
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
2.7. Молекулярная палеонтология (И. Е. Лейфман, А. Н. Гусева, В. П. Макридин)
Идея о расширении рамок классической палеонтологии была высказана в 1923 г. одним из основоположников биогеохимии Я. В. Самойловым, который писал: "Наша современная палеонтология в своих обширных и глубоких достижениях базируется на ископаемых морфологических остатках организмов, я мыслю палеонтологию биохимическую, которая открывала бы нам существование организмов (может быть, совершенно не известных нам морфологически) исключительно по их биохимическим особенностям..." [907, с. 118]. Эта мысль Я. В. Самойлова значительно опережала свое время и не могла быть практически реализована из-за отсутствия точных методов исследований, но она оставила заметный след в науке и способствовала развитию не только органической геохимии и палеонтологии, но и палеофизиологии (см. 1.6).
Возникновение молекулярной палеонтологии связано с именем лауреата Нобелевской премии М. Кальвина. В статье "Молекулярная палеонтология" [1402], а затем в монографии "Химическая эволюция" [1403, русский перевод - 460] им были сформулированы основные положения молекулярной палеонтологии как направления исследований биогенных органических соединений ("молекулярных ископаемых") в докембрийских породах, лишенных морфологических палеонтологических остатков. Очевидно, что молекулярная палеонтология возродила на современном, качественно значительно более высоком уровне научно-технического прогресса предвидение Я. В. Самойлова.
Современные успехи биоорганической химии и хемотаксономии организмов, широкое изучение органических соединений в осадочных породах и в составе горючих ископаемых, выяснение закономерностей фоссилизации и последующих преобразований захороненных биомолекул привели к расширению круга объектов молекулярной палеонтологии и областей ее применения.
Предпосылки возникновения молекулярной палеонтологии. Вторая половина XX в. характеризуется бурным развитием наук, занимающихся изучением вещества на молекулярном уровне. Ярким выражением этого явилось возникновение в середине 50-х годов молекулярной биологии - новой области естествознания, приведшей к революции в познании живой природы.
Молекулярная биология проложила дорогу новому подходу к изучению явлений жизни - исследованию элементарных биологических процессов на уровне молекул биополимеров и предельно простых систем [42]. Успехи молекулярной биологии в изучении процессов жизнедеятельности на уровне молекулярных структур и молекулярных взаимодействий [63] доказали биохимическую общность всех организмов и единство их происхождения, функционирование на молекулярном уровне не только управляющих организменных и биоценотических, но и эволюционно-генетических систем сложных организмов.
Переход на молекулярный уровень познания органических соединений в геологических объектах связан как с развитием методов идентификации органических соединений, так и с переносом достижений в выяснении химического состава биологических объектов и процессов их функционирования из неонтологии в палеонтологию. Подобную тенденцию отмечал еще в 1917-1918 гг. Я. В. Самойлов, который считал, что "настало время для расширения палеонтологических исканий в сторону раскрытия прошлой жизни в ее биохимических проявлениях. Палеонтология биохимическая (физиологическая), естественно, должна пользоваться и другими методами изучения, чем палеонтология морфологическая, а именно - геохимическими методами" [907, с. 91].
Предвидение Я. В. Самойлова оправдалось лишь в 50-х годах нашего столетия, когда внедрение современного молекулярного Уровня в исследования ископаемых остатков организмов привело в первую очередь к обособлению в качестве самостоятельной научной дисциплины палеобиохимии, возникшей на стыке палеонтологии, биохимии, молекулярной биологии и геохимии органических соединений.
Термин "палеобиохимия" был предложен Я. В. Самойловым [907]. Хотя исследования в этой области начались еще в 1908 г. [501], современный уровень в палеобиохимии был достигнут благодаря классическим работам Ф. Абельсона, опубликованным в 1954-1961 гг. В качестве двух основных направлений в палеобиохимии Ф. Абельсон [1] наметил рассмотрение сравнительной биохимии современных организмов с палеонтологической точки зрения и изучение состава органических веществ в ископаемых остатках организмов и в углеродсодержащих породах.
По определению Ч. М. Колесникова, "...в самом широком понимании палеобиохимия - это биохимия живого вещества биосферы Земли минувших геологических эпох. Конечной целью палеобиохимии в таком ее понимании является стремление изучить на молекулярном уровне происхождение и закономерности эволюции живого вещества, а также оценить влияние его на ведущие процессы, протекающие в биосфере Земли в течение геологической истории" [501, с. 9].
Развитие палеобиохимии привело на рубеже 60-х годов к обособлению в качестве самостоятельных научных направлений органической геохимии [1] и биогеохимии органического вещества [1805], а в конце 60-х годов - к обособлению молекулярной палеонтологии [1402], хотя и после этого длительное время молекулярная палеонтология рассматривалась как одно из направлений палеобиохимии [51, 501, 1925] и геохимии органического вещества [620].
Предмет, объекты и методы молекулярной палеонтологии. М. Кальвин так определил предмет молекулярной палеонтологии: "...в породах, более древних, чем докембрийские, морфологических остатков не обнаружено, и поэтому нам придется иметь дело с "молекулярными остатками", по структуре которых мы можем судить об их происхождении" [460, с. 17].
Развернутую характеристику молекулярных ископаемых привел М. Руттен [895]: "Молекулярные ископаемые - это молекулы биологического происхождения" (с. 191), "...химически распознаваемые остатки биологических соединений" (с. 203), "...это более устойчивые фрагменты крупных органических молекул, ранее входивших в состав организма, а затем распавшихся в процессе фоссилизации" (с. 192). Следует подчеркнуть, что М. Руттен отмечает распознаваемость молекулярных остатков организмов на химическом уровне.
Видимо, начало работ по выделению и идентификации из геологических объектов соединений, имеющих аналоги в живой природе среди биомолекул, связано с именем А. Трайбса, обнаружившего в 1934-1935 гг. порфирины в нефтях и горных породах. Биомолекулы и их геохимические дериваты, идентифицированные в различных геологических объектах, получили самые разные наименования, отражающие, как правило, различные аспекты геологической судьбы биомолекул. Так, Д. Фокс [1600] назвал каротиноиды, выделенные из морских осадков, "biochemical fossils". Ф. Абельсон [1252] именовал аминокислоты, выделенные из пород, "fossil organic compounds". Название "biological markers" впервые встречается в работе Дж. Эглинтона с соавторами [1777], a "chemical fossils" - в статье Э. Баргхорна, У. Майншайна и Дж. Шопфа [1303] и в обзоре М. Кальвина [1401]; в статье М. Руттена [2195] этот термин использован в несколько ином написании - "chemofossil". M. Кальвин впервые употребил термин "molecular fossils" [1401]. Ал. А. Петров [817] применил название "реликтовые соединения". Наконец, следует упомянуть термин "geochemical fossils", использованный М. Тиссо и Д. Вельте [2387; русск. перевод - 1059].
Перечисленные наименования отражают разный подход к информации, которую несут ископаемые биомолекулы и их геохимические дериваты. Так, термин "biological markers" (в русском переводе - биологические маркеры, метки или метчики, биологические индикаторы, биологически маркирующие) передает взгляд на биомолекулы и их геохимические дериваты как на соединения, характеризующие следы жизнедеятельности в породах различного возраста, особенно докембрийского. Другой аспект - унаследованность структуры от биомолекул - отражен в названии "реликтовые соединения", "реликты", хотя лишь часть из них может считаться в полном смысле истинными реликтами: неизмененными, полностью сохранившимися биомолекулами. Наконец, в названиях "biochemical fossils" (биохимические ископаемые), "chemical fossils" (в русском переводе - химические ископаемые, хемофоссилии), "molecular fossils" (молекулярные ископаемые) и "geochemical fossils" (геохимические ископаемые) выражен историко-генетический подход, т. е. участие биомолекул в геохимических процессах захоронения исходного биологического материала и его сохранность в геологических объектах на химическом (молекулярном) уровне. Все эти названия определяют ископаемые органические соединения как предмет молекулярной палеонтологии, но вместе с тем отражают судьбу биомолекул, вырванных геохимическими процессами из биотического круговорота углерода в те или иные эпохи существования биосферы, т. е. имеют четко выраженный биогеохимический характер.
Место "молекулярных остатков" как предмета палеонтологии можно определить в сравнении с традиционно изучаемым палеонтологическим материалом. Согласно В. В. Друщицу [342], материалом для палеонтологии служат любые сохранившиеся в слоях Земли остатки организмов или следы их жизнедеятельности, называемые окаменелостями, или фоссилиями. Поэтому последовательность морфологически различимых фоссилий "макрофоссилии - микрофоссилии - нанофоссилии" естественно дополнить хемофоссилиями - остатками органических соединений организмов, распознаваемыми на химическом молекулярном уровне [286].
Термин "хемофоссилии" предпочтительнее упомянутых выше терминов, поскольку показывает не только химический уровень сохранности и распознаваемости ископаемого биологического материала, но и роль химических изменений исходных биомолекул.
Молекулярная палеонтология фиксирует "молекулярные остатки" в тех или иных геологических объектах. В современной палеонтологии молекулярные ископаемые, или хемофоссилии, рассматриваются как одна из форм сохранности палеонтологических объектов, как следы жизнедеятельности организмов прошлых геологических эпох. В связи с этим молекулярную палеонтологию можно рассматривать не только как раздел палеобиохимии, изучающий преимущественно докембрийскую эволюцию, но и как раздел палеонтологии, анализирующий остатки организмов на молекулярном уровне в осадочных породах любого возраста (не только в докембрийских). До настоящего времени объем понятия "молекулярная палеонтология" еще не определился, о чем свидетельствует статья Б. Раннегара "Molecular palaeontology", опубликованная в журнале "Palaeontology" в 1986 г. Б. Раннегар отнес к молекулярной палеонтологии такие палеобиохимические, по существу, направления исследований, как изучение органических молекул в остатках ископаемых организмов, биоминерализация, изучение филогении белков.
Вместе с тем уже достаточно ясно может быть выражена специфика молекулярной палеонтологии, связанная с рассмотрением молекулярных ископаемых как палеонтологического материала, несущего обычную палеонтологическую информацию (например, об организмах прошлого и палеобиоценозах, об условиях накопления осадков и т. д.). Молекулярные ископаемые позволяют опознать на химическом уровне как метаболиты, так и молекулярные остатки различных организмов палеобиоценозов (от автотрофов до гетеротрофов-деструкторов), уцелевшие в процессах биодеструкции и захоронившиеся в осадках. Поэтому можно считать, что в самом широком понимании молекулярная палеонтология изучает молекулярные следы жизнедеятельности организмов прошлых геологических эпох с целью суждения по молекулярным остаткам об эволюции живого вещества и биосферы.
Методы исследований "молекулярных ископаемых" отличаются от традиционных для палеонтологии методов и в настоящее время базируются на достижениях газовой хроматографии и масс-спектрометрии, особенно компьютерной хроматомасс-спектрометрии и пиролитической техники в комплексе с другими современными методами молекулярного анализа [784, 1059]. При современном уровне техники аналитических работ хемофоссилии - биологически маркированные соединения - могут быть выделены и определены даже в тех случаях, когда материнские организмы полностью распались, а органические вещества рассеялись, и их содержание во вмещающих породах, возраст которых насчитывает сотни миллионов и даже более 2,5 миллиардов лет, крайне невелико.
Специфика исследования молекулярных ископаемых позволяет, по мнению И. С. Барскова, считать, что "исследования в области молекулярной палеонтологии лежат вне собственно палеонтологии и являются, скорее, отраслью химии и геохимии органических веществ" [51, с. 6]. Это отражает расширение рамок классической палеонтологии по предмету и методам исследований, сближение ее с органической геохимией - современной крупной отраслью геохимии.
Образование хемофоссилий. Хемофоссилии, по Н. Б. Вассоевичу, А. Н. Гусевой и И. Е. Лейфману [149], - это ископаемые биомолекулы, их опознаваемые фрагменты и генетические аналоги, являющиеся составной частью разновозрастных и разнофациальных осадков, пород и горючих ископаемых.
Хемофоссилии выясненной структуры в составе ископаемых остатков организмов, осадков, пород и горючих ископаемых относятся к самым разным классам органических соединений - углеводородам, кислородным и азотным соединениям, как низко-, так и высокомолекулярным. Составу хемофоссилий посвящена обширная литература. Особо следует отметить статьи в журнале "Geochimica et cosmochimica Acta" и в трудах Международных конгрессов по органической геохимии, обобщения в монографиях "Органическая геохимия" [784], Б. Тиссо и Д. Вельте [1059] и Дж. Ханта[1121].
Хотя в современных осадках выявлены хемофоссилии, генетически связанные со всеми классами биосоединений (углеводами, белками и другими азотсодержащими соединениями, лигнинами и родственными ароматическими соединениями, липидами и родственными соединениями), особая роль принадлежит соединениям липидно-липоидной природы. Эти соединения - биосинтезированные производные жирных кислот (липиды) и изопреноидов (липоиды) - содержат мало функциональных групп, вследствие чего сохраняют при разных способах фоссилизации особенности своей молекулярной структуры. Выяснение условий образования, сохранности и эволюции хемофоссилий как ископаемых органических соединений изначально биогенного генезиса [284, 286, 571, 572] показало, что существование и сохранность хемофоссилий обусловлены особенностями молекулярной структуры биомолекул, условиями их фоссилизации и последующего преобразования в стратосфере.
Еще В. И. Вернадский отмечал, что "нужно... принимать во внимание особые химические свойства углерода, между которыми наиболее характерным является устойчивость его органических природных соединений в термодинамических земных условиях. Они изменяются в них лишь очень медленно..." [164, с. 141]. Это связано с тем, что основные элементы молекулярной структуры органических соединений - углеродный скелет, стереохимическая конфигурация, функциональные группы и другие - устойчивы в широком диапазоне условий осадочной толщи. Следствием устойчивости углеродного скелета является сохранение органическими соединениями молекулярной индивидуальности в различных агрегатных состояниях, в том числе в растворах (например, при миграции), что коренным образом отличает их от неорганических соединений.
Молекулярная структура биомолекул обусловлена их физиологической ролью в живых организмах и определяет поведение биомолекул в пищевых цепях экосистем. Направленность изменений биомолекул вне синтезировавших их организмов связана с их трофической ценностью, и поэтому состав биомассы, подвергающейся фоссилизации в субаквальных осадках, в значительной степени определяется интенсивностью потребления первичной биопродукции гетеротрофами - консументами и деструкторами. Ими в первую очередь используются первичные вещества, такие, как белки, запасные углеводы, основные компоненты жиров (триглицериды кислот и сложные липиды), способные вовлекаться в основной обмен веществ самых различных организмов. Благодаря этому происходит избирательное накопление разнообразных вторичных веществ в пищевых цепях гетеротрофов. Это касается как низкомолекулярных соединений (н-алканы, 2- и 3-метилалканы, сложные эфиры восков, стероиды, терпеноиды и др.), так и биополимеров (структурные углеводы, лигнины, кутаны, суберины, спорополленины и др.), выполнявших в растениях опорные, защитные и регуляторные функции.
Закономерности поведения биомолекул при фоссилизации и на последующих этапах литогенеза изучаются молекулярной биогеохимией - разделом общей биогеохимии, рассматривающим в качестве предмета исследования геохимическую историю биомолекул [150, 152, 571, 572].
Фоссилизация органического вещества представляет собой биогеохимический процесс, широко распространенный в геологическом времени и пространстве и связанный с общим характером биосферных процессов.
Уход биомолекул - основной формы нахождения атомов углерода в живом веществе - из биотического круговорота углерода путем захоронения в осадочных отложениях связан с тем, что этот процесс является одним из звеньев в механизме саморегулирования биосферы, определяющем уровень содержания кислорода в атмосфере Земли. Как писал В. И. Вернадский, "...углерод своим выходом из жизненного цикла создает и поддерживает в механизме биосферы явление огромного первостепенного значения, так как только этим путем создается возможность для существования в биосфере соответствующих масс свободного кислорода..." [164, с. 208]. Это положение получило подтверждение в работах Н. Н. и Н. Н. Верзилиных [161], О. П. Добродеева [319] и др.
Н. М. Страхов [1019] показал, что для сохранения и фоссилизации органического вещества субаквальные условия более предпочтительны, чем субаэральные, что обусловлено исключительной ролью воды как среды протекания процессов, приводящих к фоссилизации органического вещества. Благоприятные для фоссилизации биомолекул условия наступают при высокой обводненности в тех осадках, в которых благодаря жизнедеятельности прокариот-деструкторов создаются условия анаэробиоза, способствующие сохранению части биомолекул от биогенной деструкции. Фоссилизация биомолекул в субаквальных осадках происходит в водных (пресноводных и морских) и некоторых наземных (гидрофации болот) экосистемах. Геологическими процессами биомолекулы в осадках выводятся за пределы породивших их экосистем, включаясь в круговорот углерода в стратисфере.
Фоссилизация органического вещества осуществляется молекулярными механизмами фоссилизации - совокупностью биохимических, химических и коллоидно-сорбционных процессов, которые, обеспечивая ограничение биогенной деструкции, способствуют консервации захороняющихся биомолекул и их вовлечению в состав образующихся осадков [572].
Переход органического вещества в ископаемое состояние происходит на двух уровнях - надмолекулярном (тафономическом) и молекулярном [285]. На тафономическом уровне в субаквальные осадки уходят морфологически различимые остатки организмов, не разрушенных в силу своей биохимической устойчивости (например, захороненные споры, обрывки кутикул и пробковой ткани высших растений) или в результате прекращения деструкционных процессов при неблагоприятных условиях среды (например, остатки древесины и корней растений в торфах, остатки фито- и зоопланктона в осадках зон апвеллинга и зон сероводородного заражения придонных вод). На молекулярном уровне в осадки переходят продукты распада некромы: во-первых, путем фоссилизации продуктов деструкции биополимера посредством геосинтеза (за счет включения в состав образующихся геополимеров), во-вторых, путем фоссилизации других низкомолекулярных биомолекул, в основном бывших вторичных метаболитов, с сохранением величины молекул (в виде индивидуальных хемофоссилий - неизмененных биомолекул и их геохимических дериватов).
Основные типы химических превращений в зонах седименто- и диагенеза способствуют сохранению углеродного скелета исходных биомолекул [149]. Поэтому хемофоссилии - диагенетические дериваты биомолекул наследуют разнообразные молекулярные структуры биосоединений. Это касается и макромолекул геополимеров, структурными единицами которых являются биомолекулы или их фрагменты.
Эволюция хемофоссилий. Возможность сохранности хемофоссилии в осадках и затем в породах определяется тенденцией изменений органических соединений в направлении образования продуктов, наиболее устойчивых к окружающей физико-химической обстановке.
Условия существования осадка и превращения его в породу (седименто- и диагенез) благоприятны для сохранения многих хемофоссилий любое достаточно длительное геологическое время, особенно в случаях хорошей изолированности от внешних воздействий. На это обратил внимание В. И. Вернадский [164, с. 141], писавший, что "в термодинамических и химических условиях земной коры все природные углеродистые соединения устойчивы, если защитить их от влияния жизни. Соединения, образующие живое вещество - белки, жиры, углеводы, остаются неприкосновенными и изменяются лишь крайне медленно, не утрачивая своего строения при условии изолированности от явлений жизни".
Подобная изолированность характерна, например, для ископаемых скелетных остатков организмов, в которых сохраняются молекулярная структура и конформация белков [51]; для торфов, содержащих практически неизмененными многие компоненты исходных растений - от биополимеров (углеводы, лигнины, кутины, суберины) в морфологических остатках до низкомолекулярных соединений (н-алканы, эстеры восков, стероиды, терпеноиды и др.), запечатанных в захороненных защитных восках и смолах растений. Но это не мешает развитию самопроизвольно протекающих химических реакций типа диспропорционирования водорода. Эти реакции претерпевают, например, непредельные циклические терпеноиды. Так, первичные смоляные кислоты и абиетиновая кислота (основной компонент смоляных кислот хвойных) превращаются в ретен и фихтелит - минералы, характерные для болотных отложений (рис. 83).
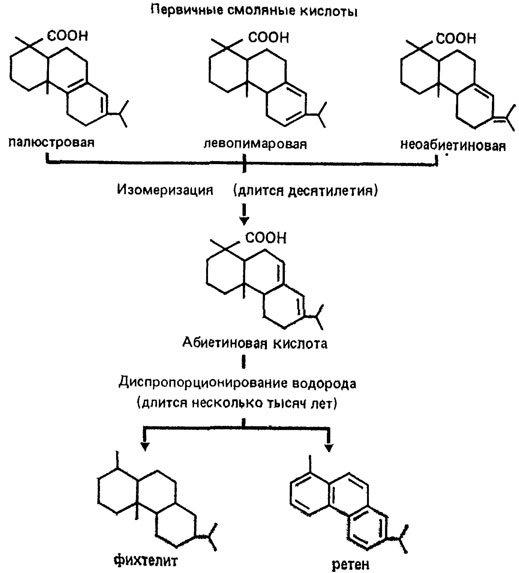
Рис. 83. Превращения смоляных кислот в погребенной сосновой древесине в торфяной залежи. По А. И. Скригану. Указанные химические превращения протекают самопроизвольно с образованием более устойчивых соединений (вначале абиетиновой кислоты, затем углеводородов) и завершаются геологически достаточно быстро, формируя своеобразные органические минералы - ретен (шерерит) и фихтелит, структура которых сохраняет углеродный скелет исходных соединений
Закономерности преобразования хемофоссилий в условиях осадочной толщи наиболее подробно изучены на примере горючих ископаемых. Известно, что в условиях диагенеза постепенно исчезают унаследованные биополимеры, например лигнины и углеводы в торфах, превращающиеся в геополимеры - гуминовые кислоты. В бурых углях (условия протокатагенеза) отмечена высокая сохранность различных биомолекул липидно-липоидного генезиса, наследующих особенности молекулярно-массового распределения этих соединений в исходных растениях (н-алканы, жирные кислоты, эстеры восков и др., например в богемских углях с возрастом 30-50 млн лет [2499]). Сохраняются частично химически активные центры (непредельные связи, функциональные группы) и даже стереоконфигурация исходных биомолекул, например в пентациклических тритерпеноидах рядов олеанана и гопана [278]. Вместе с тем продолжаются процессы диспропорционирования водорода, прослеживающиеся в рядах ароматизации тритерпеноидов, например при превращениях бетулина - основного компонента коры березы (рис. 84). В этих преобразованиях сохраняется скелет исходных биомолекул, что позволяет установить последовательность их трансформации.
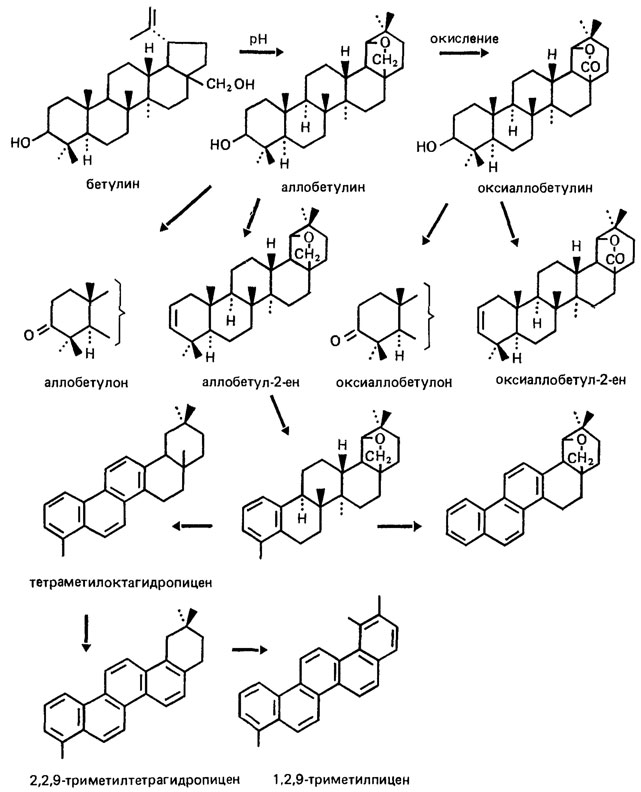
Рис. 84. Бетулин (Betula и родственные растения) и продукты его превращения, обнаруженные в богемских бурых углях с возрастом 30-50 млн. лет. По Н. Г. Голованову, М. Штрейблу и М. Героуту. Эволюция исходной химически активной биомолекулярной структуры идет в направлении образования более устойчивых форм; возможная последовательность трансформации показана стрелками
Начиная с подзоны мезокатагенеза, повышенные температуры и катализ приводят к существенной эволюции состава хемофоссилий в направлении образования "нефтяных" форм. Это проявляется в изменении молекулярно-массового распределения хемофоссилий (например, н-алканов), в появлении новых стереоизомерных форм (например, "нефтяных" стеранов и гопанов вместо биологических) в каменных углях (начиная с газовых) и горючих сланцах [279]. Все это происходит вследствие развития процессов деструкции макромолекул фоссилизированного органического вещества, особенно их липидных и липоидных составляющих.
Если же действию повышенных температур подвергается рассеянное органическое вещество, то могут развиваться процессы термокаталитического синтеза новообразований - соединений нефти с качественно новым типом углеродного скелета, несвойственным биомолекулам. Этим отличаются многие соединения нефти и в их числе хемофоссилий, включающие лишь как фрагменты структуры детали углеродного скелета биомолекул. Разнообразие хемофоссилий значительно уменьшается, и уже хемофоссилиям нефти свойственна ограниченность наследуемых типов углеродного скелета биомолекул. В основном это неразветвленные длинные углеродные цепи и цепи типа 2- и 3-метилалканов, изопреноидные цепи, циклы стероидов и терпеноидов и некоторые другие [149]. Для хемофоссилий нефти типичны продукты деструкции различных биомолекул - жирных кислот, фитола, терпеноидов, каротиноидов, стероидов, сохраняющие опознаваемость в качестве фрагментов определенных биомолекул (рис. 85).
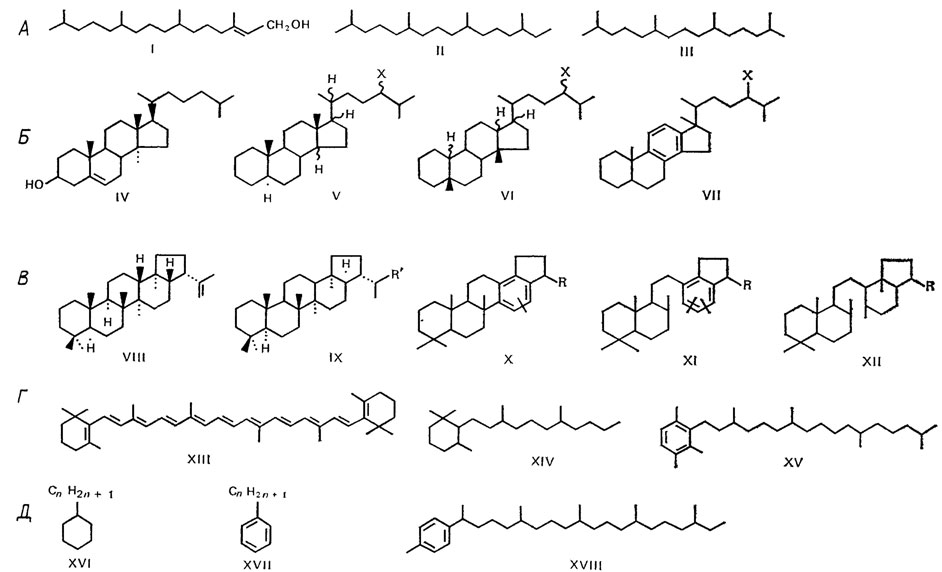
Рис. 85. Структура некоторых хемофоссилий нефти, связанных происхождением с различными группами липидов и липоидов живых организмов. По Ал. А. Петрову. А - фитол (I) и найденные в нефти фитан (II) и пристан (III); Б - холестерин (IV) и найденные в нефти стераны (V): α-стераны (14 αН, 17αH-'биологические') и изостераны (14 βН, 17 βН-'нефтяные'), перегруппированные (структурно-измененные) стераны (VI) и моноароматические стераны (VII); В - 17 βН, 21 βH-гопен - 22 (29) (VIII), или диплоптен (сине-зеленые водоросли, бактерии) и найденные в нефти 17 αН, 21 βН-гопаны (IX), моноароматические гопаны (X), моноароматические 8-14-секогопаны (XI) и 8-14-секогопаны (XII); Г - β-каротин (XIII) и найденные в нефти углеводороды каротиноидного типа строения 1,1,3-триметил-2-алкилциклогексаны С10-С22 (XIV) и 1,3,4-триметил-2-алкилбензолы С10-С30 (XV); Д - возможные продукты циклизации н-алкановых и изопреноидных кислот, найденные в нефти: н-алкилциклогексаны (XVI), н-алкилбензолы (XVII), 1-метил-4-алкилбензолы С12-C31 с изопреноидной цепью (XVIII)
Отмеченные закономерности геохимической истории биомолекул позволяют говорить о направленной эволюции хемофоссилий как о процессе изменения их молекулярной структуры после фоссилизации в ходе литогенеза содержащих их отложений и расширении состава и набора хемофоссилий в историческом аспекте - от древнейших докембрийских образований до современных [286]. Они отражают реальные процессы индивидуального развития хемофоссилий под влиянием изменения условий окружающей среды и эволюции растительного мира как основного поставщика захороняющихся биомолекул.
Понятие об эволюции хемофоссилий важно потому, что хемофоссилий нельзя считать "химическими окаменелостями", т. е. сохраняющимися неизменными в ископаемом состоянии. Напротив, хемофоссилий как органические соединения способны к тем химическим превращениям, которые вызваны сменой условий их существования по мере литогенеза вмещающих пород. При этом возможны довольно существенные изменения хемофоссилий как по сравнению с исходными биомолекулами, так и при переходе от диагенетических к катагенетическим дериватам биомолекул. Особенно резкие изменения структуры хемофоссилий связаны с зоной катагенеза, с нефтеобразованием.
Несмотря на все эти изменения, "узнаваемость" хемофоссилий как соединений изначально биогенной природы и как биомаркеров сохраняется достаточно долгое время независимо от степени литогенеза вмещающих отложений. Даже хемофоссилий зоны катагенеза, претерпевающие постепенное обеднение биогеохимической информацией, все же сохраняют ее достаточно длительное время, вплоть до перехода содержащих их пород в метаморфиты. Таковы, например, докембрийские отложения, сохранившие такие хемофоссилий, как н-алканы, фитан и пристан в образованиях возраста 2,7-3,1 млрд лет [460].
При переходе от древнейших отложений, содержащих достаточно малый набор хемофоссилий - биомаркеров сине-зеленых водорослей (н-алканы, 7- и 8-метилалканы, фитан и пристан, гопаны и некоторые другие), к более молодым отложениям увеличивается разнообразие типов хемофоссилий, отражающих появление новых биосоединений по мере эволюции органического мира. Основные этапы изменения химического состава растительного мира, как и основные тенденции изменения химического состава различных биосоединений в ходе эволюции растений, прослеживаются достаточно четко [573]. Общим является возникновение новых типов биосоединений при появлении эукариотов, при выходе растений на сушу, при появлении голосеменных, а затем и покрытосеменных растений. Для наземных растений характерно все увеличивающееся разнообразие вторичных веществ, особенно у покрытосеменных растений. Эти вещества - биохимически наиболее стойкие компоненты растений - в первую очередь подвергаются фоссилизации в субаквальных условиях и сохраняются в виде хемофоссилий в осадочных породах. Хемофоссилий сохраняют на молекулярном уровне хемотаксономические особенности организмов, что важно при сопоставительном анализе хемотаксономии современных организмов и палеохемотаксономии.
Интенсивное развитие хемотаксономии за последние два десятилетия определяется той новой информацией, которая дополняет традиционные классификации организмов. "Хемосистематические (или более узко понимаемые хемотаксономические) исследования как новое направление в получении информации о таксонах (видах, родах, семействах и др.) способствуют расширению базиса классификации" [1067, с. 321].
В хемотаксономии бактерий широко используют жирно-кислотный состав липидов [706]. У растений в качестве таксономических различий рассматривают наличие и распределение самых разнообразных биомолекул: аминокислот и ДНК, углеводов, фенольных соединений, алкалоидов, длинноцепочных соединений жиров, масел и восков, изопреноидов - терпеноидов, каротиноидов и стероидов. Основное значение имеют вторичные вещества растений. Типичные хемотаксономические различия в растениях существуют на всех уровнях иерархий - от высших таксонов до семейств, родов и видов. Таковы, например, наличие гопаноидов у прокариотов и некоторых эукариотов, а стероидов - у эукариотов [2060]. Распределение н-алканов в водорослях и бактериях сильно отличается от их распределения в высших растениях, содержащих н-алканы С23-С35 с сильным преобладанием нечетных гомологов [1699].
Некоторые хемотаксономические различия существуют на уровне типов. Так, 1- и 8-метилгептадеканы характерны только для сине-зеленых водорослей и отсутствуют в других типах бактерий, зеленых водорослях и высших растениях; пристан и фитан присутствуют у анаэробных нефотосинтезирующих и фотосинтезирующих бактерий и высших растений и отсутствуют у аэробных нефотосинтезирующих бактерий, сине-зеленых и зеленых водорослей [1699]. Все типы водорослей-эукариотов различаются набором и основными компонентами стеролов [2072] и каротиноидов [1881]. На уровне классов хемотаксономические различия проявляются, например, в различии терпеноидов эфирных масел голосеменных и покрытосеменных растений [9]; на уровне подклассов - в различии алкалоидов однодольных и двудольных покрытосеменных растений. Отмечены различия в жирно-кислотном составе жиров и в составе стероидов и каротиноидов на уровне гипов и семейств растений; известна специфичность кутикулярных восков, кутинов, суберинов, лигнинов у разных таксонов наземных растений и др. С эволюцией растений сильно увеличивается специфичность их химизма, достигая у покрытосеменных хемотаксономических различий на уровне семейств, родов и даже видов. Это фиксируется, например, в строении и составе жирных кислот жиров [1073].
Консервативность хемотаксономических признаков растений и других организмов отражает сохранение их химизма на протяжении сотен миллионов лет, о чем свидетельствует идентичность молекулярных ископаемых - остатков ископаемых организмов - и биомолекул тех же организмов, живущих в настоящее время. М. Кальвин [460] приводит в качестве примера способность сине-зеленых водорослей - как современных, так и древних (из сланцев свиты Соуден, возраст 2,67 млрд лет) - синтезировать 7- и 8-метилгептадеканы C18H38. Отмечена идентичность современных и ископаемых (возраст 3,4-3,7 млрд лет) спорополленинов [120].
В этой связи Дж. Эглинтон и М. Кальвин [1207, с. 206] высказали мысль, что "...возможность классифицировать живущие на Земле виды на хемотаксономической или химической основе... можно будет распространить... на вымершие организмы, создав науку, которую можно будет назвать палеохемотаксономией". Придавая огромное значение данным палеохемотаксономии для суждения об эволюции живого на Земле, Э. Брода даже отождествляет палеохемотаксономию с молекулярной палеонтологией. "Можно надеяться, - пишет он, - что... на основе химической палеолетописи будут найдены критерии для классификации разных организмов: иными словами, возникает новая наука - палеохемотаксономия, или молекулярная палеонтология" [119, с. 239].
Информативность хемофоссилий. Хемофоссилии несут информацию, которая может быть названа биогеохимической [284]. Эта информация отражает (сохраняет) структуру оригинала, т. е. биомолекул, в разной степени измененных геохимическими процессами, и позволяет воспроизвести структуру оригинала- исходных биомолекул - по их ископаемым дериватам. Наследование хемофоссилиями биогеохимической информации связано с сохранением углеродного скелета, стереохимических особенностей и других элементов молекулярной структуры биомолекул. Наследование этих элементов и характер их изменения в геохимических процессах обеспечивает сохранение двух видов биогеохимической информации - об исходных организмах и о процессах геохимических трансформаций биомолекул.
Наследование хемофоссилиями информации об исходных организмах базируется на сохранении на молекулярном уровне хемотаксономических особенностей организмов, зафиксированных в наборе и концентрации определенных биомолекул. Часть хемотаксономических особенностей наследуется низкомолекулярными хемофоссилиями (н-алканы, стероиды, терпеноиды, воски и др.), часть - структурными единицами геополимеров, например остатками жирных кислот [493] и других биомолекул, из которых построены макромолекулы горючих сланцев и других сапропелитов. Поэтому особенности хемофоссилий, в том числе и геополимеров, позволяют говорить об исходных организмах, которым могли принадлежать биомолекулы, ставшие хемофоссилиями после захоронения.
Интерпретация хемофоссилий как индикаторов условий осадконакопления, диагенеза и катагенеза вмещающих отложений подробно обсуждается Б. Тиссо и Д. Вельте [1059]. Она базируется на сопоставлении состава и набора хемофоссилий, степени измененности их структуры по сравнению с исходными биомолекулами, характере новообразований (диагенетических или катагенетических дериватов).
Сохранение хемофоссилиями биогеохимической информации определяет их уникальность как молекулярных следов жизнедеятельности в породах различного возраста, отражающих существование биосфер прошлого, запечатленных в истории осадочных пород. Хемофоссилий в этой связи могут характеризовать эволюцию биосферы в геологической истории Земли и, что особенно важно, различные этапы докембрийской эволюции. Уникальным объектом, содержащим хемофоссилий, являются горючие ископаемые, поэтому изучение хемофоссилий может осветить многие аспекты эволюции горючих ископаемых в истории Земли.
Молекулярная палеонтология докембрия. Последние полвека ознаменовались принципиальным сдвигом в разработке одной из самых фундаментальных проблем естествознания - проблемы происхождения и ранних этапов эволюции жизни на Земле, решительным отходом от умозрительных, нередко фантастических представлений к научному проникновению в сущность сложнейших процессов, протекавших на заре развития нашей планеты. Успехи эти связаны прежде всего с именем А. И. Опарина [778, 779, 780, 782 и др.], создавшего материалистическую теорию самоорганизации материи и положившего начало многочисленным исследованиям, развившим ее краеугольные положения [81, 219, 317, 482, 625, 828, 895, 1112, 1209, 1964 и др.].
Проблема происхождения и ранних этапов развития жизни на Земле чрезвычайно обширна, многогранна и ее разработка требует комплексного подхода. Она привлекает внимание многих естественных наук - органической химии, органической геохимии, молекулярной биологии, биохимии, астрономии, астрофизики, вулканологии, микробиологии, палеонтологии и других, каждая из которых рассматривает определенные ее аспекты. В частности, относящиеся к палеонтологии аспекты освещены в работах Б. С. Соколова [984, 985, 989, 990, 993]. Нуждается данная проблема и в философских обобщениях. Приводимое ниже схематическое изображение хронологической последовательности основных событий в докембрийской эволюции (рис. 86) позволяет получить самое общее представление о месте и роли ряда названных наук в ее познании. Совершенно очевидно, например, что предполагаемый этап химической эволюции не мог бы найти отражения в традиционной палеонтологической летописи, ведущая роль в изучении которой принадлежит наукам, выясняющим механизм и закономерности естественного синтеза веществ определенного состава, в первую очередь белков, послуживших субстратом для перехода от органических соединений абиогенного генезиса к биологической форме материи. При исследовании же этапа биологической эволюции и появлении в земных слоях все более многочисленных, разнообразных и достоверных остатков организмов значение морфологической палеонтологии быстро возрастает (см. 7.2).
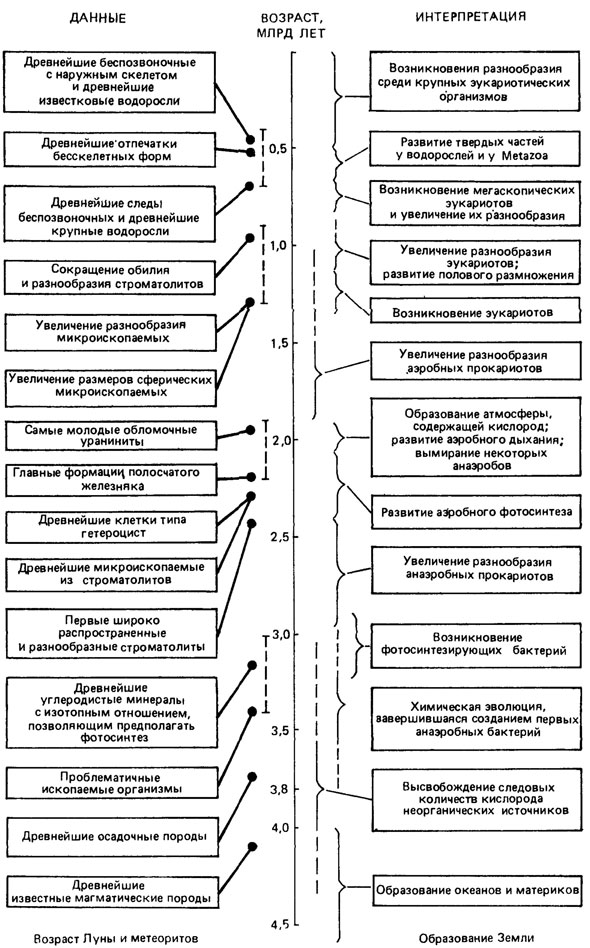
Рис. 86. Хронологическая последовательность главных событий в докембрийской эволюции по данным палеонтологии, петрографии и геохимии, сравнительного изучения метаболизма и биохимии современных организмов. По Дж. У. Шопфу
Молекулярная палеонтология первоначально сосредоточивала свое внимание на выяснении структуры органических молекул, сохраняющихся в докембрийских и архейских породах, и их эволюции. "Наша конечная цель, - писал М. Кальвин, - состоит в том, чтобы найти границу раздела между небиологическими предшественниками, или субстратами жизни и биологическими остатками жизни. Нам хотелось бы установить тот период времени, когда органические вещества небиологического происхождения существовали одновременно с веществами, образованными живыми структурами" [460, с. 40]. Сложившаяся, у него обобщающая картина изображена на рис. 87. Приводим также дополняющие и уточняющие ее данные (табл. 6).
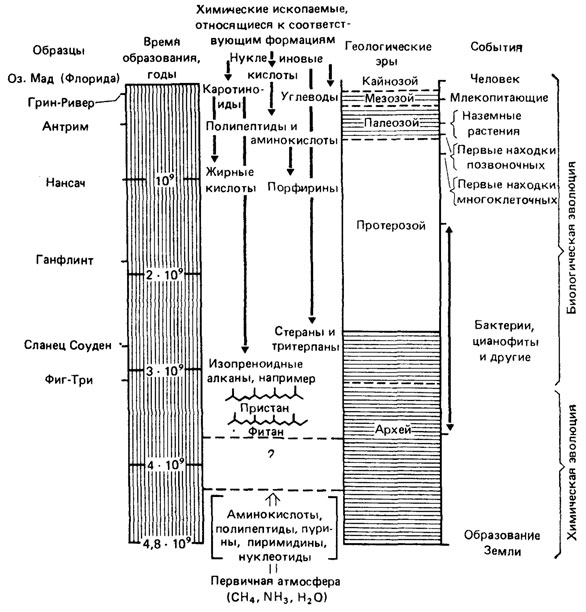
Рис. 87. Геохронологическая таблица химической и биологической эволюции. По М. Кальвину
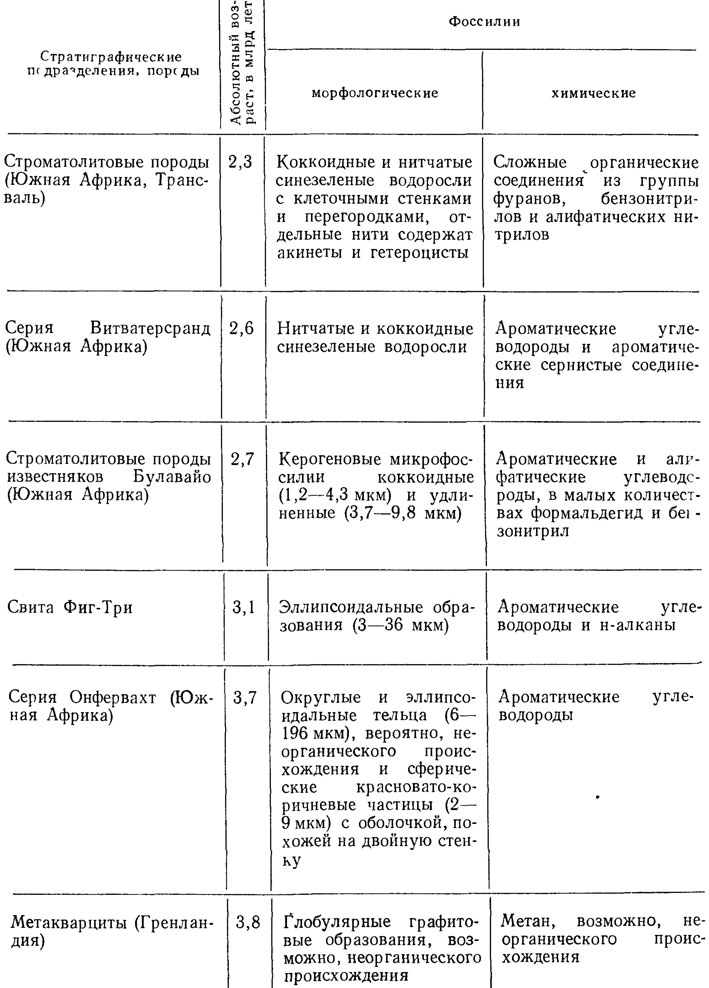
Таблица 6. Химические и морфологические ископаемые, обнаруженные в древнейших породах. По М. Кальвину, Дж. Шопфу и др.
В первично осадочных (осадочно-метаморфических) породах докембрия органическое вещество биогенного происхождения сохранилось в разных формах: рассеянное органическое вещество пород, горючие сланцы и другие обогащенные углеродом породы (например, шунгиты, битуминозные толщи) и нефти. Для познания докембрийской истории исключительный интерес представляют приуроченность и состав этих образований.
Сопоставляя результаты изучения углеводородов, выделенных из докембрийских сланцев разного возраста, М. Кальвин [460] отметил, что в отложениях Онфервахт (возраст 3,7 млрд лет) молекулярно-массовое распределение н-алканов С16-С25 настолько отличается от распределения в более молодых породах, что можно ставить вопрос об этом органическом веществе как смеси абиогенных и биогенных компонентов, т. е. о его приуроченности к переходу от химической к биологической эволюции. Однако Дж. Эглинтон [784] указывает, что подобный однородный ряд н-алканов могут дать биогенные н-алканы под воздействием процессов дегидрогенизации и графитизации органического вещества докембрийских пород. Равномерное распределение н-алканов С14-С25 характерно также для кремнистого сланца Фиг-Три (возраст 3,1 млрд лет), но концентрации н-алканов здесь значительно выше, чем в образце из Онфервахт [460].
В более молодых докембрийских сланцах Соуден (2,7 млрд лет), Ганфлит (1,9 млрд лет) и Нансач (1,0 млрд лет) распределение н-алканов, фитана, пристана и других изопреноидов не оставляет сомнения в их биогенном происхождении. В ряде случаев эти соединения могут считаться биомаркерами. Например, в сланцах Соуден идентифицирован изоалкан C18 (типа 7- или 8-метилгептадекана), биосинтезированный сине-зелеными водорослями Nostoc [460].
Можно отметить, что молекулярные ископаемые докембрийских пород подтверждают тезис В. И. Вернадского [165, с. 93]: "...органическое вещество, проникающее все вещество доступной изучению земной коры, все - биогенного происхождения".
В докембрии выделено несколько (6-8) эпох накопления толщ, богатых органическим веществом, имеющих возраст от 3,5 млрд лет до начала фанерозоя [741, 949]. Эпохи накопления богатых органическим веществом сапропелевых осадков проявляются через 200-400 млн лет и приурочены к породам различного возраста: AR1 (3,7-3,5 млрд лет), AR2 (3,4-3,1 млрд лет), AR2-PR1 (2,7 млрд), PR1-PR2 (1,9-1,8 млрд), PR2-PR3 (1,6-1,5 млрд), R1-R2 (1,3-1,2 млрд), R2-R3 (1,0 млрд), V (0,57 млрд) [741].
Возрастные "уровни" накопления этих толщ связывают с важными событиями в эволюции биоса [948] и с основными глобальными эпохами регионального метаморфизма и тектономагматической активизации, в которых шло накопление углекислоты в древних атмосферах, что могло отразиться на эволюции биоса [949].
Основными продуцентами органического вещества в раннем архее считаются анаэробные бактерии, а начиная с позднего архея - фотосинтезирующие сине-зеленые водоросли и акритархи. Экстремальная биопродуктивность бедных в видовом отношении примитивных водорослей в эпохи сланценакопления при подавленности или высокой изменчивости других групп организмов рассматривается как ответная реакция биосферы на повышение в водной среде обитания концентраций фосфора, урана и других тяжелых металлов [74].
Молекулярная палеонтология и горючие ископаемые. Традиционная проблема влияния исторического развития биоса на состав твердых горючих ископаемых обсуждается в трудах Г. Потонье, А. Н. Криштофовича, В. И. Вернадского, Ю. А. Жемчужникова, Н. М. Страхова, Л. Ш. Давиташвили и других ученых. Выяснение возрастной специфики состава углей на уровне особенностей ботанического состава растений палеозоя и мезозоя, характерное для исследований Г. Потонье [836] и Ю. А. Жемчужникова [366], переходит на современном этапе познания состава твердых горючих ископаемых на уровень изучения элементарных составляющих (микрокомпонентов) углей и горючих сланцев.
Еще В. И. Вернадский [166] высказал общее положение, что в каустобиолитах видно ясное проявление эволюционного процесса. Им же [164, с. 170] было отмечено наличие взаимосвязи состава нефтей и исходных организмов: "нефти происходят из определенных живых организмов, из живого вещества - определенного химического состава, определяющих структуру нефтей". Роль изучения горючих ископаемых для развития палеонтологии четко сформулирована В. Е. Раковским [853, с. 51]: "отложения угольного вещества и нефти являются важным памятником процессов разных геологических эпох; химический состав этих горючих ископаемых позволит в будущем пролить свет на состав растений прежних геологических эпох".
Угли и горючие сланцы. Возрастная специфика микрокомпонентов углей подробно рассмотрена в работах А. В. Лапо [561, 562], Е. М. Файзуллиной и А. В. Лапо [1088]. Подчеркнуты различия, существующие между равноуглефицированными микрокомпонентами групп витринита, лейптинита и фюзинита разного возраста - от палеозойского до кайнозойского - по элементному составу, выходу летучих и смолы полукоксования, т. е. на уровне общей химической характеристики. По данным ИК-спектрометрии показано повышенное содержание алифатических групп в витринитах каменных углей мезозойского возраста в сравнении с равноуглефицированными углями палеозоя, что связывается с эволюцией углеобразующей растительности [1088]. Можно полагать [573], что эти различия микрокомпонентов обусловлены эволюцией структурных звеньев биополимеров углеобразующей флоры. Однако недостаточная изученность химического состава микрокомпонентов углей, о чем свидетельствует сводка А. В. Лапо [562], не дает пока оснований проверить это предположение на молекулярном уровне.
Интересные данные о возрастной специфике горючих сланцев получены при изучении нерастворимой части их керогенов [493, 1083 и др.]. Поскольку горючие сланцы не подвергались в своей геологической истории деструктивному действию повышенных температур, остатки жирных кислот в составе макромолекул керогенов сохраняют специфику исходных жирных кислот. Так, черты сходства в органическом веществе докембрийских и нижнепалеозойских горючих сланцев объясняются [1083] отсутствием в сравниваемые эпохи резких изменений как в составе исходного для керогенообразования материала, так и в процессах осадконакопления. Вместе с тем длина алифатических цепей - фрагментов макромолекул керогена, т. е. "связанной" формы хемофоссилий, по А. Н. Гусевой и И. Е. Лейфману [573], - отражает специфику исходных жирных кислот водорослей (например, ордовикские кукерситы - сине-зеленых водорослей, юрские горючие сланцы - диатомовых водорослей), а в случае сноса терригенного материала - и жирных кислот наземных растений в формировании горючих сланцев и сапропелитов мезозоя и кайнозоя [493].
Нефти. Молекулярная палеонтология, как и молекулярная биогеохимия, изучает нефть как объект, содержащий хемофоссилий [151].
Нефти аналогично ископаемым остаткам организмов, осадкам, породам и другим горючим ископаемым содержат много углеводородов и неуглеводородных соединений, относящихся к хемофоссилиям. Согласно Ал. А. Петрову [818, с. 41-42], всего в настоящее время в нефти определено до 400 индивидуальных углеводородов состава до С40 разных классов и родов, причем "...большинство из определяемых в нефтях углеводородов являются реликтами, т. е. соединениями, явно сохранившими черты строения, свойственные их биологическим предшественникам". Это в первую очередь углеводороды - нормальные алканы, изопреноидные алканы, стераны и тритерпаны (особенно гопаны), несущие в своей структурной и пространственной организации явные черты исходных соединений живого вещества, таких, как н-алканы, длинноцепочные жирные кислоты, фитол, стероиды, каротиноиды и гопаноиды. Ряд кислородсодержащих соединений - кислоты, кетоны, спирты - это производные стероидов, гопаноидов и каротиноидов, также несущие явные черты исходных биомолекул. Большая группа соединений нефти, включая типичные для нефтей гибридные углеводороды и компоненты смол и асфальтенов, содержит элементы структуры биомолекул в качестве фрагментов сложных молекул, образовавшихся как продукты синтеза при нефтеобразовании и не идентичных биомолекулам.
Состав хемофоссилий нефти представляет исключительный интерес для выяснения генезиса вещества нефти*. Это связано с тем, что в структуре хемофоссилий заключена информация об их происхождении, т. е. возможно установление генетической связи с биомолекулами-предшественниками. Весь имеющийся материал однозначно свидетельствует о доминирующей роли биосоединений изначально липидно-липоидной природы как исходного вещества в нефтеобразовании, подтверждая правильность теории осадочно-миграционного происхождения нефти. В свете новейших данных нефтеобразование представляет собой длительный стадийный процесс, берущий начало, как это отметил В. И. Вернадский, еще в живом веществе и завершающийся в осадочных породах, содержащих органические вещества, на этапе мезокатагенеза [146].
* (Генезис скоплений нефти до настоящего времени является предметом интенсивных исследований. Сложность природных процессов миграции пока не позволяет однозначно ответить на многие вопросы формирования и переформирования скоплений нефти.)
В хемофоссилиях нефти заключена как хемотаксономическая информация об организмах, составляющих исходный биос палеобассейна осадконакопления, т. е. будущих нефтематеринских толщ, так и информация об условиях геохимической трансформации биомолекул в нефть и о геохимической истории самой нефти. По существу, все показатели состава нефтей, используемые в последнее время при корреляции нефтей между собой и с органическим веществом пород, основаны на количественных соотношениях таких хемофоссилий, как н-алканы, изопренаны (фитан, пристан), стераны, тритерпаны и некоторые другие.
С позиций молекулярной палеонтологии представляет интерес отражение следов жизнедеятельности различных организмов в составе хемофоссилий нефти. Б. Тиссо и Д. Вельте [1059, с. 353] считают, что хемофоссилий как биомаркеры "...могут быть использованы для характеристики, корреляции и восстановления условий осадконакопления так же, как и макро- и микрофоссилии, обычно используемые геологами". Они указывают, что хемофоссилий могут характеризовать основные типы ассоциаций организмов, участвовавших в образовании органического вещества осадков, на уровне вклада фитопланктона, высших растений и микробиальный переработки растительного материала.
Во многих случаях соотношение н-алканов нефти может отразить вклад высших растений (для них типичны гомологи С25-С33) и низших растений (бактерий и водорослей), для которых типичны гомологи С15-С19, часто резкий максимум для C17 [1059, 1699]. Как показано рядом исследователей [1334, 1449, 1721, 2104, 2132], высокопарафинистые нефти приурочены обычно к прибрежно-морским и континентальным фациям, где велик вклад наземной растительности, богатой кутикулярными восками. Считается, что соотношение фитан - пристан одинаково для нефтей, образовавшихся за счет сходного по составу органического вещества, причем доля пристана выше там, где существенней вклад зоопланктона. Такие типичные хемофоссилии, как гопаны, могут быть унаследованы нефтями от прокариотов - сине-зеленых водорослей и бактерий, участвовавших в переработке автотрофной продукции, хотя синтезируются и некоторыми эукариотами - лишайниками, папоротниками, некоторыми древесными растениями и травами [2060].
Вопросы отражения возрастной специфики хемофоссилиями нефтей достаточно сложны в силу значимости термокаталитических деструкционных процессов в образовании вещества нефти, роли миграции в формировании залежей нефти, в приуроченности залежей нефти к вмещающим отложениям, соотношение которых с предполагаемыми нефтематеринскими толщами может быть различным.
В обобщениях Н. Б. Вассоевича и Г. А. Аммосова [148], А. А. Карцева [476, 477], О. А. Радченко [852], X. Смита [970] и других ученых показаны тенденции изменения свойств и состава нефтей с возрастом вмещающих отложений (кайнозой - мезозой - палеозой). А. А. Карцев [476] по этим параметрам делит нефти на палеотипные и кайнотипные. По особенностям свойств и группового углеводородного состава нефти выделяют пять мегациклов нефтегазообразования (вендский, ранне- и средне-поздне-палеозойские, мезозойский и кайнозойский [785]). По изменению содержания реликтовых углеводородов (н- и изопреноидные алканы, тетра- и пентациклические цикланы) отмечены различия между нефтями кембрийских, девонских, пермских, мезозойских и кайнозойских отложений, что определяется в основном продуктами и условиями седиментации нефтепродуцирующих осадков и согласуется с эволюционными изменениями биосферы [915]. Вместе с тем пока, видимо, нельзя утверждать, что в нефти есть хемофоссилии-реликты, которые четко характеризуют возраст исходной биомассы.
Использование хемофоссилии нефти для реконструкции истории нефтеобразования и геохимической истории нефти в залежи подробно рассмотрено Б. Тиссо и Д. Вельте [1059]. Согласно этим авторам (с. 353), "хотя идентификация хемофоссилии часто требует значительных усилий, получаемые результаты окупают их. Они являются совершенным инструментом при корреляции различных нефтей или нефтепроявлений, обнаруженных в осадочных бассейнах в результате нефтепоисковых работ. Если различные типы нефтей уже выделены, хемофоссилии могут быть использованы для того, чтобы связать эти нефти с соответствующими материнскими породами".
Становится все более ясным, что палеонтологические и биогеохимические исследования на молекулярном уровне конкретизируют многие положения осадочно-миграционной теории образования нефти и газа. Эти исследования можно считать молекулярной основой учения о генезисе нефти, поскольку в структуре ископаемых молекул заложена информация об их происхождении и геохимической истории. Большое практическое значение исследований вытекает из того, что поиски нефти все больше ведутся на историко-генетической основе, в связи с чем возрастает и значимость той информации, которая сохраняется самим веществом нефти.
Вопросы интерпретации данных о молекулярных ископаемых. В связи со все возрастающей ролью информации, которая заключена в хемофоссилиях, в палеонтологии, в изучении возникновения и эволюции биосферы, в геохимии и биогеохимии горючих ископаемых особое значение приобретают вопросы интерпретации данных о молекулярных ископаемых.
По своему характеру химические ископаемые весьма существенно отличаются от ископаемых морфологических остатков организмов, являющихся традиционными объектами палеонтологических исследований.
Естественно, что дальнейшее развитие молекулярной палеонтологии связано прежде всего с совершенствованием аналитических методов и решением специальных методических проблем изучения хемофоссилий. Основополагающими из них следует считать две проблемы: 1) о сингенетичности хемофоссилий вмещающим породам и их вторичных изменениях и 2) о происхождении - биогенном либо абиогенном - хемофоссилий.
Наиболее важными хемофоссилиями для изучения эволюционных процессов, протекавших в архее и протерозое, считаются углеводороды (н-алканы, фитан, пристан), липиды (жирные кислоты) и пигменты (спорополленин, порфирины), обладающие наиболее высокой стабильностью и достаточно отчетливо выраженными особенностями структуры, отличающими их от других соединений. Имеются данные [2345] о находках в докембрийских водорослях Британской Колумбии "свободных", частично, по-видимому, первичных и "связанных" сахаридов. Не вполне ясным оказывается вопрос о поведении аминокислот. Ф. Абель-сон [1, 1253] привел данные в пользу химической и термической устойчивости некоторых из этих соединений, в частности аланина, и высказал мнение о возможности их сохранения в ископаемом состоянии длительное время. Согласно его представлениям, аланин и некоторые другие аминокислоты могут в течение миллиарда лет выдерживать анаэробные условия при температуре 25°С, при комнатной же температуре под действием кислорода они разрушаются за миллион лет. К сходным заключениям о сохранности аминокислот в древних породах, возможности их извлечения, исследования и идентификации пришли многие авторы [460, 945, 1403, 1580, 1586, 2466 и др.]. В частности, И. З. Сергиенко и М. И. Бобылева обнаружили в кианитовых сланцах свиты кейв (2,5 млрд. лет) различные "свободные" и "связанные" аминокислоты, а также углеводы - глюкозу и маннозу.
Согласно Т. В. Дроздовой [337, 338, 339, 621], "связанные" аминокислоты входят в состав высокомолекулярной части рассеянного органического вещества пород, будучи химически связаны в макромолекулах геополимеров.
Вопросы интерпретации данных о присутствии аминокислот в породах разного возраста и разных стадий литогенеза обсуждают Г. А. Лаврентьев и И. А. Егоров [557]. Они подвергают сомнению возможность сохранности нативных аминокислот в течение геологического времени в силу их способности к рацемизации и эпимеризации. Присутствие же аминокислот в докембрийских и более молодых, но метаморфизованных породах объясняются ими как следствие природной контаминации.
Другая группа молекулярных ископаемых - углеводороды "...относятся к наиболее стабильной группе соединений, и можно ожидать, что они сохранили существенные особенности своей первоначальной молекулярной структуры [460, с. 40]. По расчетам, приводимым М. Кальвином (там же), время жизни углеводородов при 300 К составляет 1025 лет, при 400 К - 1013,5 лет.
Интересный анализ проблемы возможности биогенного или абиогенного происхождения углеводородов в породах привел В. Г. Мейншейн [784]. Он отметил, что хотя органические соединения могут создаваться в результате анаболических или абиотических реакций, многие высокоселективные реакции непосредственно или косвенно связаны с жизнью, а "...основным недостатком большинства абиотических теорий происхождения соединений углерода в породах является, по-видимому, неспособность этих теорий предложить правдоподобный источник абиотических реагентов и катализаторов. Многие из найденных в породах сложных углеводородов достаточно устойчивы, чтобы сохранить структурную упорядоченность, характерную для биологических остатков... и эти вещества или их предшественники очень важны для выяснения биологической эволюции" [784, с. 232, 233].
Из всех биологических молекул, считает В. Г. Мейншейн, насыщенные углеводороды могут иметь наибольшее геохимическое значение; "...энергия разложения некоторых биологических алканов (алканы, согласно этому автору, - это все насыщенные углеводороды, т. е. алканы и цикланы по общепринятой номенклатуре органических соединений) (курсив наш. - И. Л., А. Г., В. М.) и присутствие алканов в породах практически всех возрастов подтверждает, что главная часть этих углеводородов может сохранять свою структурную целостность почти при любых условиях, имеющихся в осадочных отложениях. Таким образом, вопрос состоит, очевидно, не в том, являются ли алканы молекулярными остатками прежней жизни, а скорее в том, насколько или какая часть алканов в древних породах образовалась из алканов биологических липидов" [784, с. 245].
В этом плане важно то обстоятельство, что образование углеводородов при трансформации фоссилизированных кислородосодержащих компонентов липидной и изопреноидной природы является обязательным этапом геохимической истории этих компонентов [287]. В частности, новообразование н-алканов из жирных кислот (свободных или связанных в восках или макромолекулах керогена пород) изменяет молекулярно-массовое распределение унаследованных н-алканов. Изопреноидные алканы (фитан, пристан и др.), стераны, гопаны и другие терпаны обычно имеют своими предшественниками в живом различные кислородные производные (спирты и др.).
При оценке значимости молекулярных ископаемых следует учитывать, что при умеренных температурах и окислительно-восстановительных потенциалах соединения углерода подвергаются структурным изменениям, которые точно отражают воздействующие на них процессы [784]. Уникальность углеродных скелетов стероидов и терпеноидов (как циклических, так и алифатических) и сохранение ими стереохимической конфигурации, отвечающей биологической или близкой к ней (известно, что стереоконфигурация холестерина единственная из 256 возможных по числу хиральных центров), делает эти соединения, как и н-алканы, одними из основных среди молекулярных ископаемых разновозрастных и разнофациальных пород и горючих ископаемых.
Что касается син- и эпигенетичности тех или иных хемофоссилий, то многочисленные исследования показывают, что источник исключительно автохтонных хемофоссилий существует - это нерастворимая часть керогена пород. И. Дж. Галлегос [264] утверждает, что кероген, в котором содержится основная масса органического вещества осадочных пород, - это потенциальный источник незагрязненных биологических маркирующих соединений. Этот вывод важен в том отношении, что макромолекулы керогена пород формируются на стадии седиментогенеза, как показано в настоящее время на примере различных горючих сланцев [264, 493].
В макромолекулы керогена включены в виде структурных элементов биомаркеры тех организмов, остатки которых принимали участие в формировании осадков, давших породы, содержащие кероген. Методом выделения биомаркеров является пиролиз керогена, высвобождающий ранее фоссилизированные жирные кислоты в виде н-алканов, н-алкенов, кетонов, концентрационное распределение которых позволяет реконструировать начальное распределение жирных кислот [493] и изопреноиды - изопренаны, стераны, терпаны - в виде набора соединений, часть из которых, как показано К. Зейфертом, сохраняет стереохимию исходных биомолекул [2260].
Успехи органической геохимии все больше способствуют однозначности интерпретации данных о молекулярных ископаемых. В частности, остается все меньше доводов в пользу абиогенного синтеза хемофоссилий, все больше выясняется роль постфоссилизационных (абиогенных) преобразований захороненных биомолекул в формировании хемофоссилий пород и горючих ископаемых.
Несмотря на безусловную важность данных молекулярной палеонтологии для разработки проблемы происхождения жизни на Земле, задачи ее в целом представляются более широкими. Количество, разнообразие состава и геохимическая роль живого вещества в процессе биологической эволюции быстро возрастали. В связи с этим молекулярная палеонтология, охватывая своими исследованиями не только докембрий, но и фанерозой, поставляет весьма важную информацию об основных компонентах исходного живого вещества или продуктах их сравнительно неглубокого преобразования, без которой немыслимо изучение биохимии растений и животных геологического прошлого, проникновение во многие закономерности эволюции биосферы, выяснение происхождения и условий накопления горючих ископаемых. На этом пути молекулярная палеонтология вступает во взаимодействие с другими науками, изучающими органическое вещество - углехимией и углепетрографией, нефтехимией, химией горючих сланцев, биогеохимией и другими, сохраняя при этом присущие ей особенности. Прав был, вероятно, Н. Б. Вассоевич [147], утверждавший, что молекулярная палеонтология займет отчетливо выраженное положение между зарождающейся наукой об ископаемом органическом веществе - литогеохимией и палеонтологией.
Закономерности формирования химических ископаемых изучены еще недостаточно. В самом общем виде можно сказать, что их образование связано с фоссилизацией биологических молекул, сохранившихся после гибели материнских организмов, но утративших первоначальную связь с живым веществом и перешедших в ископаемое состояние. Под воздействием геохимических процессов в обстановках осадконакопления и диагенеза часть этих молекул превращается в устойчивые для данных условий соединения, сохраняющие полностью или фрагментарно углеродный скелет исходных биологических молекул и несущие информацию об их систематической принадлежности и путях эволюции.
Важное значение в реализации рассматриваемой идеи имеет разработка палеотаксономического направления исследований, занимающегося классификацией организмов геологического прошлого по их биохимическим особенностям и непосредственно связывающего молекулярную палеонтологию с палеонтологией морфологической.
Известно, что все живущие на Земле организмы вне зависимости от систематической принадлежности обладают определенным единством биохимических процессов и большим сходством состава и структуры всех основных биомолекул. Вместе с тем они отличаются друг от друга по относительному количеству синтезируемых веществ, а также наличию специфических соединений для каждого данного таксона - вида, рода, семейства и т. п. Таксономическая специфичность проявляется наиболее отчетливо в составе основных биополимеров, в частности в характерных соотношениях и последовательности составляющих их мономеров [501]. Это и обусловливает возможность осуществления хемотаксономических исследований.
При изучении биомолекул, извлеченных из молодых пород, можно непосредственно исходить из данных сравнительно-биохимического анализа современных растительных и животных организмов и, основываясь на их хемотаксономических признаках, судить о присутствии или, наоборот, отсутствии близкородственных им существ в среде накопления этих пород. С принципиальной точки зрения аналогичный подход возможен и к изучению хемофоссилий, содержащихся в древних отложениях, хотя его осуществление сильно осложнено вторичными изменениями ископаемых биомолекул и общим ходом биохимической эволюции. Рассматриваемому направлению посвящено уже немало работ, среди которых нужно выделить труд под редакцией М. Флоркэна и X. Масона [1445].
Накопившийся к настоящему времени фактический материал позволяет составить общее представление о распределении органических соединений в осадочной оболочке Земли [501, 1243]. Нельзя, однако, думать, что основным определяющим фактором является абсолютный возраст пород, вмещающих эти вещества. Уменьшение количества растворимых органических геолипидов при одновременном возрастании роли углеводородов и графитизации органических остатков обусловлено преимущественно медленными термическими и каталитическими процессами, протекающими в осадках после захоронения, уплотнения и литификации; абсолютный же возраст пород имеет, по-видимому, существенно меньшее значение, чем термическая история (рис. 88).
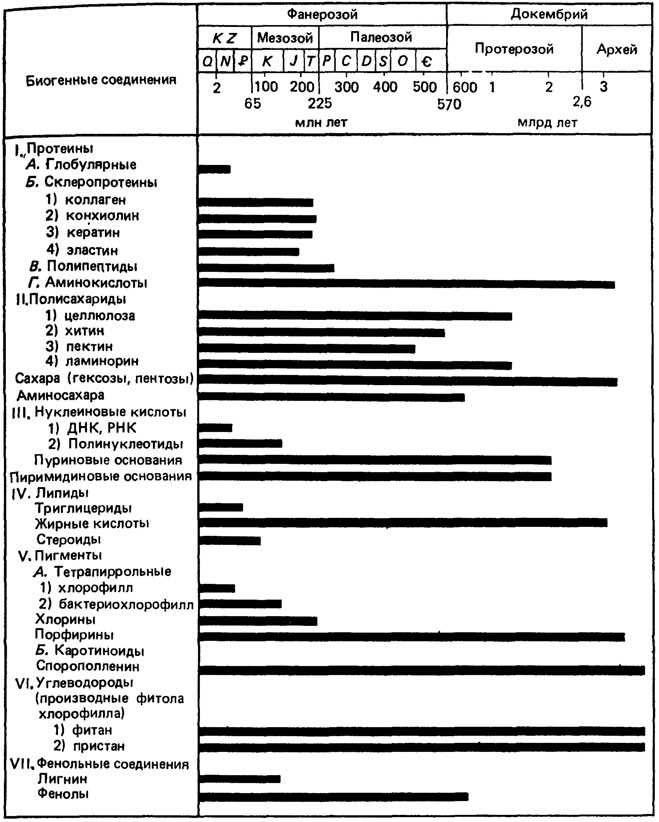
Рис. 88. Органические соединения, идентифицированные в ископаемых остатках растительных и животных организмов. По Ч. М. Колесникову
Несмотря на то что рассматриваемой проблеме посвящена значительная литература, современное положение дел нередко приводит к неоднозначному толкованию одних и тех же фактов. Весьма сложным оказывается доказательство первичной биогенной природы многих углеродистых соединений, заключенных в древних отложениях, содержащих также морфологические остатки организмов. Известно, что аминокислоты и некоторые другие соединения могут образовываться и неорганическим путем, в том числе в процессе вулканических извержений [625]. Обнаружены они и в углеродистом веществе метеоритов [1384, 1858 и др.], иногда очень древних.
Обладая высокой подвижностью, углеродистые соединения различного (биогенного и абиогенного) происхождения испытывали в течение длительного геологического времени миграцию и преобразования. Поэтому наиболее надежными являются результаты изучения нерастворимой фракции органических веществ горных пород - керогена, который менее мобилен и с большей вероятностью, чем другие хемофоссилии, сингенетичен вмещающим их образованиям.
Таким образом, изучение хемофоссилии с целью получения информации об их происхождении и истории, а в конечном итоге выяснения закономерностей биохимической эволюции представляет большие сложности. Один из путей их преодоления - дальнейшее сближение молекулярной палеонтологии с палеонтологией морфологической и экологической, без участия которых невозможно решение палеохемотаксономических и эволюционных вопросов, а также выяснение состава и закономерностей формирования материнского вещества горючих полезных ископаемых. Следует обратить внимание также на существование еще более широких возможностей, состоящих в развитии, наряду с молекулярной палеонтологией, других наук, теснейшим образом связанных с морфологической палеонтологией, - палеобиохимии, палеобиогеохимии и палеогистологии [501]. Систематическое укрепление связей между этими науками расширяет и углубляет роль палеонтологии в изучении происхождения и развития органического мира и эволюции биосферы в целом.
В заключение можно отметить, что молекулярной палеонтологией до сих пор не найдены доказательства добиологической эволюции. Исследования последних лет все стремительней отодвигают временною границу появления жизни на Земле. Непрерывность наличия органического вещества биогенного генезиса на протяжении всего времени формирования осадочных пород и существование фотосинтеза как биологического процесса на протяжении почти 4 млрд. лет подтвердились комплексными исследованиями ранней органической эволюции в рамках Международного проекта геологической корреляции (МПГК) № 157 "Ранняя органическая эволюция, минеральные и энергетические ресурсы", выполнявшегося с 1977 г. под руководством проф. М. Шидловски (ФРГ).
Согласно Г. Пфлюгу (Pflug H. D., 1984 г.), породы с возрастом 4 млрд лет содержат структурированное органическое вещество, а приемлемые условия жизни на Земле появились 4,2-4,3 млрд лет назад. Наконец, как полагает Г. В. Войткевич (1986 г.), цианобактерии возникли задолго до временной отметки 4 млрд лет, первичные гетеротрофы получили развитие 4-4,55 млрд лет назад, сама же длительность жизни на Земле соизмерима с возрастом планеты или чрезвычайно близка к нему.
Участие живого вещества в геологических процессах во все времена истории Земли свидетельствует о биогенном происхождении всего органического вещества стратисферы, как полагал В. И. Вернадский. В связи с этим следует еще раз подчеркнуть роль молекулярной палеонтологии как науки о палеонтологическом аспекте исследований всего ископаемого органического вещества стратисферы.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'