации данных химического состава наглядно может быть проиллюстрирована историей исследований изотопного состава кислорода в биогенных карбонатах. Соотношение легкого (16О) и тяжелого (18О) изотопов кислорода, входящего в состав карбонатов, осажденных из воды, в соответствии с физико-химическими расчетами [2413] зависит от температуры воды. Эта термодинамическая зависимость, подкрепленная эмпирическими и экспериментальными данными об изотопном составе кислорода биогенных карбонатов раковин моллюсков, лежит в основе палеотермометрии [1942, 2144]. В 50-60-е годы нашего столетия изотопная палеотермометрия завоевала всеобщее признание, были выполнены многочисленные исследования, проведены палеоклиматические построения для многих промежутков геологического времени. Осложняющим моментом считалась лишь возможность диагенетического перераспределения изотопов, от чего можно избавиться подбором анализируемого материала хорошей сохранности.
В дальнейшем, однако, было показано влияние на изотопный состав кислорода раковинных карбонатов изотопного состава вод, в которых обитали организмы, строящие карбонатный скелет. Колебания изотопного состава кислорода воды в различных водоемах сказывались на изотопном составе биогенных карбонатов настолько сильно, что значительно перекрывали влияние температурного фактора. В тех случаях, когда была возможность внести соответствующие коррективы, палеотемпературные определения по биогенным карбонатам могли быть выполнены. В большинстве же случаев параметры изотопного состава вод древних водоемов оставались неизвестными, и палеотемпературные определения были сомнительными [1045].
В 70-е годы появились данные о том, что на изотопный состав кислорода биогенных карбонатов оказывают влияние также особенности физиологии организмов, образующих карбонатный скелет. В одинаковых условиях разные организмы накапливают стабильные изотопы кислорода в различных соотношениях, которые при пересчете на температуру обитания дают весьма различные значения [258]. Все это существенно подорвало доверие к изотопной палеотермометрии, привело к сокращению исследований в этом направлении. Причиной сложившегося положения явилась переоценка влияния одного фактора - среды - и недооценка влияния биологических особенностей организмов.
Вместе с тем существенно увеличившаяся сложность и неоднозначность интерпретации изотопных данных вовсе не должны приводить к отказу от таких исследований. Более того, это открывает возможности для использования изотопного состава кислорода биогенных карбонатов не только и, возможно, не столько для палеотермометрии, но и для выявления биологических особенностей организмов и их эволюции, величина 18О становится комплексным показателем физиологических факторов и геохимических параметров среды [491, 1045]. Причем данные по изотопному составу кислорода могут быть правильно истолкованы только при совокупном анализе с данными по изотопному составу углерода - другого основного элемента карбонатных скелетов. Пути поступления этих элементов в скелет при его минерализации в значительной степени общие, хотя если изотопный состав кислорода в большей степени зависит от физико-химических условий среды (температуры, гидрологического режима), то изотопный состав углерода более чувствителен к биологическим факторам. Изотопный состав углерода, содержащегося в углекислом газе и углекислоте, растворенных в воде и идущих на построение скелета организмов, зависит от его происхождения. Углерод, прошедший через биологический цикл, имеет более легкий изотопный состав.
Комплексные исследования по совокупному изотопному составу кислорода и углерода дают полноценные сведения для палеоокеанологического анализа, а на более детальном уровне исследований - для решения вопросов таксономии [491].
Изотопный состав углерода из органического вещества ископаемых, помимо хорошо известного применения для радиометрических датировок по радиоактивному изотопу 14С, используют и в палеобиологических целях. Отношение стабильных изотопов 13С/12С в органической фазе ископаемых костей применяют для определения характера диеты (растительной, животной, смешанной) древнечетвертичных животных и людей.
Как можно видеть, достаточно сложно обстоит дело даже с наиболее распространенными элементами, входящими в скелетные остатки организмов, - элементами, для которых имеются теоретически достаточно точно обоснованные физико-химические модели их поведения при образовании минеральной фазы скелета.
В составе карбонатных скелетов многих организмов, помимо основных элементов (Ca, O, C), обычно устанавливается присутствие около десятка малых элементов и элементов-примесей (Mg, Sr, Mn, Fe, Ba, Si, Ti, Cu и др.). Для всех этих элементов нет пока теоретических обоснований их присутствия в скелетном веществе, неизвестны причины и механизм их поступления и участия в построении скелета, а для многих неизвестно и место, которое они занимают в системе минерально-органического комплекса (скелета). Таким образом, строгая трактовка причин изменения содержания этих элементов в настоящее время не подкреплена теоретическими физико-химическими или биохимическими моделями. Тем не менее благодаря большому числу проведенных и проводящихся исследований некоторые эмпирические закономерности в распределении малых элементов и элементов-примесей в карбонатных скелетах многих организмов установлены и используются при таксономических, филогенетических и палеогеографических построениях.
В первую очередь это относится к магнию и стронцию, которые присутствуют в карбонатных скелетах постоянно, причем в относительно больших количествах. Кроме того, ясно и их место в минеральной фазе скелетного вещества: как и в неорганически осажденном карбонате кальция они могут замещать кальций в кристаллической решетке минерала. В связи с этим и подтверждая это, установлено, что в органогенных кальцитах содержание магния выше, чем в органогенных арагонитах, тогда как в содержании стронция картина обратная. Пожалуй, единственным исключением является состав кальцитовых ростров некоторых белемнитид, в котором количество магния соответствует его содержанию в "нормальном" кальците, а содержание стронция достигает значений, характерных для "нормального" арагонита. Причиной этого могут быть либо особенности физиологии белемнитид, позволяющие им образовывать кальцит с аномально высоким содержанием стронция, либо особенности диагенеза ростров данной группы организмов.
К настоящему времени опубликовано большое количество работ, в которых анализируется и интерпретируется содержание магния в карбонатных скелетах разных групп организмов. Эти интерпретации преследуют две цели, как и палеобиогеохимические исследования вообще: выяснение таксономической специфичности и зависимости содержания данного элемента от факторов внешней среды. Вместе с тем в очень немногих работах приводятся результаты комплексных исследований и еще меньше таких работ, где анализируются закономерности диагенетического перераспределения магния в скелетных остатках.
Таксономическая специфичность. Содержание магния в кальцитовых скелетах беспозвоночных колеблется в широких пределах (от 12% до десятых долей процента). В арагонитовых скелетах пределы содержания магния существенно меньше. Высокомагнезиальный кальцит (содержание магния > 3%), который, в сущности, представляет собой специфическую минеральную фазу, встречается в скелетах иглокожих. В челюстном аппарате морских ежей установлено самое высокое содержание магния, что заставляет предполагать присутствие протодоломита, т. е. существенных изменений в кристаллической решетке минерала. Если рассматривать данные о содержании магния в совокупности по крупным филумам беспозвоночных, но можно отметить, что в целом по средним содержаниям для них характерна определенная специфичность, хотя внутри каждого из типов наблюдаются значительные колебания. При сравнении представителей, скелеты которых обладают одной и той же минералогической формой карбоната (низкомагнезиальный кальцит, высокомагнезиальный кальцит, арагонит), можно констатировать наличие тенденции к понижению содержания магния и повышению высоты организации группы.
Если рассматривать само присутствие магния, замещающего ионы кальция в решетке кальцита и арагонита, как "примесь" или как неизбежную статистическую "ошибку" при формировании "чистого" минерала, то отмеченная тенденция может трактоваться, по предложению Ю. Одума, как следствие совершенствования обменных процессов, усиления внутреннего гомеостаза и как "прогресс" биологического минералообразования. Вероятное филогенетическое следствие - выработка критериев примитивности - продвинутости эволюционного процесса. Уровень специфичности при применении этого критерия у разных групп неизвестен. Имеются данные по близким видам планктонных фораминифер, свидетельствующие о том, что виды-потомки отличаются от предкового вида пониженным содержанием магния.
Наиболее последовательные попытки изучения вариаций содержания магния и стронция в целях таксономии и филогении с построением на этой основе вариантов биогеохимической классификации проведены на мезозойских и кайнозойских брахиоподах [564, 759]. По биогеохимическим характеристикам внутри типа выделяются две группы: фосфатная и карбонатная. В свою очередь фосфатная группа подразделяется на протеиново-фосфатную (отряд Lingulida) и протеиново-фосфатно-карбонатную (отряд Discinida) подгруппы, а карбонатная - на карбонатную с низким (<1%) содержанием магния (отряд Rhynchonellida, преобладающее большинство представителей отряда Terebratulida) и карбонатную с высоким (>1%) содержанием магния (отряд Craniida, надсемейство Thecideoidea и семейство Megatheridae, условно относимые к отряду Terebratulida) подгруппы.
Сопоставление данных спектрального анализа раковин в филогенетических рядах некоторых групп юрских, меловых и палеогеновых замковых брахиопод позволило выявить определенные тенденции в изменении их химического элементарного состава, в частности понижение содержания магния и повышения содержания стронция [563, 612].
Приведенными весьма общими сведениями в настоящее время, по-видимому, исчерпываются возможности использования информации о содержании магния для таксономии и филогении, которая может быть получена с помощью сравнительно простых аналитических методов. Дальнейшее развитие этого направления потребует иных методов исследований, которые позволят получать более точные и воспроизводимые результаты, а также непременной статистической обработки.
Многие группы как современных, так и ископаемых скелетных организмов остаются пока не изученными даже на этом первом, аналитическом уровне.
Содержание магния, стронция и других элементов в скелетах и факторы внешней среды. Остановимся прежде всего на так называемом "кальций-магниевом методе" палеотермометрии, исходной посылкой которого служат общие соображения о положительной корреляции содержания магния в карбонатных скелетах морских беспозвоночных с температурой воды, основанные на эмпирических наблюдениях, согласно которым в скелетах тропических форм содержание магния относительно выше, чем в скелетах холодноводных организмов.
Начало применению этого метода было положено работой Т. С. Берлин и А. В. Хабакова [79], в которой показана корреляция данных, полученных по изотопам кислорода и по кальций-магниевому отношению в проанализированных образцах, приведена первая эмпирическая формула для пересчета на температуру водной среды. Простота и экспрессивность рассматриваемого метода благоприятствовали его применению для анализа массового материала. Этому способствовал и упоминавшийся выше кризис в изотопной палеотермометрии.
Исследования кальций-магниевого отношения получили в нашей стране заметное развитие. Были, в частности, выполнены палеоклиматические реконструкции для обширных пространств юга СССР в юре, в мелу и палеогене [823, 1245]; в работе [823] этот метод использован для определения температурных условий обитания современных морских моллюсков.
В то же время ни термодинамических расчетов, ни успешных экспериментальных наблюдений, показывающих характер зависимости и возможности использования кальций-магниевого метода пока нет. Не проводилось и серьезного его обсуждения, остаются неясными приемы, с помощью которых удается получать правдоподобные значения палеотемператур по различным скелетным остаткам и часто по единичным их образцам. В целом для дальнейшего применения данного метода необходимо получить его однозначное теоретическое и экспериментальное обоснование.
Аналогичное магнию положение с использованием для таксономических, филогенетических и палеоэкологических реконструкций складывается и в отношении стронция. Особенности геохимического поведения стронция в биогенных карбонатах изучались, пожалуй, больше, чем магния. Результаты неоднократно подвергались обобщению и обсуждению, пересматривались основные заключения [173, 1226, 1510, 2022, 2399]. Большой научный резонанс вызвали работы Х. Лоуэнстама [1898], изучавшего в сравнительном аспекте распределение стронция в раковинах современных и ископаемых брахиопод и гастропод. Он пришел к выводам, что начиная с карбона отношение Ca/Sr в раковинах брахиопод оставалось постоянным и может использоваться для анализа изменений содержания стронция в океанах. X. Лоуэнстам полагал также, что концентрация стронция в водах океана в течение указанного времени также была неизменной.
В другой работе X. Лоуэнстама [1899] приведены данные сравнительного изучения отношения Ca/Sr в арагонитовых раковинах каменноугольных, меловых и современных гастропод. По его данным, содержание стронция в этих раковинах с течением времени уменьшалось. Это трактовалось как проявление биогеохимической эволюции, имевшей ту же направленность, что и общая тенденция к дискриминации элементов-примесей у более эволюционно развитых групп. Сходная тенденция была обнаружена при изучении арагонитовых раковин цефалопод [1697]. Все эти данные были подвергнуты проверке и по другим материалам. При этом были получены как неопределенные, так и противоположные результаты, не подтверждающие указанную тенденцию. Так, проверка идеи о "стронциевой эволюции", проведенная на обширной выборке раковин меловых и кайнозойских (в том числе и ныне живущих) гастропод и пелеципод [2120], показала, что если у арагонитовых раковин эта тенденция присутствовала, то у кальцитовых, наоборот, количество стронция с течением времени возрастало. Установленные различия авторы расценили как результат диагенетических процессов, считая диагенетическим артефактом и саму "биогеохимическую эволюцию".
При исследовании распределения Mg, Sr, Fe, Mn, Ba в неперекристаллизованных раковинах неогеновых, ископаемых четвертичных и современных моллюсков было замечено, что коррелятивные связи в содержании названных химических элементов у ископаемых и современных форм одного и того же вида существенно неодинаковы. Таким образом, палеоэкологические реконструкции на основе биогеохимических данных по перечисленным элементам-примесям должны проводиться с большой осторожностью [1476, 2449]. Очевидно, что это предостережение должно относиться к использованию в таксономических и филогенетических построениях и других элементов-примесей.
Выводы, основанные на изучении химического элементарного состава ископаемых скелетных остатков, не могут считаться доказательными и заслуживающими доверия, если они опираются только на общие соображения и не подкреплены специальным рассмотрением особенностей диагенеза и других вторичных изменений анализируемого материала и оценкой степени достоверности исходных данных.
Наиболее изучено в диагенетическом процессе, по-видимому, поведение стронция [1224]. Следует внимательно отнестись к идее автора указанной выше работы о существовании "закрытых систем" карбонатного диагенеза, в которых может сохраняться первичное соотношение элементов.
Время "простых" палеобиогеохимических исследований, некритического валового набора аналитических данных и выводов, основанных на общих соображениях, прошло (это, однако, не снимает с повестки дня проведение таких работ вообще).
Дальнейшие исследования могут развиваться по нескольким направлениям, среди которых в первую очередь нужно назвать фундаментальные экспериментальные и теоретические исследования, которые позволили бы сформулировать модель, объясняющую пути поступления элементов-примесей в скелетную ткань, определяющую их место и форму присутствия в минерально-органическом комплексе биогенных минералов. Работы должны проводиться в комплексе с экспериментальными исследованиями процесса биоминерализации. В настоящее время сделаны лишь первые попытки в этом направлении. Здесь необходимо овладение методами экспериментальных биохимических исследований и базой для их проведения.
Другая важнейшая область исследований - изучение геохимических закономерностей поведения элементов-примесей в процессе диагенеза. Это должно привести к созданию новых методических основ палеобиогеохимии, к пониманию того, что могут и чего не могут дать аналитические материалы по составу скелетообразующих минералов. Судя по результатам исследований распределения малых элементов в скелете различных видов современных Nautilus, выполненных самыми совершенными методами, межвидовые различия в их содержании чрезвычайно невелики [1902]. В связи с этим необходимо весьма тщательное изучение как химических [1476], так и микроструктурных [2201] изменений в анализируемом материале.
Вместе с тем, по-видимому, остается рациональным и продолжение региональных палеобиогеохимических работ с набором больших массивов корректных аналитических данных при условии четкой постановки задач, объективной оценки степени достоверности аналитических результатов с обязательной их статистической обработкой. Не состав единичной раковины, а групповой показатель, характеризующий целое захоронение, "популяцию", должен стать основой исходных данных. Такой показатель позволит правильно оценить характер диагенеза [1476]. Совокупный анализ состава элементов-примесей и изотопного состава кислорода позволит оценить степень диагенетического воздействия и стать критерием сохранности.
Несомненный интерес для реконструкции кислородного режима в палеоморях представляет предложенный в последние годы метод изучения распределения редкоземельных элементов (РЗЭ) в ископаемых фосфатных скелетных остатках (зубах, костях, конодонтах). Метод основывается на том, что относительное содержание легких и тяжелых лантаноидов существенно зависит от условий кислотно-щелочности среды, через которую проходит содержащий их минерал.
В водах, богатых кислородом, относительное содержание церия и лантана ("цериевая" аномалия) меньше, чем в бескислородной обстановке. Исследование распределения РЗЭ в кальций-фосфатном веществе конодонтов, чешуи и зубов рыб по всему разрезу фанерозоя позволило показать [2504] общие тенденции в изменении "цериевой" аномалии и соответственно кислородного режима в морях геологического прошлого (рис. 82).
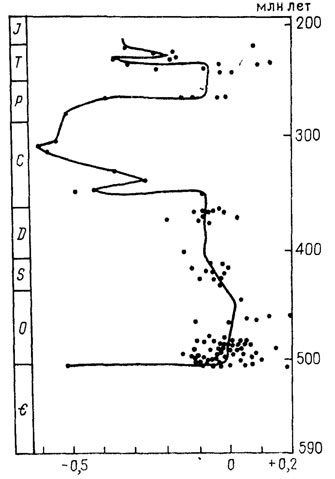
Рис. 82. Схема изменения 'цериевой аномалии' (Сеа пот = log 3 - Сеn 27Lan + Ndn), вычисленной по содержанию редкоземельных элементов в апатитовом материале скелетных остатков аппарата конодонтов из выборок разного геологического возраста. По Дж. Райт. Более отрицательные значения (сдвиг кривой влево) отвечают окислительным условиям среды захоронения остатков; положительная аномалия свидетельствует об аноксидных условиях
Палеобиохимические исследования. Изучение органической составляющей скелета ископаемых беспозвоночных начало интенсивно развиваться после того, как П. Абельсон* [1208, 1251, 1252] показал сохранение в скелетных остатках различных организмов достаточно больших количеств органического вещества. От него это направление и получило наименование палеобиохимического (палеобиохимии).
* (Нередко транскрибируема с английского как Эйбельсон.)
Органическое вещество скелета представлено в основном протеинами, гликопротеинами и полисахаридами (обычно хитином). Органическое вещество распределяется между кристаллическими слоями, между отдельными кристаллами и внутри их (см. 2.1, 2.2). Такая "упаковка" скелетного органического вещества защищает его от непосредственного воздействия разрушающих факторов среды и способствует сохранению в ископаемом состоянии. Это в свою очередь открывает возможности изучения ископаемых остатков биохимическими методами, применяемыми к исследованию современных форм, что вселило надежды на установление особенностей биохимической эволюции организмов и на ископаемом материале. Осознание этих перспектив стимулировало изучение закономерностей образования минерализованного скелета, вылившееся в новое комплексное научное направление, - исследования по проблемам биоминерализации (см. 2.1, 2.2).
В многочисленных работах 60-х и начала 70-х годов нашего столетия, выполненных в основном на моллюсках, было показано, что органическое вещество скелета определяет нуклеацию, рост, минералогическую форму, структурное и морфологическое оформление минеральной фазы. Органический матрикс скелета многокомпонентен, полное количество его фракций в скелете и их состав до конца еще не известны. В нем присутствуют ЭДТА-растворимая и ЭДТА-нерастворимая части, каждая из которых в свою очередь неоднородна. Первоначально полагали, что наиболее информативной и лучше сохраняющейся является ЭДТА-нерастворимая фракция. Именно она и привлекла в первую очередь внимание исследователей.
Были проведены многочисленные работы по изучению нерастворимой фракции ЭДТА методами аминокислотного анализа (бумажной и газовой хроматографии после гидролиза), электрофореза и инфракрасной спектроскопии на многих группах ископаемых и современных беспозвоночных [51]. Наиболее целенаправленные исследования принадлежат Е. Т. Дегенсу с сотрудниками [1490, 1491, 1492]. Изучение современных и кайнозойских моллюсков позволило выявить определенные эволюционные закономерности в аминокислотном составе нерастворимой фракции, построить "биохимические древа" для исследованных форм. Были выдвинуты некоторые общие положения биохимической эволюции. В частности, по Е. Т. Дегенсу, расхождение филумов моллюсков и членистоногих от общего аннелидного предка выразилось в увеличении протеиновой фракции у моллюсков и полисахаридной (хитиновой) - у членистоногих. Была предложена биохимическая модель [1927], объясняющая "взрывное" появление минерализованных скелетов в начале кембрия освобождением взаимосвязанных боковых цепей полярных аминокислот и присоединением к ним катионов и анионов, образовавших кристаллическую решетку минеральной фазы. В результате детальных электронно-микроскопических исследований нерастворимой фракции различных слоев раковин моллюсков была установлена их морфологическая специфичность для разных классов данного типа беспозвоночных [1446, 1667, 1668, 2437].
Однако достаточно скоро стало ясно, что ни электронно-микроскопические морфологические данные, ни результаты аминокислотного анализа нерастворимой фракции, ни изучение общего аминокислотного состава органического вещества скелета организмов не дают той информации, которую можно было бы последовательно использовать в таксономических, эволюционных и палеоэкологических построениях. Было показано, что "палеопротеины" скелета быстро и сильно изменяются после смерти организма и его захоронения. Количество аминокислот и аминосахаров очень быстро уменьшается, а в качественном их составе происходит утеря той незначительной видоспецифичности, которая установлена у современных форм (среди скелетных коллагенов современных позвоночных из 1000 аминокислотных остатков видоспецифичен состав и расположение в полипептидной цепи лишь 20 аминокислот). В течение уже первых тысячелетий после захоронения состав органического вещества (если он не был изменен до захоронения) на уровне валового состава аминокислот становится совершенно одинаковым у самых отдаленных организмов, вариации состава не коррелируют ни с систематикой, ни с геологическим возрастом. При этом процесс изменения состава является самопроизвольным внутренним "автопроцессом", который был назван М. Флоркеном "палеоизацией" [1586], чтобы отличать его от процесса диагенеза, обусловленного влиянием внешней среды.
По-видимому, закономерным результатом палеоизации является образование меланоидных комплексов, которое "уравнивает" состав палеопротеинов независимо от их происхождения. В таком виде органическое вещество становится устойчивым, сохраняется неопределенно долго, но непригодно для целей таксономии и филогении.
Другое следствие палеоизации скелетных протеинов - рацемизация входящих в их состав аминокислот, которая может, вероятно, происходить и без нарушения полипептидной цепи. Аминокислоты, образованные живыми организмами, находятся исключительно в L-форме. Но уже в течение десятков тысячелетий часть из них переходит в D-форму, свойственную "неживым" аминокислотам. Этот самопроизвольно протекающий процесс рацемизации имеет линейную направленность, что привело к созданию на его основе метода абсолютных возрастных датировок, который "работает" в пределах, по меньшей мере, голоцена, где он может дополнять и конкурировать с радиоуглеродным методом [1268, 1289].
Очевидно, следует согласиться с общим выводом о том, что нерастворимое органическое вещество скелетных остатков ископаемых во всех случаях "палеоизовано" и не соответствует первоначальному органическому веществу. Его изучение методами аминокислотного анализа не может дать результатов для прослеживания таксономической специфичности и биохимической эволюции. На процесс палеоизации накладываются, кроме того, изменения, вызванные условиями захоронения, диагенеза и катагенеза вмещающих пород. Органическое вещество ископаемых остатков скелета может, таким образом, быть важным показателем преобразования пород и его изучение имеет важное значение в геохимии органического вещества [340].
Новые перспективы палеобиохимических исследований открылись совершенно неожиданно и были связаны с изучением органических компонентов ЭДТА-растворимой фракции скелета и применением более совершенной аналитической техники. Работами П. Вестброка и Е. В. де Йонга с сотрудниками и других специалистов [1399, 1485, 1597, 2468] в экстрактах из ЭДТА-растворимой фракции скелетных остатков позднемеловых белемнитов, аммонитов, бивальвий (пелеципод) и яиц динозавров было обнаружено присутствие компонентов, которые сохранили антигенные свойства. Это означало, что исследователи получили в руки ископаемые протеиновые молекулы, сохранившие биологическую активность, т. е, полную исходную структуру и конформацию. Применение обычных и усовершенствованных для анализа наноколичеств вещества иммунологических методов [1903] позволяет проводить полное сравнение молекул различных групп ископаемых и современных форм. Имеются данные о том, что эти протеины обладают выраженными кальций-связывающими свойствами, что заставляет предполагать их непосредственное участие в биохимических механизмах биоминерализации. Такие протеины (сравнительно небольшой молекулярной массы) выделены из раковин современных и ископаемых устриц и из яиц динозавров.
Эти исследования, несмотря на трудности выделения и очистки исчезающее в малых количеств вещества, интенсивно развиваются. Показано достаточно большое разнообразие рассматриваемых протеинов, часть из которых обладает иммунными свойствами, а часть принадлежит к группе Ca-связывающих протеинов; начаты работы по расшифровке первичной последовательности аминокислот в полипептидной цепи этих протеинов с помощью самой совершенной (секвекторы) молекулярно-биологической техники. Трудности, стоящие перед исследователями, образно сравниваются с работой по расшифровке текста на порванной и смятой странице, испачканной и склеенной битумообразными продуктами палеоизации. Однако эти трудности на современном этапе развития молекулярно-биологических методов очистки и микроанализа представляются разрешимыми. Перспективы использования ископаемых остатков для прослеживания эволюции на молекулярном уровне обнадеживающие.
Комплексное изучение ископаемых остатков представителями разных научных дисциплин и разнообразными методами становится насущной потребностью палеонтологии, а палеонтологические материалы и построения входят в сферу интересов самых передовых научных отраслей, что поднимает палеонтологию на уровень экспериментальных наук.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'