
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
4.4. Проблемы филогенетической классификации организмов (С. В. Мейен)
Многие общепризнанные виды, роды и другие таксоны организмов были выделены еще в те времена, когда естествоиспытатели не заботились о филогенетическом единстве таксонов. Систему организмов, складывавшуюся начиная со второй половины XVIII в., в додарвиновские времена называли естественной и противопоставляли искусственным системам, тождественным опредительским ключам [130]. После того как эволюционизм проник в систематику и стала развиваться филогенетика, появились различные представления о соотношении естественной системы и филогении. Ч. Дарвин считал филогенез наиболее правдоподобным объяснением естественной системы, хотя и не преувеличивал возможность реконструкции филогении. В "Происхождении видов" он высказал сомнение в том, что "геологу будущего" удастся доказать происхождение пород рогатого скота, овец, лошадей и собак от одного или нескольких первоначальных корней.
Точка зрения Ч. Дарвина унаследована теми из современных эволюционистов-систематиков, которые не настаивают на необходимости полного изоморфизма системы и филогении, но все же считают, что система должна быть согласована, совместима - consistent - с филогенией [2289]. Эту точку зрения, довольно популярную среди творцов и сторонников синтетической теории эволюции [390, 594, 1283, 2289, 2322], резко критикуют кладисты, считающие, что система должна быть строго филогенетической [1682, 1724, 1833]. В отношении естественной системы среди кладистов есть две точки зрения. Одни из них [1724] отрицают наличие какой-либо единственной привилегированной системы, помимо филогенетической, и все такие системы считают искусственными. Другие противопоставляют естественную систему филогенетической. По их мнению, "филогенетическая и естественная систематики различны коренным образом. Их задачи, методы, история и факторы прогресса совершенно различны и несовместимы. Для филогении многовековая работа естественной систематики малоценна. Для нее все сделанное до Дарвина нужно переделать заново" [499, с. 8]. Сходных взглядов на отношение системы и филогении придерживаются и противники филогенетической систематики [586, 975]. Они тоже считают филогению и естественную систему несовместимыми, но отдают предпочтение второй. Наконец, возможна точка зрения, близкая к дарвинской, но допускающая и иные, помимо общности происхождения, объяснения естественной системы [382, 589, 667, 1950].
Дискуссия между сторонниками этих точек зрения длится вот уже более ста лет [469, 526, 587, 1724, 2119]. Однако лишь в немногих работах подвергнута достаточно строгому анализу наша способность устанавливать филогенетические линии и филогенетическое единство таксонов. Нередко подробно рассматривались возможные таксономические интерпретации абстрактные филогенетических диаграмм, но меньше уделялось внимания тому, как получить таксоны, входящие в подобные диаграммы, и как установить, что их отношения преемственности образуют изображенное на диаграммах, а не какое-либо иное филогенетическое древо. Создается впечатление, что эти таксоны просто берутся из обычной естественной системы, где они устанавливались помимо филогенетических гипотез. После этого сходства и различия между имеющимися таксонами излагаются в терминах конвергенции, дивергенции и параллелизма, а дальше все зависит от того, каким сходствам будет придан смысл унаследованных, а каким различиям - смысл филогенетической дивергенции. Неудивительно, что когда дело доходит до графического изображения филогении, вариантов филогенетических древес получается несколько.
Еще меньше внимания в литературе уделяется тому, какие имеются способы проверки филогенетических построений и каковы критерии выбора между конкурирующими филогенетическими гипотезами. Обычно под проверкой филогенетического построения неявно понимают его устойчивость при введении новых таксономических признаков. Например, одно время было модно считать важным филогенетическим признаком аминокислотные последовательности цитохрома С. Считалось, что если данные по цитохрому С согласуются с прежними филогенетическими построениями, то эти построения подтверждены. Наоборот, когда пришли к выводу, что современные Magnoliaceae и Ranales по цитохрому С не могут стоять в основании системы двудольных [1364], то это рассматривалось как необходимость пересмотра филогении покрытосеменных.
В большинстве случаев подобные исследования имеют косвенное значение для филогенетических построений и лишь указывают на большее или меньшее сходство таксонов по тем или иным признакам. Чтобы от этих отношений сходства и различия перейти к филогенетическим построениям, надо указать, каким образом унаследованное сходство было отделено от независимо возникшего, а дивергенция - от полиморфизма, который можно назвать транзитивным.
Последний термин нуждается в пояснениях. С исторической точки зрения полиморфизм может быть двух видов - обратимым и необратимым. Необратимый полиморфизм означает векторизованное расщепление архетипа. Это - эволюционная дивергенция. Серьезной издержкой эволюционизма стало стремление рассматривать любое разнообразие в понятиях примитивности и продвинутости как свидетельство дивергенции. Например, печеночные мхи могут быть талломными и листостебельными. В литературе издавна идет дискуссия о том, какая из форм должна считаться примитивной, а какая продвинутой [668]. Двумешковая пыльца имеет различные по форме, размерам и углу наклона (по отношению к экватору) мешки. На основании изучения современных хвойных высказывалось мнение [2402], что более крупные мешки с меньшим углом наклона - признак примитивности. Но есть и иные точки зрения на примитивность или продвинутость определенных типов мешковой пыльцы. Подобные дискуссии переполняют литературу, поскольку интерпретация морфологического и иного разнообразия организмов в перечисленных раньше эволюционных понятиях считается главной задачей биологии.
Однако известно, что не всякий полиморфизм можно толковать в понятиях примитивности и продвинутости и считать проявлением эволюционной дивергенции. Таков, например, половой диморфизм. Хотя само появление пола - следствие дивергенции, но, раз появившись у предков современных многоклеточных организмов, половой диморфизм гамет, гаметангиев и их носителей уже не может интерпретироваться как дивергенция. Едва ли целесообразно рассматривать как дивергенцию проявления сезонного полиморфизма (цикломорфизма или альтернативного полиморфизма [1724]) изменения особей в ходе онтогенеза (плеоморфизм), модификационную изменчивость. Эти виды полиморфизма могут проявлять очень резко. Например, у десмидиевой водоросли Micrasteris pinnatifida описаны 26 различных морфологических форм [2306]. Широко известны модификации листьев у растений (гетерофилия), живущих то в воде, то на суше. Эти модификации обычно иллюстрируют на лютиках, череде и других покрытосеменных, хотя то же явление известно у мхов [1913] и водорослей [1766]. Не нуждается в пояснениях полиморфизм особей у колониальных насекомых (термитов, муравьев, пчел), мшанок, кораллов и т. п. Здесь невозможно рассматривать одни из особей колонии как примитивные, а другие как продвинутые. Приходится помнить и о полиморфизме гомодинамных органов в пределах одного организма. Краевые и средние цветки и семена в корзинке сложноцветного могут сильно отличаться морфологически.
По аналогии с гетерокарпией и гетероспермией [568] можно говорить о гетероспорангиальности (описана у карбонового папоротника Botryopteris globosa) [1990]. Напомним еще гетеростилию и иной полиморфизм цветков, иногда наблюдающийся в одном соцветии и часто сопровождающийся резким полиморфизмом пыльцы. Известны случаи, когда устойчивый полиморфизм пыльцы и плодов не сопровождается полиморфизмом цветков [1504]. Важно заметить, что пыльца варьирует по признакам, имеющим важное значение в палеопалинологии (по количеству борозд, скульптуре экзины и проч.). С этим явлением можно сопоставить диморфизм самцов у ряда насекомых и млекопитающих. Широко известен генетический полиморфизм популяций, устойчивость которого помимо прочего связана и с повторными одинаковыми мутациями, проявляющимися как в пределах вида, так и в разных видах. В связи с этим Д. Вуджер [2500] напоминал слова Д. Гексли [1769] о том, что две мутантных линии withe-eyed в двух видах дрозофилы не происходят от какой-либо общей линии withe-eyed.
Перечисленные и многие другие случаи внутривидового (или даже наблюдающегося в пределах особи) полиморфизма характерны тем, что здесь разнообразие передается в ряду поколений и переход от одних полиморфических модификаций к другим легко обратим. Такой "передаваемый" в чреде поколений полиморфизм и будем называть транзитивным.
Явление транзитивного полиморфизма широко известно, но обычно его ограничивают внутривидовым уровнем. Между тем транзитивный полиморфизм возможен и на гораздо более высоких таксономических уровнях. Например, в порядке Peltaspermaies в течение перми и триаса сохранялся полиморфизм листвы (по типу ее расчленения) и кладоспермов (пальчатонервных и двусемянных, или пельтатных, и многосемянных), причем то, что этот полиморфизм именно транзитивный, хорошо видно по ассоциирующим органам. Есть и другие палеоботанические примеры транзитивного полиморфизма [644, 1955]. В литературе в течение многих десятилетий обсуждался вопрос о первичности или вторичности травянистых и древесных форм покрытосеменных. В последние годы намечается тенденция не решать этот вопрос в такой форме. Допускаются встречные преобразования жизненных форм в разных таксонах и сохранение разнообразия по этому признаку в течение эволюции крупных таксонов [947, 2322]. Возможно, что именно к транзитивному полиморфизму следует отнести явление, описанное в литературе как итеративное формообразование [74, 648, 674, 975]. Речь идет о неоднократном появлении в геологической летописи сходных форм, например моллюсков типа Cardium и Pecten.
Однако наиболее ярким проявлением транзитивного полиморфизма на надвидовом уровне можно считать "гомологические ряды" Н. И. Вавилова, или "рефрены" (см. 4.1). Проявление рефренов не имеет верхнего таксономического предела.
Оно неспецифично для какого-то класса признаков. Ясно, что рефрены будут проявляться вне зависимости от того, как будут трактоваться филогенетические отношения между таксонами. Если рефрен устанавливается между двумя таксонами, находящимися в прямой филогенетической связи, то можно заключить, что описываемая этим рефреном изменчивость и есть транзитивный полиморфизм. Но если рефрены не имеют верхнего таксономического предела, т. е. проявляются в таксонах любого ранга, то снимается таксономический предел и с транзитивного полиморфизма.
Здесь неслучайно было уделено такое внимание транзитивному полиморфизму и его связи с рефренами. Дело в том, что главная трудность филогенетических реконструкций, а следовательно, и филогенетической систематики организмов заключается в том, чтобы отличить унаследованное сходство (единственно существенное для филогенетики) от независимо возникшего, а унаследованный (транзитивный) полиморфизм (он филогенетику не интересует) от необратимых (векторизованных) дивергентных изменений. Делались попытки решить эти трудности за счет интерпретации рефренов как следствия единой генотипической основы. Сам Н. И. Вавилов указывал, что чем ближе родственны таксоны, тем полнее повторность в их изменчивости. Это мнение Н. И. Вавилова затем неоднократно повторялось в литературе.
К этому тезису Н. И. Вавилова можно предъявить серьезные претензии. Одна из них чисто логическая. Дело в том, что близость таксонов не есть нечто заранее данное. Ее требуется установить, а не постулировать. Для этого надо, как уже говорилось, отделить независимо возникшее сходство от унаследованного. Соответственно рефрены, будучи сами проявлением сходства, сами же нуждаются в филогенетическом истолковании и не могут служить для него основой. В этом смысле показательно, что гомологическая изменчивость, наиболее полно изученная на родах культурных злаков, ни на шаг не приблизила нас к филогенетическим отношениям этих родов. Вторая претензия касается самой характеристики, данной Н. И. Вавиловым гомологическим рядам и имеющей важные недочеты. Н. И. Вавилов обратил преимущественное внимание на повторность признаков от таксона к таксону, но, по сути, прошел мимо того существенного момента, что повторяются не только отдельные признаки, но и способ их изменения. Инвариантны не просто признаки (точнее, мероны), инвариантно, а потому и закономерно, их преобразование; эта одинаково упорядоченная совокупность полиморфических модификаций и названа рефреном.
Эмпирический материал показывает, что изменчивость меронов одного класса следует рефреном вне зависимости от таксономического расстояния. Известны много случаев, когда рефрены выражены полнее у далекоотстоящих в системе таксонов, чем у соседних таксонов [1950]. При этом перестройка системы по такому сходству рефренов явно невозможна. Например, можно построить хорошие рефрены по строению листьев папоротников и пельтаспермовых (голосеменные). В то же время рефрены по строению листьев между пельтаспермовыми и хвойными будут несравненно менее полными, из чего никак не следует, что пельтаспермовые филогенетически ближе к папоротникам, чем к хвойным. Сейчас уже достаточно хорошо установлено, что предками голосеменных были прогимноспермы, которые произошли от тримерофитовых независимо от папоротников. К этому добавим, что меньшая повторность изменчивости между одной парой таксонов по сравнению с другой парой может быть связана всего лишь с разной изученностью таксонов.
Другой недочет более существен. Он касается трактовки Н. И. Вавиловым радикала, т. е. признаков, разделяющих таксоны с параллельной изменчивостью по прочим признакам. Получилось, что радикал - нечто неизменное в таксоне, позволяющее с абсолютной надежностью отличить его от других таксонов. Очевидно, признаки радикала и будут рассматриваться как результат дивергенции. Соответственно филогенетическая классификация должна строиться прежде всего по радикалам. Однако радикалы тоже подвержены изменчивости, образующей рефрены. Чтобы в этом убедиться, надо взять очень большое количество представителей каждого таксона и, главное, не отбрасывать в сторону данные по так называемой тератологической изменчивости см. 1.7. В литературе не раз указывалось на важное значение тератов (уродств) для морфологии и систематики. Обычно тератологические формы используются для гомологизации частей или расцениваются как атавизмы (когда систематик считает, что ему известен филогенез группы, и при этом в качестве тератологического появляется признак, бывший нормой у предкового таксона). Однако в подавляющем большинстве случаев остается в стороне важная особенность терат, известная еще биологам XVIII в. и детально и описанная Н. П. Кренке [533], который ввел особый "закон родственных отклонений" - правило Кренке [653, 1950, см. 4.1]. При детальном изучении тератологической изменчивости на массовом материале видно, что признак, считающийся уродством у данного таксона, встречается в качестве нормы у другого, часто очень близкого таксона (нередко у другого вида того же рода).
Поскольку признаки радикала наиболее константны в пределах таксона, то естественно, что их изменчивость как раз и расценивается как тератологическая. Правило же Кренке указывает, что такие тераты сближают данный таксон с тем, где "уродство" становится нормой. Иными словами, тератологическая изменчивость с учетом правила Кренке превращает признаки радикала в обычные признаки, подчиняющиеся, хотя и "с большим трудом", закону Вавилова и входящие в рефрены. Тогда оказывается, что таксоны, особенно близкие, отличаются не присутствием - отсутствием каких-то признаков (т. е. качественно), а лишь частотой встречаемости признаков одного набора (рефрена). Но частота встречаемости - количественное, а лучше сказать, статистическое отличие. Таким образом, можно сделать вывод, что таксоны, вопреки мнению Б. А. Вайнштейна [133], имеют статистическую природу. Было показано [666, 1950], что правило Кренке (как и рефрены) не имеет верхнего таксономического предела и действительно для таксонов любого ранга. Соответственно мы можем предполагать, что филогения не сводится к простой смене места таксона в признаковом пространстве, а, скорее, может быть описана как сдвиг частот встречаемости признаков на фоне очень медленно меняющегося признакового пространства. При этом само необратимое изменение признакового пространства чрезвычайно трудно зафиксировать.
Сказанное о природе таксонов и характере перехода между ними в свое время основывалось на сравнении морфологических признаков, причем исходным материалом служили преимущественно растения [1950]. Поскольку "ряды Вавилова" (а стало быть, и рефрены), транзитивный полиморфизм [75] и правило Кренке свойственны также животным и бактериям [382], есть основания считать, что таксоны всех организмов имеют статистическую природу, выраженную в большей или меньшей степени. Можно сделать предположение, что филогенез идет путем перехода потенциально присутствующего и актуально проявляющегося путем расширения потенций.
Подтверждение приведенному выше выводу о характере перехода между таксонами дают молекулярно-биологические исследования. Показано [1832], что электрофоретическая дистанция по белкам между человеком и шимпанзе в 25-60 раз больше, чем между расами людей, но примерно та же, что между видами-сиблингами (двойниками) дрозофил и некоторых млекопитающих. Результаты гибридизации ДНК примерно такие же. Виды лягушек и белок отличаются биохимически примерно в 20-30 раз больше, чем человека и шимпанзе. Из этих данных был сделан вывод, что в ходе эволюции, особенно с переходом ко все более высокоорганизованным существам, не столько меняется генетический код, сколько механизмы, контролирующие проявление генов. С этим выводом согласуется мнение, что макромутационные изменения происходят за счет системных мутаций, преобразований в "генах-регуляторах", а не в "структурных генах" [8, 527, 1483, 1650, 1664, 2106, 2417].
Таким образом, хотя закон Вавилова и мнение о связи "гомологических рядов" с генотипическим единством остаются в силе, при том что "гомологичность" изменчивости проявляется и безотносительно к генотипическому единству [647, 1081, 1950], но сам по себе этот закон (точнее, наличие рефренов) не только не расширяет возможности выявления филогенетических связей между таксонами, но, наоборот, сильно затрудняет или даже временами делает невозможным установление этих связей.
Сказанным не исчерпываются фундаментальные трудности, стоящие перед филогенетическими реконструкциями, а значит, и перед филогенетической систематикой. В последнее время появляется все больше данных о том, что поток генов не ограничивается внутривидовыми рамками. Описано много различных способов неполового переноса генетической информации [509, 510, 2513], и этот способ обогащения генофонда таксона за счет поступления генов со стороны, в том числе от сколь угодно далеких таксонов, уже используется для филогенетических гипотез [525, 2479].
Особое значение при этом придается вирусной трансдукции (трансгрессии) [1159]. Нетрудно заметить, что этим механизмом можно объяснить любые сходства и любое филогенетическое преобразование, вплоть до появления скелетов у разных групп организмов в низах кембрия [645]. Методологический недостаток этих гипотез, как и стремлений объяснить достижения человеческой цивилизации дарами "пришельцев", состоит в том, что нет научного способа их опровергнуть, т. е. фальсифицировать [835], как, впрочем, и собрать доказательства в их пользу. Это такой тип биологического механизма, который мог проявиться когда угодно, в любом таксоне, в любом свойстве организма. С общих филогенетических позиций этот механизм чрезвычайно интересен, поскольку он позволяет говорить не о раздельной эволюции генофонда разных таксонов, а об эволюции некоей единой геносферы [509]. Но для конкретной филогенетики и особенно для филогенетической систематики реальная возможность неполового переноса генетической информации означает крушение фундаментальных постулатов, принципиальную невозможность отыскать исторический источник тех признаков, которые лежат в основе характеристики таксонов.
Мы не останавливались на других трудностях филогенетической систематики, о которых не раз говорилось в литературе [382, 390, 526, 586, 587, 588, 1724, 2300 и др.] и которые имеют немаловажное значение. Нельзя сказать, чтобы все это не было известно приверженцам филогенетической систематики. Обычно предлагались те или иные способы обойти эти трудности (впрочем, следствия вирусной трансдукции и подобных механизмов для филогенетической систематики пока почти не обсуждались). В частности, возлагаются большие надежды на метод гибридизации ДНК [23, 1021]. Этот метод уже привел к некоторым парадоксам [526], заставляющим вводить контроль со стороны классических методов систематики. К тому же до того, как опираться на этот метод, надо убедиться в его действенности. Для этого его надо проверить на надежных объектах. К сожалению, таких объектов филогенетическая систематика предложить не может, особенно если речь идет о таксонах надвидового ранга. Ведь любая уже построенная филогенетическая схема делалась в обход перечисленных выше трудностей. По существу, методика гибридизации ДНК подгонялась под требования некоторой широко признанной системы каких-либо организмов. Неудивительно, что, будучи приложенной к другим организмам, полученная методика гибридизации давала несуразные результаты. Так было, когда методика гибридизации, отработанная на животных (в частности, на рыбах), была применена к однодольным растениям [23, 961]. Полученные связи родов радикально отличаются от всех предполагавшихся по другим методам. Это противоречие можно трактовать в пользу метода гибридизации ДНК, но можно обратить и против него. В самом деле, какие есть основания считать, что одна из систем рыб, к которой подгонялась процедура гибридизации, более "филогенетична", чем якобы дискредитированная система однодольных растений?
К сказанному добавим, что этот метод не в состоянии разграничить унаследованные кодоны от привнесенных, если имела место вирусная трансгрессия. Есть и чисто технические ограничения метода - его невозможно применить для музейного и ископаемого материала. Пока почти ничего не известно о внутривидовом полиморфизме с точки зрения этого метода. Наконец, его фундаментальным недостатком является то, что он дает валовое, не дифференцированное по отдельным "осмысленным" кодонам сходство ДНК. Вполне допустимо подозрение, что этот метод сравнения таксонов равносилен сравнению литературных произведений по суммарной общности применяемой лексики. Такая общность может дать интересный лингвистический материал, но едва ли позволит судить о более интересной для литературоведа идейной преемственности писателей.
В качестве критериев, важных для филогенетической систематики, указывались географическое распространение и состав паразитов (для самих паразитов - таксономия хозяев). Эти признаки важны для таксонов низкого ранга - видов, реже родов [1724], но и здесь они не могут играть решающую роль. Биогеографические и паразитологические данные сами основываются на знании таксономии соответствующих организмов и могут лишь ее корректировать методом "взаимного освещения". Биографические связи, кроме того, интересны для таксономии лишь при условии полноценного палеогеографического фона, обычно весьма гипотетичного. Паразитологические данные тоже не всегда надежны из-за возможной смены паразитом хозяина. Недавно были обнаружены грибы типа Albugo в карбоновых семезачатках Nucellangium [2337]. Современные грибы Albugo паразитируют исключительно на покрытосеменных и стимулируют такие же разрастания ткани (наподобие раковых), что и обнаруженные у карбонового голосеменного.
Из сказанного вполне можно было бы сделать вывод, что филогенетические реконструкции принципиально нереализуемы, и, следовательно, филогенетическая систематика принципиально невозможна. В некоторых случаях, например для определенных микроорганизмов [382], это, видимо, так и есть. Сомнительно, чтобы когда-нибудь удалось доказательно реконструировать филогению риниевых и зостерофилловых проптеридофитов с очень полно изученными эпиморфологическими и анатомическими признаками, поскольку основные признаки образуют у разных родов чуть ли не все мыслимые комбинации (рис. 121), отчего система этих родов приобретает облик многомерной решетки. По-видимому, не могут считаться филогенетическими, даже условно, схемы сходства конхострак или бивальвий, когда они основаны лишь на таких признаках, как внешний контур раковины, ее размеры и характер линий прироста.
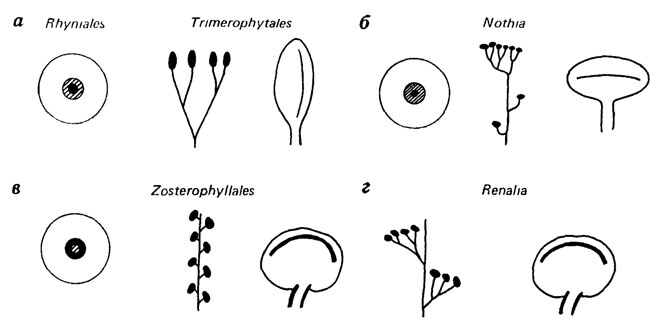
Рис. 121. Комбинативность признаков у девонских проптеридофитов (центрархная или экзархная протоксилема). Составил С. В. Мейен, оригинал. Спорангии эллиптические (а, б) или почковидные (в, г), раскрывающиеся продольной или поперечной щелью; терминальное (а, б) или боковое (в, г) положение спорангиев
Судьба филогенетических гипотез показывает, что всем им не удается избежать двух главных дефектов. Во-первых, они связывают филогенетически только уже известные таксоны и, как правило, перестраиваются, причем в главных чертах, при открытии новых таксонов. Это особенно характерно для филогенетических схем, построенных без учета палеонтологии. Такова была судьба предложенной А. Н. Северцовым филогении рыб [765], многочисленных филогении растений после открытия прогимноспермов и каллистофитовых, а также детального изучения разных палеозойских растений. Часто палеонтологические данные, хотя и не опровергают филогенетических гипотез, но ставят их под сомнение. Например, палеоботанические материалы не только не дали подтверждения, но и вступили в противоречие с весьма популярной среди ботаников "раналиевой гипотезой" происхождения покрытосеменных [см. подробнее: 1503]. Во-вторых, в филогенетических реконструкциях, даже подкрепленных палеонтологическим материалом, обычно смешиваются общие утверждения о филогенетической близости таксонов и конкретные утверждения о прямой филогенетической связи таксона. Только первые утверждения могут быть хорошо аргументированны, тогда как для вторых надо предъявить доказательства, что в будущем никогда не будет открыт таксон, еще более подходящий на роль предка (или потомка).
Приведенные трудности и обусловленная ими недостоверность филогенетических реконструкций отмечались многими исследователями, как защитниками, так и противниками примата филогенетических классификаций над иными. Осознание трудностей и дефектов любого подхода может вести к попыткам его усовершенствования или к отказу от самого подхода. Так случилось и с филогенетическими классификациями. Если В. Генниг [1724] и его последователи пытались изыскать новые критерии филогенетической близости организмов, то А. А. Любищев [586] нацело отвергал полезность филогенетики для систематики.
С методологической точки зрения обе позиции оказываются уязвимыми для критики. Радикальный кладизм В. Геннига и его школы не учитывает возможность того, что хотя бы в некоторых случаях филогения принципиально нереконструируема. Возможно, нам не следует "указывать пальцем" на подобные случаи, поскольку всегда может обнаружиться способ решения задач, сейчас кажущихся неразрешимыми. Поэтому есть риск неудачным примером скомпрометировать аргументированную позицию, вполне состоятельную теоретически. Эту позицию можно сформулировать так: теория систематики, даже если она выдвигает филогенетическую систематику как идеал, должна предусматривать принципы систематизации таксонов, не поддающихся филогенетическому анализу.
Кладизм В. Геннига не учитывает и того, что филогения реконструируется не для индивидов (филогения - не синоним генеалогии), а для таксонов. Когда мы относим к одному таксону сходные индивиды, занимающие дизъюнктный ареал, то историческое единство таксона, в том числе вторичность дизъюнкции, будет выводом, а не предпосылкой таксономического решения. Иными словами, кладизм не указывает универсальных филогенетических методов получения таксонов всех рангов, а занимается филогенетической интерпретацией таксонов, полученных существенно нефилогенетическими методами, и перестройкой системы уже имеющихся таксонов на основе некоторых филогенетических гипотез.
Методологические просчеты радикальных противников филогенетической систематики иные. Если отказываться от всех филогенетических принципов систематики, то придется перестраивать решительно всю систему организмов или искать какие-то нетривиальные доводы в пользу сохранения общепризнанных таксонов. Дело в том, что, даже отказываясь от филогенетичности таксонов, мы вынуждены учитывать генеалогические связи индивидов, т. е. филогению низшего уровня; по аналогии с микроэволюцией ее можно назвать "микрофилогенией". В противном случае исчезнут основания для объединения в один вид индивидов с ясно выраженным половым диморфизмом и иными проявлениями внутривидового транзитивного полиморфизма. С радикально антифилогенетической точки зрения для систематизации организмов не нужна ни географическая, ни стратиграфическая привязка материала. Можно собрать вместе все экземпляры, найденные когда-либо и где-либо, и сортировать их по признакам сходства и различия.
Ясно, что при этом будут пропущены виды-двойники, из-за межвидовых гибридов придется объединять вполне самостоятельные по прочим признакам виды. Особенно большие трудности возникнут при систематизации организмов, у которых таксоны хорошо различаются преимущественно по признакам одного пола. Возникнут и многие другие трудности, которые обычно не возникают только из-за того, что материал подбирается с учетом географической, экологической и (в палеонтологии) стратиграфической общности. Мы видим, что особи с трансгрессирующими признаками двух видов приурочены к местам перекрытия ареалов и относим этих особей к гибридам. Отмечается непременно сосуществование индивидов с разными половыми (или иными) признаками в одной колонии или популяции, причем между носителями разных признаков порой устанавливаются прямые генеалогические отношения. Тогда мы говорим о внутривидовом полиморфизме, не всегда отдавая себе отчет в том, что мы воспользовались "микрофилогенетическим" критерием единства вида. Таким образом, процедура установления таксонов неизбежно включает в себя некоторое количество филогенетических принципов. Если их изымать из теории, то требуется ясно сформулировать заменяющие их нефилогенетические принципы, особенно те, которые позволяют обойтись без "микрофилогенетических" гипотез и допущений. Этого радикальные противники филогенетических методов в систематике пока не сделали и даже не осознали самого существования проблемы.
Нельзя чрезмерно драматизировать и трудности филогенетических реконструкций, предъявляя к ним слишком сильные требования, а аргументируя невозможность самих реконструкций, обращать основное внимание на невозможность окончательного выбора между конкурирующими вариантами филогенетических древес. Фенетическая систематика, ориентировавшаяся на отказ от филогенетических гипотез, не может похвастаться ни преодолением трудностей, ни стабильностью дендрограмм (кладограмм), ни умением выбрать один метод их составления из множества конкурирующих и приводящих к разным результатам. Наконец, нельзя не упомянуть еще одну непреходящую непоследовательность, демонстрируемую антагонистами кладизма. Недостатки филогенетической систематики они критикуют, отталкиваясь от достижений именно филогенетики. Например, выступая против принципа монофилии таксонов, они утверждают полифилетичность таксонов. Однако для того чтобы судить о полифилии таксона, надо реконструировать отдельные монофилетические ветви, складывающиеся в полифилетический таксон. Логика требует выставлять против монофилетической доктрины не полифилетические взгляды, а отказ от какой бы то ни было зависимости систематики от филогенетики. Но такой отказ, как уже было показано, невозможен.
Итак, анализ проблем филогенетической систематики привел нас к парадоксу. Сначала мы выяснили, что создание системы, заведомо повторяющей филогенетическое древо, невозможно. Затем оказалось, что и отказ от филогенетической систематики невозможен. Чтобы решить этот парадокс, надо прежде всего осознать, что систематика - не тот вид деятельности, в которой мы начинаем работу "с нуля" и заканчиваем неким результатом, не подлежащим пересмотру. Систематика упорядочивает наши знания, которые никогда не были нулевыми (иначе не было бы предмета исследования) и никогда не будут окончательным (из-за невозможности полного познания объектов). Систематика всегда лишь перестраивает и дополняет уже имеющуюся систему. Она итеративно связана с мерономическими (морфологическими, физиологическими и экологическими) исследованиями [667], неокончательность результатов которых очевидна.
С другой стороны, подмена изменяющегося объекта его статичным образом - всегда лишь исследовательский прием. Подмена истории таксона суммированием свойств входящих в него индивидов - такой же исследовательский прием. Если мы хотим охарактеризовать таксон Homo, то мы можем включить в его диету (а это таксономический признак) и мамонта, и синтетическую икру. Но можно совершить грубую ошибку, если не учитывать, что в одну и ту же историческую эпоху люди не ели и того и другого. Признаки таксона не могут не меняться во времени, а совмещать диахронные признаки в недифференцированном хронологически диагнозе можно ценой лишь натяжек, оправдываемых только нашими субъективными целями.
То, чем фактически занимается филогенетика и есть прослеживание не статичного облака, а траектории таксона в признаковом пространстве. На филогенетических схемах это многомерное пространство вырождается в двумерную неструктурированную плоскость, а сложная в своей структуре траектория - в одномерную линию или двумерный контур. Чрезмерное графическое упрощение филогенетических отношений вуалирует самую суть дела. Сложные взаимоотношения таксонов в признаковом пространстве редуцируются до схемы, иллюстрирующей мнение филогенетика о некоей интуитивно суммированной близости или удаленности таксонов. Такие схемы вполне могут быть одним из результатов совмещенного филогенетического и таксономического исследований. Хуже, когда они воспринимаются как результат чисто филогенетического и объект дальнейшего чисто таксономического исследования. Тогда возникает путаница, а с ней и тот парадокс, о разрешении которого идет сейчас речь. Те же недоразумения возникают и тогда, когда фенетическая дендрограмма (не явно нагруженная "микрофилогенетическими" соображениями) подается как результат чисто таксономического анализа и объект для дальнейшей филогенетической реконструкции.
Выше говорилось о невозможности и чисто фенетического, и чисто филогенетического подходов к систематике. Хотя и неявно (неосознанно) оба метода так или иначе сочетаются. Иногда это сочетание воспринимается как методологическая ошибка и делаются обреченные на неудачу попытки размежевать оба подхода. На самом деле требуется разработка таких принципов систематики, в которых будет ясно показано совмещение подходов, относительная роль которых будет меняться в зависимости от ситуации. При этом в некоторых случаях роль филогенетических гипотез будет сводиться к минимуму, как бы принимать "нулевой вес". В других случаях филогенетические допущения могут определить структуру всей системы в ее самых важных чертах.
Сказанное можно иллюстрировать примерами. При классификации древнейших докембрийских микрофоссилий, когда неизвестно даже то, относятся ли они к эвкариотам или прокариотам, филогенетические соображения вырождаются в "микрофилогенетический" учет всего лишь приуроченности остатков к определенному слою. В остальном систематики могут опираться на очень узкий круг наблюдаемых признаков. Они группируют фоссилии по внешнему виду и придают полученным группам статус родов и видов, даже не зная, допустимо ли само приложение категорий вид и род к этим организмам. В сходном положении оказывается исследователь многих проблематичных или просто редких фоссилий (празинофитовых и эбридиевых водорослей, некоторых групп сколекодонтов и конодонтов, меловых цветков и др.), современных прокариот и некоторых современных монотипных таксонов, особняком стоящих в системе и не оставивших следов в геологической летописи.
Систематика других организмов, наоборот, вынуждена широко опираться на филогенетические представления, играющие важную эвристическую роль. Такова система позвоночных, особенно тех из них, по которым есть обширные палеонтологические материалы (млекопитающие, многие рептилии и амфибии, некоторые рыбы) В систематике высших растений филогенетический анализ оказался очень продуктивным для одних групп, например для высших таксонов голосеменных [660, 1955], но малопродуктивным для других (лептоспорангиатных папоротников, членистостебельных).
Опыт филогенетической систематики показывает, что анализ обширного современного материала, подкрепленный достаточно полными данными палеонтологии, приводит к получению филогенетико-таксономических схем высокой стабильности, т. е. выдерживающих привлечение все новых и новых признаков. Такое свойство систем соответствует главному критерию естественности, или (в данном случае это одно и то же) объективности.
Может возникнуть вопрос, а как же преодолевать перечислявшиеся выше многочисленные трудности, стоящие на пути реконструкции филогенезов. Ни общих, ни частных (на каждую трудность) рецептов здесь быть не может. Более того, речь должна идти не о рецептах, позволяющих преодолевать частные трудности, а о выработке принципов систематического исследования.
В общем случае необходимо решать вопросы в более широком контексте. Например, выше указывалось на возможность неполового переноса генетической информации - процесса не поддающегося доказательной расшифровке на палеонтологическом материале. Действительно, привести доказательства, что данное новообразование - результат трансдукции, нельзя. Но гипотеза трансдукции может стать сомнительной, если рассматривать более широкий класс новообразований в данном таксоне. Например, высказывалось предположение, что становление цветка покрытосеменных сопровождалось вирусным обменом генетическим материалом между разными голосеменными. В то же время исторические преобразования пыльцы и листьев древнейших покрытосеменных происходят постепенно по расходящимся линиям, ни в одной из которых не отмечается нарушающее постепенность привнесение признаков со стороны. Получается, что органы, по преобразованию которых есть богатый палеонтологический материал, эволюционировали без явных следов трансдукции. Ее действие приходится ограничить органами, преобразование которых плохо документировано в геологической летописи. Этим сопоставлением мы не отводим гипотезу о трансдукции, как заведомо несостоятельную, а лишь убеждаемся в том, что придавать этой гипотезе серьезное значение преждевременно. Сначала надо показать, что процесс преобразования цветка шел принципиально иначе, чем пыльцы и листьев.
Порой трудности филогенетических реконструкций могут оказаться непреодолимыми. Тогда надо научиться не обходить, не игнорировать их, а адекватно описывать сложившуюся таксономическую ситуацию. Таким образом, если мы не знаем филогенетических связей таксона и его места в системе, то именно это и нужно отразить на филогенетической или таксономической схеме. Само осознание и фиксация трудностей и неясностей - важный результат работы.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'