
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Экосистемные исследования
Данные исследования заключаются в выяснении географического распространения организмов в непосредственной связи с условиями их существования и последующем сравнительно-географическом изучении разнообразных экологических систем. Поэтому в отличие от биохорологических исследований экосистемное районирование носит значительно более отчетливо выраженный синтетический характер и предусматривает выделение районов, различающихся условиями внешней среды и находящимися в единстве с ними комплексами растительных и животных организмов.
Экосистемные исследования проводятся в трех основных направлениях: климатобиогеографическом, биофациальном и ландшафтно-географическом.
Климатобиогеографическое направление. Данное направление расположено на рубеже био- и палеобиогеографии соответственно с климатологией и палеоклиматологией и занимается изучением географического распределения организмов в зависимости от влияния климатических факторов.
Существование связи географического распространения растений и животных с климатом было установлено давно. В биогеографическую науку это одно из ее краеугольных положений вошло во второй половине XVIII столетия под влиянием трудов Ж. Бюффона [727]; в наше время оно известно как закон географической зональности В. В. Докучаева - Л. С. Берга.
В палеофитогеографии данные о распределении фитохорий сочетались с палеоклиматическими реконструкциями с самого начала. О зависимости палеофлористических смен в разрезе от климатической эволюции писал Ад. Броньяр [1381]. Он же отмечал, что однообразие ископаемых флор на разных материках свидетельствует о сходстве палеоклиматов. После того как были получены данные о палеофлористической дифференциации в позднем палеозое (было установлено своеобразие гондванских флор, тогда же было распознано ледниковое происхождение развитых на Гондване тиллитов) и кайнозое, палеоклиматические интерпретации стали непременной составной частью палеофлористического районирования. Именно такое сочетание подходов имеет место в палеофитогеографических исследованиях О. Геера, Р. Зейллера, Э. Унгера, Д. Давсона, А. Энглера и других ботаников и палеоботаников прошлого века.
Первые попытки палеоклиматозоогеографических исследований принадлежат Ф. Ремеру [2171]. Современная литература, посвященная этому направлению, насчитывает сотни наименований, причем вспышка в его развитии, особенно отчетливо обозначившаяся в течение последних 10-15 лет в морской палеозоогеографии, в большой мере связана с разработкой проблем биостратиграфии [86, 94, 545, 574, 579, 676, 723, 876, 877, 878, 1227, 1228, 1229, 1230, 1231 и др.].
Основной единицей районирования в рассматриваемых исследованиях чаще всего считают "биогеографический пояс" [2444], хотя некоторые специалисты [878] предпочитают термин "надобласть". Под биогеографическим поясом в палеобиогеографии понимают подразделение, флора, фауна и отложения которого указывают на принадлежность к определенной климатической зоне [1227, 1228].
Изменениями географической широты обусловлены в настоящее время наиболее общие (глобальные) изменения систематического состава растительных и животных организмов. Однако не следует упускать из виду, что биогеографическая зональность возникла не сразу, а развивалась длительное геологическое время. Поэтому положение и контрастность границ, состав флор и фаун биогеографических поясов претерпевали в течение фанерозоя многочисленные и сложные изменения, обусловленные эволюцией климата, органического мира и палеогеографической обстановкой в целом. Следовательно, исторический подход к климатобиогеографическим построениям столь же важен, как и в ранее рассмотренных направлениях исследований.
Для подтверждения сказанного достаточно сослаться на весьма тщательные исследования В. Н. Сакса [903], свидетельствующие, в частности, что в триасовых морях северного полушария Бореальный пояс не прослеживается и его возникновение датируется лишь средней юрой. Если в юре граница Бореального и Тетического поясов совпадала с климатической зональностью, то южнее зоогеографические границы приобрели ориентировку, близкую к меридиональной, и увязать их с различиями в температурном режиме невозможно. Наконец, в течение почти всего мезозоя в южном полушарии вообще отсутствовал аналог Бореального пояса.
Анализ фактических материалов свидетельствует, что в течение большей части фанерозоя удается проследить наличие экваториального климатического (и биогеографического) пояса, характеризующегося богатыми и таксономически разнообразными флорами и фаунами, и двух высокоширотных поясов с относительно более бедными и однообразными флорами и фаунами, обладающими иногда заметным сходством [1086].
Необходимо помнить, что и реакция различных групп организмов на явление географической зональности неодинакова. Наиболее чутко на него реагируют наземные флоры и фауны, в связи с чем на суше биогеографические пояса выражены значительно более резко, чем в Мировом океане. В свою очередь, для прослеживания поясов в морских бассейнах особенно важными оказываются пелагические фауны, тогда как бентосные шельфовые организмы более тесно связаны с эдафическим и другими фациальными факторами, а глубоководные - находятся под косвенным воздействием климатических изменений либо вообще вне их. Они дают о себе знать в батиали через посредство структуры водных масс [1010], практически не затрагивая лишь олиготрофные области. Естественно, границы между смежными биогеографическими поясами выражены здесь значительно менее отчетливо, чем в зоне мелководья.
Если для существования наземных растений и животных большое значение имеют все климатические факторы - инфляционный режим, тепло и атмосферная влага, то для морских животных наиболее важным является температурный режим. Позвоночные суши, хотя и испытывают более сильное влияние климата, чем морские животные, обладают в силу более высокой организованности способностью в той или иной степени избегать неблагоприятных воздействий климата путем перемещений, выработки особого поведения и т. п. [954].
Все сказанное, несомненно, имело место и в геологическом прошлом и, следовательно, должно как-то учитываться в палеоклиматобиогеографических исследованиях.
Так или иначе, огромная роль климатических факторов в географическом распространении и в целом в эволюции жизни на Земле бесспорна. Это и определяет важность дальнейшей разработки рассматриваемого направления. Нельзя, однако, согласиться с предложением ряда специалистов [797, 1227, 1228 и др.] рассматривать биогеографические пояса в качестве наиболее крупных подразделений палеобиогеографического районирования вообще. Мы уже видели, что пояса представляют собой биохории, контролируемые климатом. В то же время биохории высшего ранга в хорологическом районировании - царства, хотя и имеют определенные климатические отличия (как и любые другие биохории), но их границы и соотношения на палеоклиматическом фоне таковы, что считать их подразделениями преимущественно климатической природы нельзя. Тем более это касается биохории низкого ранга, возникновение которых связано с влиянием не только климатических, но многих других факторов эндемизма. Для всех организмов, вне зависимости от условий их существования, таковыми являются: географические преграды, конкуренция в борьбе за пространство и пищу и т. д.; кроме того, для растений суши - характер рельефа и другое, а для морских животных - глубина, соленость и газовый режим бассейнов, действие течений и т. п. Естественно, что это не позволяет рассматривать пояса в одной иерархии с упоминавшимися ранее биохориями [580, 606, 607].
Исходя из сказанного, вызывает возражение и своеобразная точка зрения В. А. Собецкого, который хотя и именует "биогеографический комплекс высшего ранга" областью и использует для обоснования его выделения не только климатический критерий, тем не менее пишет: "Широтные границы областей определялись климатической зональностью, меридиональные - распределением материков" [973, с. 72]. Остается неясным, почему, скажем, наличие непреодолимых географических преград и глобальных океанических течений не могло оказать существенного влияния на положение широтно-ориентированных рубежей области?
Номенклатура климатобиогеографических подразделений нуждается в дальнейшем уточнении и детализации. С этой точки зрения представляют несомненный интерес работы В. Н. Семенова [939], предложившего новую широтно-зональную номенклатуру прибрежной морской биоты, применимую и в палеобиогеографии, и К. Н. Несиса [743].
На фоне зональных явлений все более отчетливо прослеживаются случаи азональности, состоящие в долготной дифференциации широтно-ориентированных биохории. Если в биогеографии данное явление, обусловленное существованием материков и океанов, меридионально протягивающихся горных цепей и т. д.. известно давно, то в палеобиогеографии оно изучено весьма недостаточно; в частности, в палеофлористике оно было описано впервые совсем недавно [325]; подробнее об этом см. 5.3.
Согласно Ю. И. Чернову [1039], широтным зональным биохориям должны соответствовать типы животного и растительного населения, а азональным - фаунистические и флористические комплексы, связанные общностью происхождения. Из этого следует, что детальное палеобиогеографическое районирование позволяет поэтапно проследить районы возникновения, пути и характер процессов расселения организмов и т. д. [2371]. В связи с разнообразием условий и масштабов рассматриваемых явлений соотношение азональных биохорий - стран, секторов с широтно-зональными подразделениями могут быть различными. В качестве одного из примеров приведем карту-схему, заимствованную из работы И. А. Добрускиной (рис. 2).
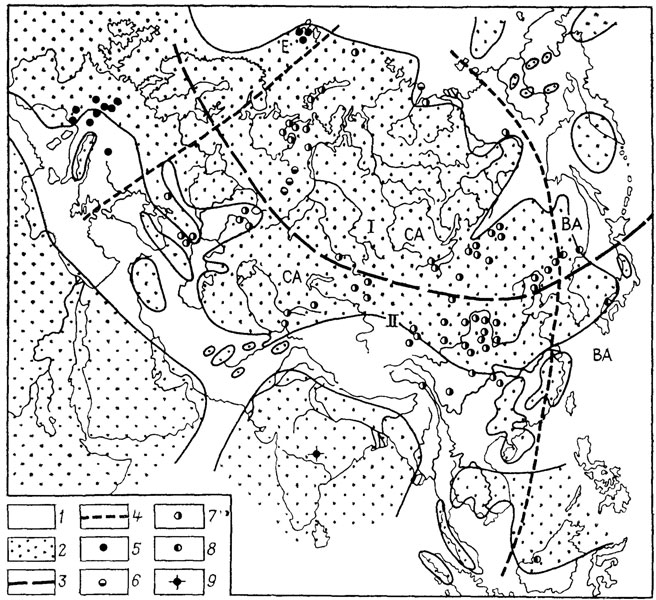
Рис. 2. Фитогеографическая зональность в середине триасового периода. По И. А. Добрускиной с дополнениями. I - Сибирская палеофлористическая область; II - Европейско-Синийская палеофлористическая область Лавразиатского царства, Е - Европейский сектор; СА - Среднеазиатский сектор, ВА - Восточноазиатский сектор. 1 - море; 2 - суша; границы; 3 - палеофитогеографических областей, 4 - палеофитогеографических секторов; местонахождения флор: 5 - европейского типа, 6 - уральского типа, 7 - среднеазиатского типа, 8 - восточноазиатского типа, 9 - Малери
Биофациальное направление. Данное направление для палеобиогеографии традиционно. Оно состоит в изучении фациальной приуроченности организмов. Его формирование связано прежде всего с трудами основоположника учения о фациях А. Грессли [1672], а также И. Вальтера [2452, 2453], О. Абеля [1248, 1249], Р. Гессе [2452, 2453], Р. Ф. Геккера [223, 224, 226 и др.], Е. А. Ивановой [419, 421 и др.], Б. П. Марковского [622] и многих др.
Основным подразделением, выделяемым при палеоэкологических исследованиях, является "биологическая фация", или сокращенно "биофация". Однако, как это часто бывает с основными понятиями, его формулировка связана с большими трудностями. Суть проходящих дискуссий достаточно полно отражена в работе В. А. Собецкого [973]. Чтобы сузить круг спорных положений, мы предпочитаем различать понятия современной и ископаемой биофации и в дальнейшем говорить преимущественно о последней.
С нашей точки зрения ископаемая биофация - это совокупность остатков организмов, приуроченная к определенной фации, т. е. к осадочным породам определенного литологического состава; в сочетании фоссилий с последними запечатлена исторически сложившаяся и экологически (неразрывная связь биотических и абиотических условий осадконакопления и существования рассматриваемого комплекса организмов. Принимая эту формулировку, мы не должны упускать из виду, что ископаемые биофации не отражают на синхронной поверхности среду того или иного участка Земли, а оказываются лишь приближенными их реконструкциями [973].
Методика рассматриваемого способа районирования разработана Р. Ф. Геккером и весьма успешно применима им при изучении разнофациальных толщ Главного девонского поля [223], а затем совместно с А. И. Осиповой и Т. Н. Вельской в исследованиях палеогена Ферганы [235]. По существу, она представляет собой комплексные палеоэколого-литологические исследования целых бассейнов геологического прошлого (или их крупных частей), либо других палеогеографических регионов с охватом значительной части их геологической истории [226]. Поскольку характер и распределение биофаций в пространстве и времени испытывают изменения, причем порой очень частые, одним из краеугольных положений этой методики является исторический подход к анализу разрезов и толщ, вмещающих окаменелости. С особенно большими сложностями обычно связаны исследования биофаций континентальных и лагунных отложений. Во многих случаях даже при тщательном подходе не удается закартировать отдельные биофации и ограничиваются лишь фиксацией их комплексов. Поэтому вопрос о классификации и номенклатуре биофациальных комплексов заслуживает особого внимания.
Ландшафтно-географическое направление. Основными подразделениями этого направления районирования принято считать биогеоценозы и палеобиогеоценозы, выделением которых занимаются соответственно физическая география и палеогеография, задачи же био- и палеобиогеографии - изучение приуроченных к ним комплексов организмов.
Под биогеоценозом понимают неделимый природный комплекс, представляющий собой совокупность живых и неживых компонентов наиболее однородного участка территории или акватории. Некоторые биогеографы [196] склонны отождествлять "биогеоценоз" с "современной фацией". Ландшафтом же обычно называют конкретную региональную географическую единицу, образованную неоднократно повторяющимися биогеоценозами или сходными совместно развивающимися их комплексами-урочищами одного или разных типов.
Из сказанного видно, что ландшафтное районирование является прежде всего основой физико-географического исследования. Именно на это обратил внимание Л. Б. Рухин [896], определяя сущность палеогеографии как науки о географических ландшафтах прошлого и их развития. Необходимо при этом подчеркнуть, что если объектами физико-географических исследований служат современные ландшафты, доступные всесторонним наблюдениям, то палеогеография изучает сохранившиеся в ископаемом состоянии фрагментарные, притом часто косвенные показатели, обобщение и анализ которых позволяют воссоздать общие черты ландшафтов геологического прошлого. К тому же захоронения ископаемых остатков организмов, с которыми имеют дело палеобиогеографы, очень разнообразны по характеру и сравнительно редко представляют собой их прижизненные ассоциации - палеобиоценозы. Поэтому для палеобиогеографов, как правило, лишь частично сохраняется перспектива в восстановлении первичного систематического состава комплексов организмов и в выяснении их приспособлений к разным факторам среды - температуре, атмосферным осадкам, пище, субстрату и т. п., в той или иной мере отражающих их ландшафтную приуроченность.
В исследованиях палеоландшафтов важную роль играют фактор геологического времени, а также степень вторичных изменений пород и содержащихся в них организмов. Поэтому получение соответствующей информации из более древних отложений в общем случае сложнее, чем из более молодых.
Методика рассматриваемых исследований очень многообразна и по своей сложности существенно превосходит ранее описанные направления. Согласно К. К. Флерову и др. [410], в самом общем виде она включает:
1) полевые палеоэкологические и тафономические наблюдения, проводимые в процессе изучения захоронений остатков организмов;
2) определение систематической принадлежности собранных ископаемых;
3) морфофункциональный их анализ и выяснение функций отдельных органов; изучение аутэкологии отдельных групп, их приспособлений к перемещению, пользованию разной пищей и т. п.;
4) основывающийся на ранее полученных данных всесторонний, в том числе исторический, анализ синэкологии исследуемых организмов и их связей с физико-географическими элементами палеоландшафтов;
5) установление связи изучаемой флоры и (или) фауны с климатической зоной и определенным типом палеоландшафта;
6) изучение прямыми и косвенными методами географических ареалов видов и вышестоящих таксонов;
7) обобщение всех собранных данных о географическом распространении, факторах эндемизма и возможностях преодоления изученными видами, родами и т. д. миграционных преград;
8) построение карт-схем ландшафтно-географического районирования с показом на них путей расселения видов, их крупных ассоциаций, целых флор и фаун; выяснение характера миграционных процессов - прохореза, аллагии и др. [2371].
Остановимся на наиболее важных общих положениях палеобиогеографии, вытекающих из всего сказанного выше, и прежде всего на вопросе о взаимосвязях рассмотренных путей и направлений районирования.
Кажется бессмысленным противопоставлять друг другу объективно существующие самостоятельные направления исследований. Наоборот, сегодня для изыскания наиболее рациональных решений конкретных теоретических и практических задач необходимо все более глубокое проникновение в сущность и возможности каждого из них и вместе с тем определение дальнейших перспектив создания единого комплексного подхода к разработке проблемы палеобиогеографического районирования.
Несомненно, что палеобиогеографическое районирование, как и биогеографическое, должно основываться на последовательном использовании принципов, соответствующих избранному направлению. Неприемлемым следует считать эклектический подход к выделению одних биохорий на основании одного направления, а других - на основании другого направления, когда возможности логического развития исследований по каждому из этих путей остаются еще неисчерпанными.
Сказанное ни в коей мере не означает, что связи между различными направлениями невозможны и тем более запретны. Ранее мы [606, 607, 649] уже затрагивали этот вопрос, сейчас появилась возможность несколько расширить и углубить его постановку.
Совершенно очевидно, что взаимосвязи направлений, относящихся к одному и тому же пути исследований - биохорологическому или экосистемному, логически гораздо более прочны, чем между направлениями, принадлежащими разным путям. Если первому этапу биохорологического районирования соответствует ареалологическое направление, то следующий этап предусматривает уже использование теснейшим образом сочетающегося с ним флористико-фаунистического направления. Первым этапом экосистемного районирования оказывается биофациальное направление. В дальнейшем оно вместе с климатобиогеографическим сочетается с ландшафтно-географическим направлением [610]. Обращение же в ходе одного исследования к направлениям, относящимся к другому пути районирования, должно быть всегда ясно оговорено и специально обоснованно.
Подобно тому, как в биогеографии происходит сейчас постепенное слияние двух основных путей развития этой области знаний, существующих уже около 200 лет, - биологической географии и географической биологии в теоретическую биогеографию [631], так, по-видимому, и в палеобиогеографии можно ожидать объединения биохорологического и экосистемного путей исследований в теоретическую палеобиогеографию. По аналогии в задачи последней должно входить изучение распространения вымерших растений, животных и их разнообразных сообществ на фоне палеогеографических условий среды, подходя к этому с различных позиций.
В данном случае, утрачивая свою самостоятельность, хорологический и экосистемный пути будут играть роль последовательных этапов в решении указанной выше основной задачи палеобиогеографии.
В каком бы направлении ни шло палеобиогеографическое исследование, оно сталкивается с некоторыми общими проблемами. Главнейшая из них - выявление используемых признаков и определение их веса. Именно к этому сводятся такие обычные вопросы, которые задает себе исследователь, - "на что следует обратить внимание?", "чему надо отдать предпочтение?", "стоит ли учитывать это различие?", "надо ли выделять такую-то биохорию?" и т. п. Как уже было сказано, проблема веса признаков в науке не имеет общего удовлетворительного решения. Тем не менее разработаны некоторые частные способы ее решения, одним из которых служит обращение к цели исследования. Мы классифицируем и расчленяем объекты по-разному, т. е. придаем признакам разные веса, ориентируясь на разные цели. Этот подход не снимает проблемы хотя бы по той причине, что цели сами поддаются классификации и расчленению. Ставя вес признаков в зависимость от разных, т. е. различающихся по каким-то признакам целей, мы лишь переносим проблему веса признаков на иной уровень или в иную область. Правда, и здесь мы можем опереться, как и в случае веса признаков, на прошлый опыт, в который уже вычленены некоторые цели исследования. В случае палеобиогеографии такими целями издавна были разные приложения палеобиогеографических обобщений, в том числе и схем районирования. Это реконструкции климатов, очертаний континентов и акваторий, история литосферных плит, происхождение океанов и т. д. Нетрудно видеть, что упорядочение этого перечня равносильно классификации разных направлений в исследовании или, что то же самое, классификации смежных научных дисциплин.
Таким образом, начав с внутренних проблем палеобиогеографии (нас интересовал вес признаков), мы пришли к значению палеобиогеографии для других областей знаний. Иными словами, палеобиогеография не может решить собственных проблем, не заботясь о проблемах других наук, с которыми она связана. Это, заметим, свойственно всем наукам и отсюда вытекает необходимость единства науки. Поэтому анализ приложения палеобиогеографических данных в смежных дисциплинах имеет первостепенное значение для самой палеобиогеографии.
Поскольку в данной работе имеется специальная глава о практическом значении палеонтологических, в том числе палеобиогеографических исследований (см. 8.1), мы сосредоточим внимание исключительно на взаимосвязях палеобиогеографии со смежными науками.
Зависимость палеобиогеографии от систематики очевидна (см. например, сказанное выше об ареало-генетических исследованиях и соответствии ранга биохорий и рангов эндемичных таксонов). Однако и систематика широко опирается на палеобиогеографические данные. В традициях филогенетической систематики таксоны организмов признаются историко-генетическими совокупностями, поэтому нарушение географической целостности этих совокупностей служит предостережением систематику и заставляет его проверить, не объединил ли он в одном таксоне конвергентные (гомеоморфные) формы.
Общность географического распространения паратаксонов, выделяемых по дисперсным частям, может указывать на прижизненное сочетание частей, вести к реконструкции целых организмов и далее выделению вместо паратаксонов полноценных таксонов. Географическая определенность рассматривается как один из важнейших критериев вида как такового [376], тем более в палеонтологии, где сонахождение во многом заменяет критерий скрещиваемости. Именно различие в распространении служит главным критерием, отличающим подвид от других внутривидовых категорий. Ясно, что взвешивание признаков таксонов путем обращения к палеобиогеографии легко может быть извращено, если разная географическая приуроченность выступает как единственное основание для разделения таксонов. Уже из приведенных примеров видно, что связь палеобиогеографии с таксономией обогащает обе дисциплины.
О связи палеоклиматологии и палеобиогеографии уже говорилось. С одной стороны, не обращаясь к палеоклиматам, мы не можем понять очень многого в распределении и изменении биохорий. С другой - без обращения к палеобиохориям невозможно реконструировать палеоклиматические зоны. Даже при изучении современных климатов приходится обращаться к распределению организмов, так как оно отражает климатические параметры, усредненные за многие годы, и позволяет экстраполировать климатические наблюдения в пространстве. Не случайно современные карты климатической зональности во многом совпадают с зонами растительности, а для обозначения типов климатов используются названия растений (например, климат березы или климат фуксии Б классификации Кеппена). В. А. Красилов [520] рекомендовал использовать этот подход и в палеоклиматологии; об этом пишут и другие авторы [158, 656].
Палеобиогеография соприкасается с тектоникой особенно-тесно, когда обсуждается проблема мобилизма материков, о чем неоднократно писалось в литературе [524, 801, 2032]. Если раньше географическое распределение организмов рассматривали в связи с их расселением из центров происхождения, то в настоящее время его все чаще связывают с движением материков. При этом различаются [1697] три типа изменений географической дифференциации фаун и флор во времени: 1) увеличение сходства между ними по мере сближения материков, 2) уменьшение сходства по мере расхождения материков и 3) увеличение сходства у одних при одновременном его уменьшении у других фаун (флор). Следует, однако, заметить, что при обсуждении этой проблемы часто смешиваются два вопроса: былое соединение материков (или акваторий) и перемещение материков. Нередко устанавливается палеобиогеографическая общность материков и это считается достаточным для утверждения об их смещении. В действительности же это свидетельствует лишь о том, что биохория объединяла материки до того, как возникла разделяющая их акватория, которая могла образоваться и при устойчивом положении материков [524]. О перемещениях же материков мы можем судить только тогда, когда известна некоторая заведомо устойчивая координатная сетка, по отношению к которой материки смещаются в разных направлениях. В роли такой координатной сетки до сих пор выступали лишь палеомагнитные широты и палеоклиматические границы, причем такие, которые нельзя связать с перемещением полюсов и местными палеогеографическими факторами. Пока в числе таких границ могут выступать лишь границы областей с морозным и безморозным климатом и именно они однозначно указывают на взаимное смещение материков [641].
В некоторых случаях на горизонтальные перемещения литосферных плит может указывать и сам характер палеобиогеографических границ, а именно: их экстраординарная резкость, отсутствие экотонных зон. Таковы, например, палеобиогеографические границы, проходящие вдоль Тетиса и ограничивающие позднепалеозойские зоо- и фитохории [356, 357, 656 и др.]. Ясно, что не все резкие границы имеют тектоническое происхождение, о котором можно заключить лишь в результате комплексного изучения районов, в которых проходят эти границы.
Давая важный материал для геотектонических реконструкций, палеобиогеография сама может опираться на них. После того как широким комплексом методов было установлено раздвижение материков, расположенных сейчас по обе стороны Атлантического океана, палеобиогеография в своих дальнейших построениях основывается на мобилистских реконструкциях, хотя бы и гипотетических. Так, недавно было показано, что древнейшие покрытосеменные раньше всего и в наибольшем количестве появились на западе нынешней Экваториальной Африки [1277]. Здесь намечается центр происхождения и расселения покрытосеменных, т. е. группы, определившей всю последующую эволюцию наземной биоты и лежащую в основе общего палеобиогеографического районирования суши, начиная с позднего мела. Ясно, что знать размеры и положение центра происхождения такой группы очень важно. Мобилистские реконструкции указывают на соединение Африки и Южной Америки в начале мелового периода, так что есть все основания экстраполировать наблюдения в Африке на Южную Америку, а точнее, на ту ее часть, которая, согласно мобилистским реконструкциям, примыкала к Экваториальной Африке. И действительно, в Бразилии и Аргентине мы также сталкиваемся с очень ранним появлением пыльцы покрытосеменных. Более того, прослеживание расселения покрытосеменных по палинологические данным хорошо согласуется с предполагаемыми эпизодами дрейфа. Палеофитогеографические и тектонические данные, взятые во взаимосвязи, дополняют друг друга и приводят к более полной и убедительной картине.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'