
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Биохорологические (собственно биогеографические) исследования
Биохорология (от греч. choros - место, пространство) и ее разделы фито- и зоохорология объединяют два основных направления: ареалологическое и флористико-фаунистическое. Исследования внешней среды не находят непосредственного отражения в их принципах, но полученные результаты привлекаются для выяснения действовавших факторов эндемизма и истолкования построенных карт-схем.
Ареалологическое направление. Рассматриваемое направление состоит в изучении географических ареалов таксонов органического мира, т. е. тех площадей земной поверхности или водных бассейнов, в пределах которых локализовано распространение каждого вида, рода, семейства и т. д.
Методике исследований ареалов современных организмов посвящена довольно большая литература [1069], позволяющая говорить о существовании учения об ареалах или ареалологии.
К сожалению, даже основы ареалологии еще недостаточно освещены в палеонтологической литературе, хотя они крайне важны для понимания краеугольных положений палеобиогеографии и самой сущности представлений о таксонах, поскольку географический ареал столь же характерен и специфичен для них, как и морфофизиологические особенности. Поэтому мы считаем необходимым хотя бы частично восполнить этот пробел.
Наибольшее внимание обычно привлекают ареалы видов как основной систематической единицы, без которой невозможно изучение ареалов вышестоящих категорий.
Ареал каждого вида объединяет все местонахождения - пункты, где он встречен, вне зависимости от количества экземпляров. Плотность заселения ареала представителями вида зависит прежде всего от распределения оптимальных и допустимых для него условий существования. Многочисленные данные биогеографов свидетельствуют, что ареалов, сплошь и равномерно заселенных особями вида, в природе не существует. Всегда обнаруживаются участки, занятые видом, и участки, где он в связи с непригодностью условий отсутствует. В результате образуется так называемое кружево ареала.
Участки видового ареала в связи с разнообразием их экологических особенностей характеризуются обычно различной плотностью заселения особями. Даже при высокой однородности экологических условий по всему ареалу в отдельных его местах нередко наблюдается максимальная плотность популяций. Эти места обычно рассматриваются как центры расселения. К периферии ареала часто приурочены наиболее уклоняющиеся, в том числе и архаичные формы. Это связывают [1739] с оттеснением их от центров расселения новыми и более прогрессивными формами. Распределение плотностей заселения данного ареала популяциями и особями вида складывается в некую общую картину, которая именуется структурой, или топографией ареала.
Таким образом, изучение видовых ареалов включает не только установление их географических границ, меняющихся во времени, но и выяснение их кружева и структуры, которые, как и границы ареала, анализируются с учетом особенностей самих организмов и различных факторов внешней среды. Уже из этого следует теснейшая связь био- и палеобиогеографии с экологией (палеоэкологией).
Было сказано, что подлинно сплошных, гомогенных, географических ареалов не существует. Тем не менее биогеографы допускают некоторую идеализацию явлений, зависящих от принятых критериев гомогенности, и пользуются термином "сплошной ареал". Практически сплошными принято называть ареалы, в границах которых экземпляры видов встречаются на всех соответствующих их природе местообитаниях, а популяции имеют возможность для свободного обмена особями. Необходимо отметить, что даже такие идеализированные сплошные ареалы встречаются крайне редко. Преобладают же политопные ареалы.
Дизъюнктивные (прерывистые) ареалы характеризуются обособленностью отдельных участков и разрывом связей между обитающими на них популяциями. Степень прерывистости ареалов может быть самой разнообразной. При расчленении ареала на большое количество участков возникают так называемые дисперсные ареалы, а когда от основной сплошной части ареала отделены один или несколько значительно меньших по площади участков - "островов", то образуются островные ареалы.
Возникновение дизъюнктивных ареалов не имеет однозначного толкования. Согласно одной точке зрения, оно связано с нарушением сплошности первичного ареала, согласно другой, - с независимым происхождением данного таксона в разных местах. В пользу второй точки зрения убедительные доводы в литературе приведены не были, тогда как примеров явного нарушения сплошности ареала много. Поэтому можно считать, что все дизъюнктивные ареалы первоначально были сплошными, а нарушения сплошности возникали позже, когда в местах разрывов условия оказывались неблагоприятными для жизни видов.
Размеры географических ареалов бывают очень различными - от точечного до космополитного, причем обширными ареалами обладают (и обладали в геологическом прошлом) преимущественно крупные таксоны и сравнительно немногие роды и виды. Таксоны и их сообщества с небольшими по площади ареалами, приуроченными к одной биохории низкого ранга, называют эндемичными.
Причинами эндемизма могут быть различные географические и экологические факторы. Иногда эндемизм возникает вследствие уникальности требуемых для таксона факторов среды, имеющихся лишь на небольшом участке Земли. В других случаях эндемизм оказывается результатом крупных палеогеографических перестроек, влекущих за собой сильное сокращение ареала, вплоть до небольшого остаточного участка; такое явление называют реликтовым эндемизмом.
Очевидно, что ареал рода и таксонов более высокого ранга существенно отличается по своей структуре от видового уже по той причине, что в нем отражены также структуры видовых ареалов (или ареалов иных таксонов подчиненного ранга). Сочетания ареалов подчиненных таксонов могут быть очень различными. Они перекрываются или примыкают один к другому, создавая нередко сложные картины. Поэтому нельзя рассматривать ареал таксона данного ранга как некую простую сумму ареалов подчиненных ему таксонов.
Участок родового ареала с наибольшим количеством обитающих видов данного рода называют центром разнообразия; обычно это наиболее древняя часть данного ареала. Здесь нередко происходят процессы образования автохтонных видов [196].
Исследования географических ареалов включают и классификацию их границ. Согласно В. В. Станчинскому и А. Г. Воронову [цит. по 196], границы ареалов подразделяются на подвижные и неподвижные. Подвижные границы, в свою очередь, могут быть прогрессивными (ареал увеличивается) и регрессивными (ареал уменьшается). Неподвижные, а точнее, медленно смещающиеся границы классифицируются по видам факторов, обусловливающим саму границу. Соответственно различают эдафические, конкурентные, климатические и непроходимые границы; под последними понимают географические преграды. Некоторая логическая непоследовательность этого деления очевидна, поскольку перечисленные факторы контролируют и подвижные границы, а выделенные типы факторов перекрываются (например, географические преграды являются в конечном счете и эдафическими; климатические факторы влияют на эдафические и конкурентные и т. д.). В большинстве случаев можно говорить не о видах границ, соответствующих этим или другим факторам, а о разных свойствах (аспектах) границ, каждая из которых контролируется сочетанием разных факторов.
Все приведенные основные положения ареалологии в общем приложимы и к палеобиогеографии. Однако их необходимо дополнить особенностями, свойственными исключительно ареалам ископаемых форм.
При исследовании ареалов и их изменений на основе палеонтологического материала наталкиваются на неполноту геологической летописи, в том числе на фрагментарность и разнообразие форм сохранности ископаемых остатков, затрудняющие, а подчас и исключающие привязку окаменелостей к таксонам, выделяемым по целым организмам. Особенно трудно идентифицировать остатки растений, которые могут быть представлены листьями и их фрагментами, древесиной, отпечатками коры, миоспорами и т. д. (см. 4.1).
Древнейшие достоверные континентальные отложения имеют силурийский возраст. Мы не знаем, была ли заселена суша в силуре и до него; возможно, что на древнейших континентах из-за отсутствия развитой растительности и нерегулируемости стока вод не образовывалось стабильных захоронений ископаемых остатков. Если так, то мы в принципе лишаемся возможности не только прослеживать ареалы древнейших наземных организмов, но и вообще встретить их остатки. Еще сложнее обстоит дело с изучением древних океанических организмов и их ареалов.
Поскольку прямое прослеживание ареалов на ископаемом материале далеко не всегда возможно, большое значение приобретают косвенные методы, состоящие в привлечении геоморфологических, литологических, климатологических и других данных [410].
Очень важным для палеобиогеографической ареалологии является указание А. А. Борисяка [106, 108] о различиях между биологическим и палеонтологическим видами и соответственно их ареалами. Ареал палеонтологического вида представляет собой не плоскую фигуру, а некоторое объемное тело, отображающее пространственно-временное распространение вида, осложненное к тому же и его эволюционными изменениями. Правда, объемный ареал имеют и многие современные организмы, занимающие некоторый вертикальный интервал в толще воды, почвы или атмосферы. Но при этом на картах ареала этот объем "сплющивается" до двумерного контура, что обычно недопустимо в отношении изменчивых во времени ареалов ископаемых форм.
Естественно, что при изучении объемных ареалов стратиграфическими методами выявляют их последовательные сечения. Для этого требуется детальная корреляция разрезов на обширных площадях распространения видов. Палеобиогеографу, как правило, доступно картирование видового ареала не в синхроническом срезе, что соответствовало бы ареалу биологического вида, а лишь части объемного ареала, отвечающей времени накопления некоего стратиграфического подразделения. Д. У. Валентайн [2415] предложил называть суммарную объемную фигуру, отражающую общую картину изменений ареала палеонтологического вида во времени, "биопровинцией", а ее часть, соответствующую определенному этапу существования этого ареала, - "тейльпровинцией". Мы полагаем, что эти термины излишни, так как точно в том же значении употребляются термины "биозона" и "тейльзона" [635]. Это совпадение понятий при различии терминов не случайно. Биостратиграфические подразделения, если они понимаются не как хронологические единицы, а как объемные тела, всего лишь охарактеризованные таксоном или совокупностью таксонов безотносительно к изохронности границ этих тел, оказываются полными синонимами объемных палеобиогеографических подразделений [608, 613].
Каждый вид или таксон более высокого ранга проходит за время своего существования определенные фазы распространения, которые можно классифицировать. В частности, В. Н. Дубатолов и Н. Я. Спасский [352] предлагают различать четыре последовательные фазы: 1) неоэндемика, характеризующуюся появлением таксона в первичном ареале, или центре возникновения; 2) полирегиональную, отличающуюся расширением ареала на соседние регионы; 3) полипровинциальную, соответствующую наибольшему распространению таксона; 4) реликтоэндемическую, когда таксон сохраняется только на остаточных (реликтовых) участках, часто расчлененных и удаленных от первичного ареала.
Эта схема, с нашей точки зрения, нуждается в коррективах, особенно в отношении второй и третьей фаз. Целесообразно связывать фазы распространения таксона с общей биогеографической ситуацией. Тогда первой фазе будет соответствовать распространение таксона в одной биохории низшего ранга или даже ее части. Второй фазой можно считать распространение ареала на смежные биохории, причем время максимального распространения, т. е. охвата наибольшего числа биохории, можно при желании считать отдельной (третьей) фазой. Возвращение таксона в одну биохорию низшего ранга можно, следовательно, оставить в соответствии с четвертой фазой. Между двумя последними фазами (по аналогии со второй фазой) можно выделить еще одну фазу. Несомненно, изучение этапности формирования ареалов - очень важная сторона ареалологических исследований, совершенно необходимая для выяснения истории самих таксонов.
Значительное место в методике изучения ареалов занимает картирование. Для целей палеобиогеографии наиболее целесообразно применение точечного и контурного способов, описанных и проиллюстрированных А. И. Толмачевым [1069]. Каждая карта-схема, построенная с помощью любого из названных способов, отображает ареал на плоскости. Для получения же представлений об объемном ареале необходимо создать серию таких карт-схем по располагающимся друг за другом снизу вверх срезам, т. е. проследить последовательные изменения ареала. При изучении полиморфных видов карты-схемы ареалов (рис. 1) целесообразно сочетать со схемами структуры этих видов (см. 4.6, рис. 125), отражающими их дифференциацию на подвиды [611].
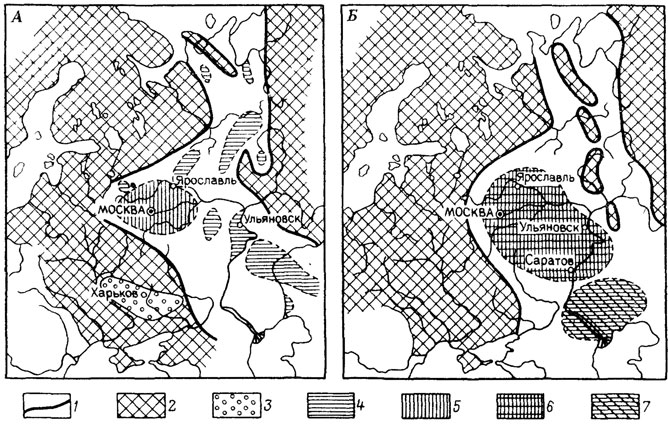
Рис. 1. Схематическая карта географических ареалов подвидов Russiella royeriana (d'Orb.). Волжский ярус Русской платформы. По В. П. Макридину. А - ареалы подвидов R. royeriana royeriana (d'Orb) и R. royeriana undorae Markid. во время Dorsoplanites panderi и в начале времени Virgatites virgatus; Б - ареалы подвидов R. royeriana royeriana (d'Orb.) и R. royeriana tenuis Makrid. в начале поздневолжского времени; 1 - береговая линия; 2 - области сноса; 3 - пестроцветные континентальные отложения; ареал подвида: 4 - дисперсный R. royeriana undorae Markid., приуроченный к иловым впадинам; 5 - R. royeriana royeriana (d'Orb.), приуроченный к области накопления песчаных и песчано-глинистых глауконитовых фосфоритоносных отложений сублиторали; 6 - R. royeriana royeriana (d'Orb.), приуроченный к области накопления песчаных глауконитовых фосфоритоносных отложении сублиторали; 7 - R. royeriana tenuis Makrid., приуроченный к области накопления карбонатных, местами глауконитовых отложений относительно углубленной области моря
При картировании ареалов родов и более крупных таксонов используют другие приемы. Рациональная методика была предложена Ю. И. Кацем [611]. Она заключается в обозначении, например, отряда знаком определенной формы (квадратом, ромбом и т. п.), каждого подчиненного ему семейства - определенной ориентировкой этого знака, а каждого рода, входящего в состав данного семейства, - определенной штриховкой или краской внутри нее. Можно, конечно, обозначать местонахождение буквами или цифрами [453], но такие карты-схемы менее наглядны.
Палеонтологи издавна интересуются географическим распространением таксонов и постоянно включают эти данные в их описания. Но регистрация местонахождений все еще редко завершается картированием ареалов и последующим их анализом, хотя имеющиеся работы [605, 611, 681 и др.] свидетельствуют о перспективности этого направления исследований.
Обобщение данных ареалологических исследований связано с применением ареало-генетического принципа, предусматривающего определенную последовательность действий. Прежде всего выявляется комплекс видов, родов и других таксонов, ареалы которых соответственно совпадают и изучаются их филогенетические отношения. При этом комплекс ряда относительно хорошо совпадающих ареалов именуется коареалом [1715] и фиксируется на карте-схеме как ареал-тип, т. е. некоторый реальный или воображаемый (абстрактный) ареал, отображающий множество конкретных совпадающих ареалов таксонов, которые объединяются в один комплекс [940, 941]. При выделении коареалов и ареал-типов не учитывается мозаичность реальных ареалов и допускается полное их совпадение. Чтобы перейти от хорологических подразделений - коареалов и ареал-типов - к единицам биогеографических подразделений, В. Н. Семенов [939, 940, 941] ввел понятие α, β и т. д. - регион (район), понимая под ним акваторию (территорию) неопределенного ранга, определяемую ареал-типом. При этом индекс "α" указывает на видовой, "β" - на родовой и т. д. уровни подразделений, принятые в данном исследовании.
Анализ полученных результатов позволяет оконтурить площади распространения эндемичных таксонов, причем оказывается, что каждый такой район (регион) характеризуется некоторыми связанными только с ним таксонами, имеющими общую историю, совокупность которых служит биогеографическим индикатором. Очевидно, что ареал вышестоящего таксона больше каждого из ареалов подчиненных ему таксонов, и он соответственно получит в схеме ареало-генетического районирования более высокий ранг.
Следует иметь в виду существенные отличия районирования на уровне видов и вышестоящих таксонов. Если схемы, составленные на видовом уровне, отражают преимущественно условия существования организмов в данный момент, то районирование на уровне более крупных таксонов содержит главным образом сведения исторического порядка [406].
Номенклатура биохорий пока еще между биогеографами не согласована. Большинство биогеографов-фаунистов предпочитают пользоваться иерархией, включающей в себя царство, область, подобласть и провинцию. Нет видимых причин, чтобы не принимать ту же иерархию и в палеобиогеографии [606, 607]. В фитогеографии чаще всего используется приведенная выше иерархия, но еще предусматриваются подцарство, подпровинция, округ и самая мелкая единица - подокруг. Обстоятельное обоснование этой иерархии для современной флористической структуры Земли приведено в книге А. Л. Тахтаджяна [1043]. Возможно, что она может оказаться полезной и при некоторых очень детальных палеобиогеографических исследованиях.
На начальных этапах био- и особенно палеобиогеографического районирования, когда выделение биохорий еще только намечается и ранг их в приведенной выше системе неясен, возникает необходимость в наименованиях свободного пользования, в качестве которых обычно выступают "районы" [606, 607, 1043] или "регионы" [938, 941]. Необходимость в терминах свободного пользования постоянно возникает при районировании лагунных и пресноводных бассейнов, так как здесь определение ранга биохорий в рассмотренной нами иерархии обычно крайне затруднено или практически невозможно. Применение же терминов свободного пользования позволяет избежать указаний стандартного ранга биохорий, когда он не вполне обоснован, хотя некоторые соотношения биохорий уже выявлены и их относительный (первый, второй и т. д.) ранг может быть назван.
При зоогеографическом районировании открытых морских бассейнов на основании бентосных организмов таксонами-индикаторами царств обычно оказываются эндемичные отряды, реже подклассы и классы, развитие которых длительное время протекало в их пределах; областей - семейства и подсемейства; подобластей - подсемейства и особенно роды и наконец, провинций - виды и подвиды [281, 282]. Согласно К. В. Беклемишеву и О. Н. Зезиной [62], зоогеографический ранг эндемичных таксонов пелагических организмов соответственно ниже, причем главную роль в обосновании выделения биохорий играют комплексы видов.
Приведенные соотношения обычно выдерживаются и при палеозоогеографическом районировании мезозойских и кайнозойских морей [606, 607, 611]. Однако в процессе развития биосферы, несомненно, происходили перестройки географической дифференциации органического мира, обусловившие и изменения ранга таксонов-индикаторов биохорий [606, 607]. О. А. Липина [580] склонна связывать изменения ранга биохорий с чередованием таласократических и теократических периодов. В целом вопрос о соотношении ранга биохорий и ранга их таксонов-индикаторов весьма сложен и остается дискуссионным.
С одной стороны, мы не можем слишком тесно связывать био- и палеобиогеографические представления с таксономическими, тем более, что последние, в свою очередь, порождают разногласия между исследователями. Во всей систематике происходит непрерывное повышение ранга таксонов, но далеко не всегда это отражается в ранге биохорий. Это хорошо видно в современном флористическом районировании Земли. Основные фитохории были очерчены еще более полувека назад, и дальнейшие перемены связаны не с изменениями таксономических взглядов (с тех пор многие семейства стали рассматриваться как самостоятельные отряды, подклассы и даже классы), а с более полным знанием распределения таксонов в пространстве. В то же время нецелесообразен и отрыв ранга биохорий от ранга приуроченных к ним таксонов, поскольку он систематически и обычно не без успеха используется как в био-, так и в палеобиогеографическом районировании.
По-видимому, решение этой проблемы надо искать в следующем. Сама возможность био- и палеобиогеографического районирования предопределена тем, что распределение организмов в биосфере Земли не континуально, а образует некоторую структуру, причем распространение разных таксонов скоррелировано в некоторых пределах. Более того, в этой структуре различается соподчинение подразделений, их иерархичность. Эмпирический подход привел к выделению конечного числа членов в этой иерархии; в самых полных списках их число не превышает восьми. И эта структура биосферы, и ее иерархичность были установлены с помощью анализа распространения таксонов разного ранга.
Если бы дело ограничивалось лишь этим, то можно было бы поставить ранг биохорий и ранг таксонов в прямую связь. Однако схемы районирования получили дополнительное обоснование в других наблюдениях, в том числе климатических (палеоклиматических), ландшафтно-географических и т. д. Оказалось, что биохорий - нечто большее, чем просто комплексы ареалов каких-то таксонов. Как уже говорилось, границы биохорий увязываются с эдафическими, климатическими и иными факторами. Были выявлены некоторые границы высшего ранга, контролируемые планетарными причинами (так, особенно высокий ранг имеет биогеографическая граница, совпадающая с границей морозного и безморозного климата). Такие границы, очевидно, уже не зависят от наших таксономических представлений. Кроме того, границы низшего ранга оконтуривают участки с высокой гомогенностью их населения, и смена таксономических взглядов ничего не изменит в ее оценке. Опыт свидетельствует, что повышение ранга многих таксонов не привело к увеличению числа членов в иерархии биохорий. Если высшие и низшие по рангу биохорий оказались, таким образом, фиксированными, как фиксировано и число рангов, то дальнейшие изменения в ранге таксонов уже не будут существенно влиять на систему биохорий.
Иное дело, когда меняются представления об объеме таксонов и соотношениях, в том числе филогенетических, между ними. Когда выяснилось, что растения, доминирующие в верхней перми Русской платформы, принадлежат не глоссоптеридиям, свойственным Гондване, а пельтаспермовым птеридоспермам (в перми Гондваны их нет), то резко изменились палеобиогеографические представления. В некоторых случаях и изменение ранга характерного таксона биохории без изменения ранга прочих таксонов может изменить представление о ранге самой биохории. В любом случае вопрос о ранге биохории должен решаться не путем формального учета рангов населяющих только ее таксонов, а путем анализа всей картины районирования обширной территории (акватории) за достаточно большой промежуток времени. Критика ареало-генетического подхода [639, 973, 974 и др.] справедлива, если она имеет в виду не сам этот подход, как таковой, а его издержки, порождаемые чисто формальным отношением к рангу таксонов, вовлекаемых в построения.
Есть и такие критические замечания в адрес ареало-генетического подхода, с которыми согласиться трудно. Так, В. А. Собецкий [973, с. 71] пишет: "Недостатком ареал-генетической концепции биогеографического районирования является то, что она предполагает столько схем районирования, сколько групп организмов существовало или существует на земном шаре... В конечном итоге такой подход может привести к отказу от признания объективного существования таксономических категорий биогеографии...". Приведенное мнение возвращает нас к давно пройденному этапу, когда А. Якоби [1786] в противовес сторонникам "классического" направления - А. Уоллесу и другим утверждал, что районирование не может основываться на многих группах организмов, так как каждая из них обладает свойственными только ей способностями к преодолению географических и экологических барьеров. Дискуссия на эту тему продолжается и сейчас, хотя есть все основания считать ее в основном исчерпанной. Составление схем районирования по отдельно взятым группам представляет собой с историко-научной точки зрения лишь первый этап накопления материалов, сменяющийся затем созданием схемы, основывающейся на учете данных по ряду групп организмов, близких друг другу в экологическом отношении [607].
Безусловно, схема районирования, построенная с учетом ареалов существенно разных таксонов, будет сильно отличаться от схем, полученных для представителей одного таксона. Поэтому создание схем, отражающих закономерности географического распространения комплекса таксонов, принадлежащих к разным группам организмов, представляет собой более сложное, синтезирующее ареалологическое исследование. Такой подход, вскрывающий специфические черты географической дифференциации каждой группы организмов и в то же время возможности их комплексирования, приводит к познанию более значимых биогеографических закономерностей и, следовательно, к установлению реальности прослеживаемых ареало-генетических биохорий.
Синтез данных по разным группам организмов обращает нас к проблеме веса признаков, которая стоит одинаково не только для всех направлений биогеографии, но и для науки в целом. Взвешивание признаков и есть та сторона исследования, в которой проявляется опыт и талант исследователя, когда дело доходит до интерпретации данных. Не углубляясь в эту общенаучную проблему, отметим лишь, что при определении веса признаков мы обычно просто заимствуем его из прошлого опыта и оставляем тем же до тех пор, пока не получим противоречивой картины
Флористико-фаунистическое направление. Рассматриваемое направление возникло в биогеографии раньше других [727]. Оно все чаще находит применение и в палеобиогеографии, хотя сталкивается здесь с большими трудностями.
Наиболее распространенный способ этого районирования как в био-, так и в палеобиогеографии предусматривает выяснение различий в систематическом составе флор и (или) фаун, населявших смежные районы, которым в зависимости от их масштабов придается определенный ранг.
Каждая современная или ископаемая флора и фауна представляют собой весьма сложное, исторически сформировавшееся явление, составные элементы которого тесно взаимосвязаны друг с другом и окружающей внешней средой. Поэтому их изучение предусматривает прежде всего выяснение систематического состава и биогеографической структуры, под которой понимают взаимосвязанную систему групп (ассоциаций, комплексов) видов и других таксонов, развившихся в пределах данной территории или акватории различными путями. Естественно, что для характеристики флоры и (или) фауны и занимаемой ими биохорий наиболее важное значение приобретают региональные формы, возникшие более или менее одновременно и существующие (существовавшие) здесь автохтонно, в отличие от космополитных и викариантных форм, особенно тех, которые проникли сюда позже из соседних районов.
Следующий шаг, имеющий исключительно важное значение в познании истории формирования флор, фаун и их биохорий, состоит в генетическом анализе входящих в их состав видов, родов, семейств и вышестоящих таксонов.
Кроме районирования по составу флор и (или) фаун может быть использован способ районирования по степени значимости границ выделяемых биохорий, приводящий к существенно иным результатам [941].
Поскольку систематический состав каждой флоры и фауны обычно очень разнообразен, для объективной характеристики его качественных изменений, как и для анализа эффективности биогеографических границ, большое значение приобретает статистический метод, заключающийся в применении разнообразных эмпирических формул [1425], в составлении таблиц, диаграмм и т. п.
Статистический метод используется в биогеографии уже сравнительно давно, а в последние 10-15 лет все чаще проникает и в палеобиогеографические работы [402, 1118, 1119, 1129, 1405 и др.]. Появляются специальные исследования, посвященные оценке эффективности применения некоторых эмпирических формул [1572, 1723] и разработке новых способов определения флористического и (или) фаунистического сходства [2126].
Обоснование выделения районов (регионов) составляет лишь одну сторону флористико-фаунистических исследований. Другой не менее сложной задачей является определение их биогеографического ранга. На примере меловых двустворчатых моллюсков Э. Г. Кауфман [1818] разработал основывающуюся на статистическом методе схему выделения биохорий, согласно которой (без учета космополитов) царства содержат 75% и более эндемичных родов, области 50-75%, провинции 25-50% и т. д. Но пользовавшийся ею А. Д. Буко [121] указал, что классификация Э. Г. Кауфмана ведет к ряду несоответствий, вследствие которых возникают большие номенклатурные трудности, особенно при изучении развивающихся биохорий.
С нашей точки зрения очевидно, что решение вопроса о ранге биохорий не может основываться на одном отдельно взятом и тем более формально используемом методе, а связано с сопоставлением всей суммы данных, характеризующих каждый из оконтуренных районов (регионов), в том числе, естественно, и результатов статистических подсчетов. Определенный интерес в этом отношении представляет сравнение критериев выделения современных и древних биохорий одинакового ранга [1405].
Не вызывает сомнений, что по мере накопления материала значение статистического метода и в целом формализации и математизации био- и палеобиогеографических исследований будет быстро возрастать. Но столь же несомненна недопустимость механического обращения со статистическими данными без анализа исторически сложившихся родственных взаимоотношений между видами сравниваемых флор и (или) фаун.
Уже говорилось, что выделение флористических и фаунистических подразделений - царств, областей, подобластей, провинций и других возможно только при охвате исследованиями обширных пространств материковой суши или морских (океанических) акваторий. Поэтому стремление некоторых палеонтологов, располагающих материалом по сравнительно ограниченным площадям, "находить" здесь всю эту иерархию биохорий неизбежно приводит к серьезным ошибкам и прежде всего - к сильному завышению их ранга.
С другой стороны, в настоящее время недостаточно внимания уделяется решению обратной задачи, а именно, уточнению положения исследуемых местностей в уже имеющейся общей схеме палеобиогеографического районирования. Между тем такие исследования приобретают важное значение в разработке более детальных схем.
Приведенные нами общие положения флористико-фаунистического направления не должны создавать ложного представления о возможности использования совершенно идентичного подхода к биогеографическому изучению как растений, так и животных, а тем более их ископаемых остатков. Разработка методики этих исследований составляет одну из проблем палеобиогеографии.
Еще более глубоким оказывается различие между биогеографией суши и моря. Л. А. Зенкевич [408, с. 249] по этому поводу писал: "...биогеографически подразделения Мирового океана и материковой суши не могут быть едины. Топография отдельных частей Мирового океана и материковой суши, связи между ними, факторы, обусловливающие распределение организмов на суше и в море, вертикальный диапазон заселенности той и другой среды (плоскостной на суше и объемный в океане), наличие пелагической жизни в морях и отсутствие аналогичного явления на суше и многое другое не дают возможности строить единые биогеографические схемы".
Отмеченные Л. А. Зенкевичем особенности распространения морских организмов обусловливают необходимость раздельного проведения биогеографических исследований литорали и других зон морей и океанов. Более того, их районирование, как правило" не может основываться на одновременном учете планктонных" нектонных и бентосных организмов, распространение которых контролируется разными факторами. Если распределение планктона зависит главным образом от освещения, температурных инверсий, градиентов плотности воды, силы и направления течений и т. п. [2511], то в формировании бентосных сообществ большую роль играют глубины, характер субстрата, температурный и газовый режим придонных вод и т. д. Естественно, что сказанное совершенно необходимо принимать во внимание и при палеобиогеографическом районировании, хотя нельзя не признать, что палеонтологи все еще часто испытывают серьезные затруднения при определении принадлежности пород и приуроченных к ним остатков организмов к той или иной батиметрической зоне моря.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'