
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Методы исследований в палеоэкологии
Среди методов палеоэкологии выделяются следующие: морфофункциональный, актуалистический, тафономический, количественные, экспериментальный и биогеохимические. Эффективность исследований связана с комплексным использованием методов, однако значимость каждого из них определяется конкретными условиями работы и зависит от ряда причин - степени близости вымершей группы к ныне живущей, древности отложений, сохранности окаменелостей и т. д.
Морфофункциональный метод. Это один из наиболее ранних методов палеоэкологии, позволяющий на основании изучения формы окаменелости или следа жизнедеятельности судить об образе жизни и отчасти условиях обитания организма-хозяина. Анализ базируется на представлении об адаптивном значении большинства структур или раковины. Например, сильно выпуклая левая створка Fatina рассматривается как приспособление к лежанию на ней взрослой особи, а очень толстая раковина - результат обитания в теплых мелких водах палеогенового моря [235, 391]. При этом нужно помнить, что, с одной стороны, не всякая адаптация реализуется в морфологии, а с другой - морфологическая лабильность ограничена эволюционными возможностями той или иной группы организмов. Так, мелкораковинные двустворчатые моллюски обычно менее специализированы и редко имеют какие-либо особые приспособления. Многие структуры не могли развиваться из-за несовместимости с первичными структурами. В связи с жесткими морфологическими ограничениями многие признаки повторялись, что обусловило конвергенцию [2316].
Классическим примером, до сих пор не утратившим своего значения, является эволюционный анализ В. О. Ковалевского формы скелета третичных лошадиных в связи с изменением функции органов передвижения и захвата пищи, вызванным сменой условий их обитания [494]. Исследования беспозвоночных (тетракораллов, брахиопод и морских лилий) в этом направлении были проведены Н. Н. Яковлевым [1239].
Результаты грамотно выполненного морфофункционального анализа считаются в палеоэкологии самыми надежными для суждения об образе жизни организмов. Однако получаемые выводы могут быть неравнозначны. Наиболее обоснованными следует считать суждения об образе жизни вымерших организмов, потомки которых сохранились до настоящего времени. Так обстоит дело с крупной ветвью детритоядных двустворчатых моллюсков - ктенодонтов (палеотаксодонтов). Сходство морфологических структур синуса мантийной линии, рострообразно удлиненный задний край, а также наличие внутренних и наружных лигаментных образований позволяют не только предположить похожий образ жизни этих моллюсков в течение всего фанерозоя, но и высказать достаточно убедительное мнение об условиях их обитания исходя из одинаковых приспособлений для добывания пищи [678, 911]. Уникальные находки пиритизированных ядер кишечника нукулан (ктенодонты) из нижнеюрских отложений Англии, показавшие сходство в анатомическом строении, а следовательно, способе питания ископаемых и современных нукулан, еще раз подчеркивают высокую эффективность морфофункционального анализа [1458].
Критерием достоверности реконструкции образа жизни полностью вымерших групп могут быть тафономические наблюдения, нахождения их скелетных остатков в прижизненных захоронениях. Например, находки раковин нескольких экземпляров раннепермской Meeckella, соединенных в один пучок (макушками) и несших следы совместного роста, убедили Н. Н. Яковлева [1236, 1238, 1239], что эти брахиоподы прирастали при жизни к твердому морскому дну.
Менее надежны, но все-таки заслуживают внимания сходные реконструкции, выполненные другими исследователями. Так, В. И. Бодылевский [95] и В. Швайцер [2251] независимо друг от друга пришли к единому мнению об образе жизни раннеюрского двустворчатого моллюска Plicatula (Harpax) spinosa, лежавшего на взрослой стадии на правой створке, покрытой тонкими иглами. Б. В. Наливкин [725] и Д. Никол [2008] на основе функциональной морфологии раковины Conocardium сделали вывод об эпибентосном образе жизни этого рода (рис. 89).
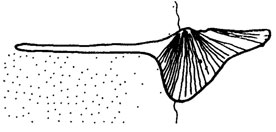
Рис. 89. Предполагаемое положение раковины Conocardium на грунте: По Б. В. Наливкину. Реконструкция дана на основании морфофункционального анализа
Хорошо известно, что сходный образ жизни нередко приводит к конвергенциям. Конвергенции обнаружены среди ряда групп ископаемых. Особенно часты они у беспозвоночных, что, возможно, связано с морфологическими ограничениями: использование в качестве адаптивных сравнительно небольшого числа морфологических структур приводило к повторениям признаков. Таковы хрестоматийные ряды кубковых у жесткоприрастающих - губок, археоциат, ругоз, усоногих раков, двустворчатых моллюсков, брахиопод [342]; примером могут служить сосуды свободнолежащих морских двустворок (грифеид, пектинид) и брахиопод (продуктид).
При выяснении функции той или иной структуры не следует забывать о мультифункциональном значении некоторых из них. Необходимо обратить внимание на корреляцию признаков. Например, наличие глубокого синуса у двустворок коррелятивно связано с удлинением задней части раковины.
Одна из ярких особенностей экзоскелета беспозвоночных (моллюсков, брахиопод и др.) - линии роста на раковинах, свидетельствующие о неравномерности (периодичности) роста животного. Периодичность роста бывает связана с приливно-отливными, лунными, месячными или сезонными циклами. Расшифровка этих событий по характеру знаков роста также входит в круг вопросов, решаемых функциональной морфологией. В этой же связи нередко изучается микро- и ультраструктура раковин.
Наиболее сложным случаем морфофункционального анализа является восстановление функции животного по форме оставленного им следа. Известно, что в одном случае разные животные оставляют весьма сходные следы, а в другом случае одно и то же животное - разные следы в зависимости от поведения. Критерием, повышающим достоверность заключения о природе следа, могут быть находки в тех же слоях остатков животных-хозяев, а также аналогов-построек, сделанных ныне существующими животными. Именно таким способом была доказана связь нор в палеогеновых отложениях Средней Азии с жизнедеятельностью десятиногих раков (Calianassa) на прибрежной песчаной литорали Ферганского залива палеогенового моря [235]. По отпечаткам конечностей трилобитов (Phacops, Homalonotus?, Illaenus?, Redlichia) были сделаны заключения об их форме и функциях [2265].
Актуалистический метод. В основу этого метода положен широко распространенный в геологии принцип униформизма, основанный на единстве и постоянстве законов природы. Применительно к палеоэкологии актуалистический подход состоит в использовании сведений об образе жизни и условиях обитания ныне живущих организмов разного таксономического ранга и биоценозов для суждения об образе жизни и условиях обитания родственных организмов и палеоценозов, существовавших в геологическом прошлом. При таком подходе лишь в редких случаях полностью отождествляются характеристики современных и вымерших организмов. Существующие ограничения сводятся к следующему: 1) чем древнее организм, тем менее определенно суждение об его экологии по аналогии с родственным рецентным организмом из-за экологической лабильности биоты; 2) в течение геологического времени постоянно изменялась и биотическая, и абиотическая составляющие среды; 3) в ряде случаев не хватает информации по экологии современных организмов, в особенности морских беспозвоночных.
Для иллюстрации первого ограничения рассмотрим возможности использования актуалистического метода в пределах биологических единиц разного таксономического ранга. Согласно представлениям современной биологии, каждый вид экологически специфичен [1156]. Таким образом, можно предполагать сходную экологию вида в течение времени его существования. Известно, что темпы эволюции различны не только у крупных групп организмов, но и в пределах таксонов сравнительно невысокого ранга, например семейств. Поэтому продолжительность существования видов у разных групп организмов и внутри одной группы неодинакова. Вероятно, большинство ныне живущих видов появилось в плейстоцене, некоторые уходят корнями в плиоцен. Для всех этих видов предполагается устойчивая экологическая толерантность во времени [1259].
Род - более консервативная единица эволюции. По сравнению с видами роды, как правило, более долговечны. Достаточно сказать, что в современных морях живет немало родов беспозвоночных, известных с палеогена и даже с раннего мезозоя. Экологический диапазон рода слишком широк, чтобы успешно интерпретировать эту особенность в отдаленное геологическое прошлое. Однако большинство родов в течение времени своего существования устойчиво сохраняют присущий им образ жизни - поведение и способ питания. Нередко тип пищи и способ питания характеризуют и более высокие таксоны беспозвоночных. Так, все замковые брахиоподы относятся к сессильным фильтраторам высокого уровня и т. п. Способ же питания часто непосредственно связан с определенными абиотическими характеристиками среды. Например, собиратели-детритофаги, питающиеся с уровня осадок - вода, могут существовать только в спокойных (глубоких? низкотемпературных? с аномальным газовым режимом?) водах, обеспечивающих достаточную концентрацию пищи на этом уровне.
Для подкрепления второго ограничения обратимся к истории биосферы. Специалисты почти единодушны в том, что первичная атмосфера Земли (период преджизни) была бескислородной. После достижения точки Пастера (1% кислорода от современного насыщения) и реализации органического фотосинтеза (около 1,5 млрд лет назад) количество свободного кислорода в атмосфере неуклонно возрастало и резко увеличилось с момента выхода растений на сушу, что стало возможным, когда насыщение палеоатмосферы кислородом достигло 10% современной величины.
Вероятно, лишь в середине палеозоя было достигнуто современное насыщение кислородом, однако с тех пор и до настоящего времени постоянно происходили слабые колебания его содержания в атмосфере. В соответствии с изменением содержания кислорода менялось и содержание диоксида углерода - оба эти компонента атмосферы весьма существенно влияли на ход развития биосферы Земли*. Изменялась в таксономическом и в структурном плане и сама биота. Разнообразие типов адаптации в органическом мире должно было возрастать по мере дифференциации обстановок обитания и усложнения взаимоотношений между организмами (конкуренция, хищничество, комменсализм, мутуализм, паразитизм и др.). Следовательно, применение метода актуализма в палеоэкологии при исследовании дораннепалеозойской биоты должно быть сильно ограничено.
* (Превращения биотической и абиотической составляющих среды тесно взаимосвязаны. Крупные эволюционные новации в биоте нередко коренным образом изменяли физико-химические характеристики среды. Так, практически весь свободный кислород в атмосфере имеет биогенное происхождение. Содержание кислорода, освобождаемого при неорганической фотодиссоциации воды, не могло подняться выше так называемого уровня Юри, т. е. менее 0,001% его содержания в современной атмосфере [895].)
Третье ограничение, касающееся недостатка знаний об экологии современных, в частности морских беспозвоночных, будет со временем снято. Пока же данных по экологии рецентных видов и сообществ недостаточно для более эффективного использования в палеоэкологии метода актуализма. В гидробиологии в настоящее время интенсивно накапливаются сведения о расселении видов в зависимости от физико-химических факторов среды: температурных, эдафических, батиметрических, солевых, газовых, величины рН и других; много внимания уделяется анализу разнообразия и структуры сообществ. Однако еще мало работ по тафономии беспозвоночных, в которых рассматривается посмертная судьба разных групп бентоса и планктона, географические ареалы ископаемых остатков на разных стадиях захоронения популяций, механизм их транспортировки, результаты воздействия организмов на различные типы субстратов и др. Наконец, водные биоты изучены хуже, чем наземные, относительно этологии беспозвоночных и биотических взаимоотношений (комменсализм, мутуализм, паразитизм и др.).
Не все эти вопросы одинаково волнуют экологов и палеоэкологов. Некоторые особенности экологии и тафономии сообществ интересны только палеоэкологам. Неслучайно именно палеонтологи первыми начали изучать современные прибрежные морские сообщества для познания палеоэкосистем. Данное направление получило название актуопалеонтологии [2157]. Начатые Р. Рихтером более полувека назад актуопалеонтологические и актуогеологические исследования приливно-отливных зон (ваттов) Северного моря дали богатый материал для расшифровки системы "организм - среда" в геологическом прошлом [2140, 2212]. Число публикаций, посвященных биоценологии и тафономии гидробионтов, неуклонно растет, причем авторами многих из них являются палеонтологи. Это свидетельствует не только о возрастающем интересе палеонтологов к современным водным экосистемам, но и об их потребности в расширении сведений по экологии и тафономии данных сообществ для анализа палеоэкосистем.
Важная сторона актуопалеонтологических исследований - наблюдения над следами жизнедеятельности организмов, которым гидробиологи почти не уделяют внимания [1606]. Однако ни результаты современных экологических процессов, ни условия, в которых они протекают, не могут быть непосредственно использованы для истолкования палеоэкосистем. Применение метода актуализма в палеоэкологии основано на признании единых законов и правил, управляющих развитием биот в течение всего времени их существования. Действие экологических правил определяется фундаментальными законами биологической эволюции: изменчивостью, наследственностью и естественным отбором. Так, любой вид, как продукт эволюции, всегда занимал строго определенное место в биосфере (действие экологического закона лимитирующих факторов). Всякая экосистема в геологическом прошлом, как и теперь, характеризовалась структурностью, энергетикой, биотическим круговоротом, давлением отбора и гомеостазом. Одним из постоянно действующих биотических факторов отбора следует признать конкурентное взаимоисключение из сообществ экологически сродственных видов (правило Гаузе). Общие с современными законы управляли организацией живых существ на популяционном уровне на всех этапах существования биоты Земли.
Тафономический метод. Сбор тафономической информации для целей палеоэкологии нельзя отнести в строгом смысле к чисто палеоэкологическим методам. Особенности тафономии лишь отчасти обусловлены биологической природой объектов. В значительной степени они связаны с физико-химическими сторонами процессов седиментации, диагенеза и последующих вторичных преобразований. Специфика тафономических наблюдений состоит в том, что они полностью проводятся в полевых условиях. Хотя изучение захоронений фоссилий в геологических разрезах является, по существу, пропалеоэкологическим, оно чрезвычайно важно, так как позволяет получить более половины общего объема данных, привлекаемых для последующего палеоэкологического анализа.
Тафономические наблюдения являются многоцелевыми. Во-первых, они нередко дают возможность судить об этологии животных. Находки захороненных в прижизненном положении экземпляров отдельных видов описаны почти для всех типов беспозвоночных: простейших [424, 1254, 1308, 2112 и др.]; археоциат [371]; кишечнополостных [223, 1860]; мшанок [33, 392 и др.]; брахиопод [223, 418, 602, 1239, 2422]; членистоногих [614, 2158]; иглокожих [223, 226, 1860]; двустворчатых моллюсков [392 и мн. др.]. Во-вторых, тафономические данные позволяют довольно точно оценить одну из сторон процесса древнего осадконакопления на конкретном участке палеобассейна - установить его динамический режим. Преобладание автохтонно захороненных остатков организмов свидетельствует о стабильности процесса осадконакопления, а наличие прижизненных захоронений и о высоких темпах накопления осадков. Аллохтонные захоронения, как правило, связаны с изменчивым режимом. Ракушниковые скопления - гнездовидные, линзовидные, пластообразные, типа "ракушниковая мостовая" и типа "роза" - могут указывать на отрицательный баланс седиментации.
И все же первейшей задачей тафономических наблюдений в палеоэкологии является установление типа ценоза, позволяющего судить о прижизненных ассоциациях видов в палеосообществах. Палеоэколог правомочен использовать концепцию современного биотического сообщества по отношению к комплексу видов, встреченному в элементарном ориктоценозе (слое или части слоя), только доказав, что этот комплекс представляет собой прижизненную видовую ассоциацию. Такое Доказательство основано главным образом на тафономических наблюдениях, сопровождающихся предварительными седиментологическим и палеоэкологическим полевыми анализами. Тип ценоза оценивается для популяции каждого вида, обнаруженного в элементарном ориктоценозе. Тафономические исследования предусматривают определение типа захоронения, ориентировки окаменелостей, степени сохранности и сортировки, а также посттафономических преобразований [226, 394, 971, 2003]. Дополнительные данные о наличии или отсутствии послесмертной транспортировки могут быть получены в результате анализа размерно-частотных кривых, подобных показанной на рис. 90 [1360, 1460, 1474, 1570, 2283, 2372, 2396 и др.].
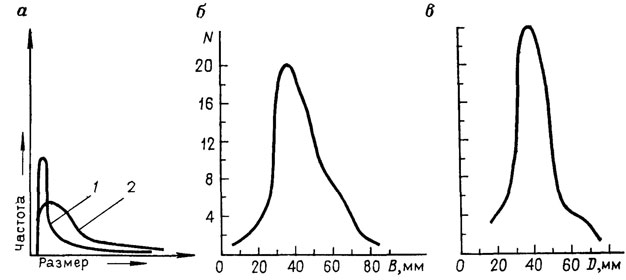
Рис. 90. Размерно-частотные кривые распределения экземпляров в ископаемых популяциях. По В. А. Захарову. а - теоретические (1 - молодые экземпляры, 2 - взрослые, из Boucot); б-в - природные захоронения бухий в берриасе на севере Сибири, р. Боярка. N - количество особей; В - расстояние между плоскостью смыкания створок; D - расстояние между наиболее удаленными точками переднего и заднего края
При тафономических наблюдениях нельзя упускать из виду возможность полного растворения скелетов на всех стадиях захоронения: от момента погружения в осадок (например, растворение [70] раковинок планктонных фораминифер на поверхности осадка ниже "критических" глубин - 3500-4000 м в высоких широтах и 4500-5000 м в низких) до выхода в обнажениях на современную дневную поверхность, где они могут подвергаться выщелачиванию агрессивными грунтовыми водами и т. п. Здесь следует подчеркнуть, что наблюдаемые в ориктоценозе биотические ассоциации всегда будут лишь частью сложной системы древнего биоценоза.
Количественные методы. Привлечение количественных методов в палеоэкологии необходимо для оценки таких важных биотических характеристик сообществ, как разнообразие и популяционная плотность составляющих видов. Уже отмечалось, что использование концепции современного биотического сообщества в палеоэкологии тормозится в значительной степени из-за отсутствия общепринятых методов количественного учета окаменелостей. Существующие же приемы часто субъективны и весьма трудоемки.
Широко распространенные в палеоэкологии приемы количественного учета основаны на словесной оценке встречаемости окаменелостей (фон, обилие, часто, редко, единично и др.). Они не могут применяться для анализа и сравнения сообществ, их можно лишь с ограничениями использовать для характеристики динамики ископаемых популяций.
Количественная оценка осуществляется прямым подсчетом окаменелостей на определенной площадке поверхности напластования или в фиксированном объеме породы. Число опробований конкретного ориктоценоза устанавливается опытным путем. Минимальный предел считается достигнутым, если дальнейшее увеличение числа проб (площади квадрата, объема пробы) не дает эффективной прибавки информации (рис. 91). Полученные средние показатели обычно распространяются на весь элементарный ориктоценоз.
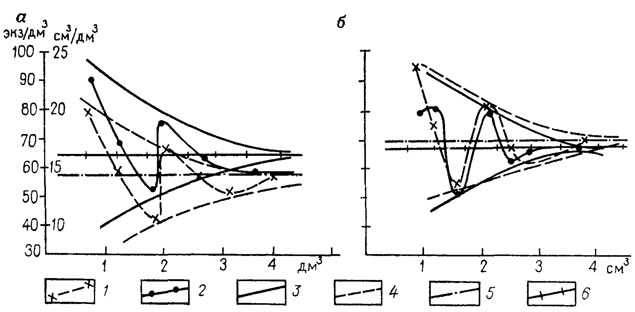
Рис. 91. Определение минимально допустимого объема пробы по графику. По Ю. С. Салину. На оси абсцисс показан объем пробы в 1 дм3; на оси ординат - содержание окаменелостей в экземплярах в 1 дм3; пробы или (справа) в 1 см3; биообъема на 1 дм3; пробы. При объемах пробы от 1 до 3 дм3; содержание окаменелостей резко меняется; с увеличением объема пробы колебания уменьшаются и при значении 3-4 дм3; нет существенных отклонений содержаний окаменелостей по разным пробам; а - проба № 1, б - проба № 2; 1 - содержание см3/дм3, 2 - содержание, экз/дм3; 3 - линии, ограничивающие поле колебаний содержания см3/дм3; сокращающегося с увеличением объема пробы; 4 - то же, в отношении содержания экз/дм3; пробы; 5, 6 - среднее содержание по этим же показателям
Для опробования макроокаменелостей площадным способом рекомендуется метод квадратов. Площадь квадрата устанавливается экспериментально исходя из условия работы и характера распределения окаменелостей. При исследовании морской мезозойской и кайнозойской шельфовой бентосной фауны наиболее эффективен квадрат со стороной 1 м [1259]. Наиболее существенным ограничением надежности данного метода являются следующие допущения: 1) встреченные на плоскости напластования виды жили совместно, 2) одновременно жили только те виды, которые встречены. Известно, что некоторые эндобионты в прошлом, как и ныне (например, аномалодесматы, двустворчатые моллюски, лингулиды, многие иглокожие и др.) могли зарываться глубоко в грунт и захороняться в прижизненном положении ниже уровня одновременно обитавшего эпибиоса. С другой стороны, многие эпиобионты, в особенности на биогенных, например рифовых постройках, после смерти падают на дно и их погребение осуществляется на единой поверхности напластования совместно с организмами другой природы. Рассматриваемый метод совершенно непригоден при изучении ракушниковых скоплений.
Метод прямоугольника рекомендуется использовать при изучении вертикальных стенок в пределах однородного слоя. Для этого же случая предлагается и метод "протяженной линии". Суть последнего в следующем: параллельно поверхности напластования проводится линия, затем подсчитывается каждый, пересекаемый ею объект. Недостаток этого метода в том, что виды с крупной раковиной будут чаще пересекаться линией, чем виды с мелкой. Описанный метод малоэффективен при изучении ориктоценозов со слабой насыщенностью окаменелостями [1259].
Объемные методы опробования оправдывают себя при работе с микрофоссилиями. Количественная оценка встречаемости вида на 100 г породы позволяет, например, судить о структуре комплекса в ориктоценозе и проводить сравнения с другими ориктоценозами. Однако при отсутствии сведений об условиях формирования комплекса использование приемов обработки материалов с позиции биотического сообщества невозможно.
Объемные методы количественной оценки содержания макро-окаменелостей допустимы потому, что перед опробованием ориктоценоза на основании тафономических и других полевых наблюдений можно определить тип (автохтонный или аллохтонный) ископаемого танатоценоза. Однако этот метод весьма трудоемок. Существует несколько модификаций объемного метода. Одна из них предложена Ю. С. Салиным [905]. Мощность опробуемого интервала 5-15 см. Экспериментально установленный минимально допустимый объем пробы 3-4 дм3, но может достигать 6-10 и даже 30 дм3 (см. рис. 91). Такие объемы соответствуют объемам дночерпальных проб гидробиологов. Объем породы в полевых условиях устанавливается исходя из объема воды, вытесненной пробой из фиксированного объема воды в сосуде. Окаменелости извлекаются в лаборатории, где определяется и их объем. В каждой пробе подсчитывается количество экземпляров каждого вида и рода, общее число экземпляров, процент целых и разрозненных створок. Выясняют также таксономический состав каждой пробы. Количественные соотношения различных группировок беспозвоночных определяются не по числу экземпляров какой-либо жизненной формы, а по ее биообъему, который в случае полной сохранности экземпляра может быть установлен с помощью изложенного способа или по специальной, разработанной Ю. С. Салиным методике [905]. Затем в каждой пробе вычисляют: 1) общее содержание окаменелостей в 1 см3 биообъема на 1 дм3 породы или в процентах; 2) соотношения (по объему) группировок (трофических, эдафических, термальных, гидродинамических и др.); 3) индексы преобладания группировок и объемных форм; 4) количественную структуру сообщества.
Описываемый метод лишен недостатков, присущих глазомерным оценкам. Он ставит на объективную основу палеоэкологические характеристики и позволяет проводить сравнительный анализ сообществ в пространстве и времени. Однако точность данного метода прямо пропорциональна объему полевых и лабораторных работ, что делает его чрезвычайно трудоемким и неоперативным. Имеются также отдельные технические ограничения, в частности, использование объемного метода невозможно при изучении рыхлых и мерзлых пород.
В последнее время получила распространение полуколичественная оценка. Суть ее - в приближенной числовой характеристике частоты встречаемости таксона в ориктоценозе или образце (пробе), выраженной в баллах. Предложены два варианта оценки. Для микрофауны разработана шестибалльная шкала: 0 - отсутствие представителей таксона в образце; 1 - единичные экземпляры; 2 - от 10 до 30; 3 - от 30 до 50; 4 - от 50 до 100; 5 - больше 100. Выбор интервалов, соответствующих баллам, как и число баллов, может быть различным [870]. Другой вариант рекомендован для макрофауны [783]. Здесь предлагается семибалльная шкала, основанная на частоте встречаемости какого-либо вида в определенном интервале разреза (едином ориктоценозе). При этом использованы следующие категории: изобилие (сотни экземпляров - 100 баллов) очень много (многие десятки - 30); много (первые десятки - 9); очень часто (11-15 экземпляров - 5); часто (6-10 экземпляров - 3); редко (3-5 экземпляров - 2); очень редко (1-2 экземпляра - 1). Баллы - 100, 30, 9, 5, 3, 2, 1 - показывают условную долю каждой категории в ориктоценозе, поэтому они могут быть названы показателями частоты встречаемости.
Полуколичественная оценка рассчитана на 1,5-2 ч непрерывных наблюдений. Практика повторных наблюдений на разрезах показала, что расхождения в оценке лежат в пределах соседних категорий и касаются лишь редко встречающихся таксонов. Резко дифференцированная шкала баллов, предложенная во втором варианте, позволяет без дополнительной обработки данных с помощью простых графиков и циркограмм описать и оценить такие важные характеристики сообществ, как доминирование (с учетом видового разнообразия), выровненность и трофическая структура (рис. 92).
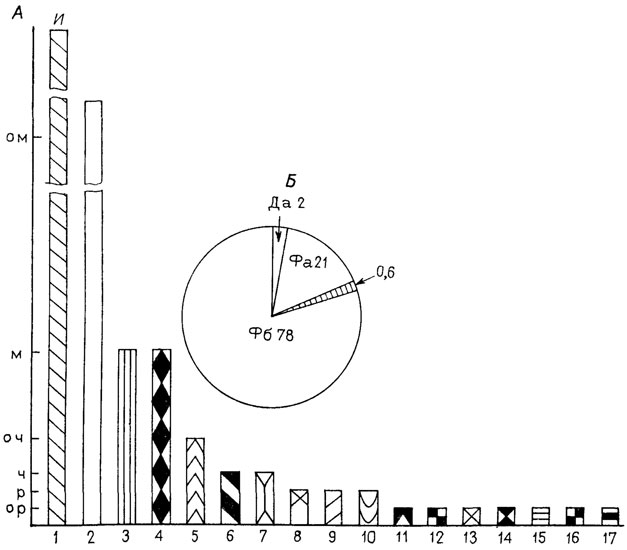
Рис. 92. Структура климаксного мелководного устричного сообщества. Рекомендация по ориктоценозу слоя XXXI опорного разреза неокома, нижний валанжин. Север Сибири, р. Боярка По В. А. Захарову. А - таксономическая структура. По оси абсцисс - показатель частоты встречаемости, о. р - очень редко, р - редко; ч - часто; о. ч - очень часто; м - много; о. м - очень много; и - изобилие. По оси ординат - рода беспозвоночных. 1 - Ostrea, 2 - Arctica, 3 - Musculus, 4 - Modiolus, 5 - Boreionectes, 6 - Chlamys, 7 - Artichnus, 8 - Cuculea, 9 - Astarte, 10 - илоеды-черви, 11 - Aguilerella, 12 - Pinna, 13 - Buchia, 14 - Oxytoma, 15 - Pteuromya, 16 - Quenstedtia, 17 - Pseudolimea и Limea. Б - трофическая структура (циклограмма): Да - детритофаги низкого уровня (под грунтом); Фа - фильтраторы низкого уровня; Фб - фильтраторы высокого уровня; цифры показывают процентное содержание трофических групп. Заштрихованная площадь - доля Buchia среди фильтраторов высокого уровня
Полуколичественная оценка окаменелостей в ориктоценозе достаточно оперативна и нетрудоемка. Использование этого приема не исключает выборочного количественного контроля, позволяющего повысить достоверность оценки посредством введения поправочного коэффициента для конкретных условий.
При использовании количественных методов в палеоэкологии можно воспользоваться некоторыми формулами, предложенными экологами для характеристики видовой структуры сообществ [767].
Экспериментальный метод. В морской экологии проводятся эксперименты по выяснению влияния на состав и структуру сообществ таких важных факторов среды, как соленость и температура, путем изменения их параметров в аквариумах, заселенных подопытным сообществом [1343]. Другие эксперименты направлены на выявление особенностей формирования сообществ в связи с оккупацией "новых" биотопов, например искусственно созданных прудов или очищенных на мелководье площадок [1701, 1904]. Закономерную смену сообществ во времени (сукцессии) иногда удается наблюдать в открытых морских экосистемах. Например, описаны сукцессии бентосных сообществ на прибрежно-морском мелководье у берегов Англии после суровой зимы 1962-1963 гг. [1259]. Результаты таких экспериментов используются при культивировании морских экосистем. Они представляют интерес и при интерпретации палеоэкосистем, прямые эксперименты с которыми исключены.
При этом следует помнить, что экспериментальные ситуации, хотя и реальные, но все же детерминированные: в открытых морских экосистемах связи сложнее и последствия обычно непредсказуемы. Многочисленные попытки моделирования морских экосистем пока не дали существенных результатов. То же касается и палеоэкосистем.
Более обнадеживают эксперименты с окаменелостями как физическими телами с целью выяснения особенностей посмертной транспортировки и ориентировки скелетов во время захоронения, а также интерпретации функциональной морфологии. Для этих экспериментов используют хорошо сохранившиеся раковины и другие скелеты современных беспозвоночных, а также имитирующие их форму пластиковые и другие модели. Эксперименты по транспортировке и ориентировке проводятся в гидродинамических лотках с меняющимся режимом потока или в природных водоемах, обычно в их прибрежной полосе. Объектом экспериментов служат чаще всего раковины моллюсков [1292, 1320, 1371, 1432, 1615], реже фораминифер [1844], остракод [1843] и другие. Функциональное значение некоторых особенностей строения раковин вымерших организмов выясняется с помощью пластиковых или плексигласовых моделей.
Так, на пластиковых моделях раковин цефалопод цератитового и дактилиоцератитового типов была установлена прямая зависимость плавучести от степени эволютности раковины [1996]. Опыты с разнообразными плексигласовыми моделями показали, что форма раковины, характер ее вентральной стороны и размеры умбиликуса существенно влияли на коэффициент торможения (и, следовательно, плавучесть) раковин цефалопод [1421, см. 1.3]. Эксперимент с механическим зарывающимся роботом, имеющим форму раковины двустворчатого моллюска, был поставлен для проверки высокого адаптивного значения прозогирной формы раковины и ротационного механизма зарывания этих организмов [2317].
Нередко экспериментальные исследования комбинируются с наблюдениями непосредственно на разрезах. Совместный анализ этих данных позволяет получить наиболее достоверные результаты [1891].
Биогеохимические методы. В современной биогеохимии имеются методы, которые позволяют дать количественную оценку таким важным абиотическим факторам палеобассейнов, как температура и соленость.
Палеотермометрия. Для определения палеотемператур используются следующие отношения: 18О/16О; Ca/Mg, Sr/Ca и арагонит/кальцит.
Наиболее надежные результаты получены по изотопам кислорода. Этот метод основан на установленной зависимости равновесного распределения тяжелого изотопа кислорода (18О) между кислородом воды и карбоната, образующегося в этой воде [1045]. Отношение 18О/16О определяется в СО2, извлеченном из СаСО3 раковин беспозвоночных. Объектами исследований являются, как правило, раковины моллюсков (например, в юрской и меловой системах), ростры белемнитов. Значение палеотемпературы устанавливается обычно с точностью до 1°С, иногда до 0,5°С [115]. Теоретически изотопный метод позволяет определить температуры образования карбонатов с возрастом до 700 млн. лет. Однако из-за сильных изменений древних пород в результате диагенеза достоверность полученных значений палеотемператур для палеозоя невелика. Кроме того, использование изотопного метода предполагает одинаковое (или близкое) содержание изотопов кислорода в водах современного и древнего океанов, что маловероятно. Впрочем, имеющиеся данные изотопной палеотермометрии для мезозоя и кайнозоя, как правило, не противоречат представлениям о температурах морских вод и климатах, полученным с помощью палеобиологических и геологических методов.
Отношения Ca/Mg и Sr/Ca для целей палеотермометрии теоретически малообоснованны и менее надежны, чем метод изотопов кислорода, так как распределение магния и стронция в раковинах беспозвоночных зависит от минеральной формы карбоната (кальцит или арагонит), филогенетического уровня организмов и их физиологии, солености бассейна и других факторов среды [1045]. Тем не менее Ca/Mg-отношения используются в палеотермометрии юрского и более молодых периодов. Большое преимущество этого метода по сравнению с изотопно-кислородным - оперативность и дешевизна, однако погрешности данного метода выше и в несколько раз превосходят ошибки изотопного. Минимальные погрешности в определении палеотемператур Ca/Mg методом составляют 1-2°С [79, 80].
Пока не удалось выявить основной фактор, влияющий на распределение стронция в органогенных карбонатах [1045]. Известно, что этот элемент концентрируется предпочтительнее в арагонитовых, а не в кальцитовых раковинах [115]. У исследователей нет единого мнения в вопросе о том, температура или соленость оказывает доминирующее воздействие на соотношение стронция и кальция в скелетах морских беспозвоночных. Эти расхождения в ряде случаев объясняют таксономическим и физиологическим контролем [1045]. Для некоторых моллюсков, в частности мидий, установлена положительная связь между отношением арагонита и кальцита в составе раковин с температурой воды, в которой они живут. Однако это отношение меняется в процессе роста раковин. На него влияют также некоторые особенности биогенного синтеза СаСО3: определенный набор составляющих органической матрицы стимулирует образование то арагонита, то кальцита [1045]. Таким образом, в настоящее время не может быть указан какой-либо конкретный фактор, влияющий на химический и минеральный состав карбонатных скелетов беспозвоночных. Но экспериментально выявлены некоторые группы организмов, например отдельные моллюски, раковины которых могут быть использованы для суждения о палеотемпературах и палеосолености (см. далее) среды обитания.
Палеогалометрия. Биогеохимические методики определения солености основаны на устойчивых корреляциях между содержанием химических элементов в воде и в карбонатных скелетах беспозвоночных. Один из таких методов разработан на раковинах устриц. При исследовании раковины устрицы Crassostrea virginica (атлантическое побережье Северной Америки) в водах с известной соленостью (13-30,5‰) было установлено, что содержание Mg + Sr и Mn обратно пропорционально солености, а содержание Na с возрастанием солености, в общем, увеличивается [2188].
Палеосоленость может определяться двумя способами: по графику линейной регрессии (рис. 93) и по формулам статистики [399]. Обобщенная линия регрессии описывается двумя формулами: 1) S = 43,0921 Na/Mg+Sr+Mn - 10,1536 (интервал отношений 0,5-0,9); 2) S = 28,8273+1,6681 Na/Mg+Sr+Mn (для отношений >0,9), где S - соленость, ‰. Соленость по первой формуле определяется в пределах 13-30‰ с довольно большим стандартным отклонением (до 6,75%), по второй - в пределах 30-36‰ с точностью до 1,013%. Для оперативного определения палеосолености достаточно эмпирического графика, так как расхождение между соленостью, вычисленной по формуле и определенной по графику, не превышает 0,5%. Этот метод был апробирован на нижнемеловых устрицах с севера Средней Сибири [399] и плейстоцен - плиоценовых арктиках Исландии [387]. Установлена удовлетворительная корреляция с качественными моделями, основанными на литолого-палеоэкологических данных. Применение данного метода в палеоэкологии предполагает следующие допущения: 1) мезозойские устрицы и плиоценовые арктики ассимилировали присутствующие в воде катионы Na, Mg, Sr и Mn в тех же пропорциях, что и современные, 2) с момента захоронения изучаемой раковины до обнажения ориктоценоза отношение этих элементов оставалось постоянным.
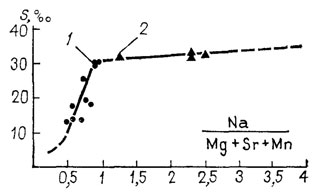
Рис. 93. Определение палеосолености по графику. Линия отражает зависимость содержания Na, Mg, Sr и Mn в раковинах устриц от солености воды. 1 - Crassostrea virginica, 2 - С. gigas
Другим методом определения палеосолености являются исследования карбонат-протеиновых отношений. Предполагается, что это отношение растет в кислых опресненных водах, поскольку усиливаются конхеолиновый слой и органическая матрица скелета, защищающие минеральную его составляющую от растворения при низкой рН, и падает в щелочных условиях повышенной солености, когда организм интенсивнее наращивает минеральную составляющую скелета. Выполненные к настоящему времени работы показали, что сказанное справедливо в отношении некоторых донных моллюсков. Так, кривая карбонат-протеиновых отношений по раковинам раннемеловых устриц на севере Средней Сибири демонстрирует четкую прямую корреляцию с кривыми палеобстановок и солености [400, рис. 1]. Однако эта кривая по рострам белемнитов дает обратную корреляцию с палеообстановками и палеосоленостью. Вероятно, защитный механизм белемнитов качественно отличался от такового у устриц. Следовательно, карбонат-протеиновое отношение находится под физиологическим контролем. Его использование для реставрации палеосреды бассейнов возможно лишь после экспериментальной проверки на той группе, которая подвергается испытанию.
Как показала практика, данные палеобиогеохимии и палеобиохимии не могут быть использованы в палеоэкологических исследованиях в чистом виде. Физико-химические методы должны привлекаться для реставрации отдельных факторов палеосреды только в комплексе с другими литолого-палеоэкологическими исследованиями. Положительная корреляция палеоэкологических и физико-химических данных значительно повышает достоверность количественных методов.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'