
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Эмпирические закономерности (правила) эволюции
Правило направленности эволюции (ортогенеза). Пока действует заключенная в структуре система запретов, вся динамика развивающейся системы будет обладать свойством гомеорезиса - направленного канализованного развития [1079]. Развитие не может быть абсолютно ненаправленным.
Между двумя состояниями системы, предыдущим и последующим, можно указать вектор изменения. Такая направленность касается только данного акта выбора и в общем случае не требует памяти. Указание направления каждого акта выбора не означает направленного развития. Последнее возникает там, где неслучайные процессы охватывают не менее трех преемственно связанных состояний (предшествующее операнду состояние - операнд - выбор), обозначенных нами как элементарное звено развития. В этом случае состояние, предшествующее операнду, может влиять на выбор только благодаря памяти.
Оператор непосредственно может влиять только на исход данной операции выбора, причем единственным образом: ограничивая пространство выбора, заданное структурой операнда. Следовательно, направление развития в рамках данной операции выбора определяется исключительно внутренними свойствами развивающейся системы - операнда. Внешняя среда - оператор - может влиять не только на направление данного, но и последующих выборов, направлять развитие только в меру его отображения в структуре системы - запоминания. Таким образом, роль среды в определении направления развития, как отмечал Ч. Дарвин [310], исключительно косвенна. Характер и адекватность отображаемого аспекта среды определяется свойствами развивающейся системы. Опосредование оператора операндом в процессе его отображения всегда глубже, чем опосредование операнда оператором. В противном случае среда реорганизует систему по своему образу и подобию, что равносильно потере индивидуальности и гибели системы. Все рассуждения о прямом влиянии среды на направление биологического развития, а тем более определяющей ее роли, основаны на отсутствии полного разграничения направленности отдельного выбора и направленности развития.
Правило адаптивной направленности развития (адаптиогенеза). Приспособление в биологии часто определяется как ансамбль свойств организации, обеспечивающих устойчивое поддержание в заданных пределах ее количественных и качественных параметров. И это явление можно описать при помощи понятия "устойчивость". Адаптация - понятие, призванное оттенить еще один аспект структуры - конкретные способы поддержания ее устойчивости. Поскольку для структуры альтернатива устойчивости - небытие, всякое длительное развитие должно протекать с явным преобладанием специализации над эманацией, запоминания над забвением, преемственности над новизной и адаптации над процессами ее разрушения. Адаптация - необходимый аспект любой специализации, а ее разрушение - необходимое следствие любой эманации. Это правило накладывает дополнительное граничное условие на предыдущее.
Жизнь, как всякая неравновесная диссипативная структура, поддерживающая свою устойчивость благодаря потреблению внешней энергии и (или) вещества, немыслима без постоянного обмена со средой. Отсюда определение адаптации как соответствия- адекватности структуры системы и среды. Эта адекватность означает наличие структуры отношений системы к среде - умвельта [2403], обеспечивающего поддержание устойчивости системы в среде. Адекватность умвельта важным для жизнедеятельности системы факторам среды и есть адаптация. Можно говорить не только о внешнем, но и внутреннем умвельте - структуре отношений целого к своим элементам. Потеря адекватности внутреннего умвельта фактической структуре элементов равносильна потере целостности.
Возможность активной адаптации для диссипативных, в том числе живых структур, т. е. отыскание ими устойчивых стационарных состояний (приобретение необходимой адекватности умвельтов) вдали от состояния термодинамического равновесия, непосредственно вытекает из достижений неравновесной термодинамики и синэргетики [747, 1116, 1204, 1210].
Адаптация - целостное свойство, аспект структуры. Поэтому из адаптивной стратегии развития целого не следует, что каждая изолированная его часть или аспект должны нести вполне определенную собственную адаптивную функцию. Никакое адаптивное толкование конечного аспекта структуры (указание его роли Б поддержании устойчивости целого) не будет полным в меру сопряженности разных аспектов целого и их функций.
Правило постепенности эволюции (градуализма). Постепенное преобразование органических форм в палеонтологических рядах, наличие переходных форм и постепенная трансформация видов из одного в другой в последовательных биозонах были известны еще додарвиновской палеонтологии [291]. Эти факты находились в согласии с представлением об эволюции путем накопления мелких наследственных различий, поэтому Ч. Дарвин рассматривал их как косвенные доводы в пользу своей теории.
Были указаны три граничных условия, при соблюдении которых правило должно выполняться: 1) последовательные биозоны не разделены перерывами в осадконакоплении; 2) в верхней, более молодой, биозоне не происходит замещения близким видом, мигрировавшим из другой географической области; 3) остатки организмов присутствуют в достаточном количестве на протяжении всей осадочной толщи. Иными словами, правило выполняется при условии полноты стратиграфической и адекватности палеонтологической летописи, что почти никогда невозможно утверждать. Эмпирическое обоснование правила на массовом материале летописи практически невозможно. Однако немногие специально подобранные случаи сравнительно полных стратиграфических и палеонтологических последовательностей с возможно более узкой пространственной локализацией филогенетических процессов позволяют обнаружить тенденцию ископаемых организмов к рядоположенности и гомотаксису с их стратиграфическим положением [250, 737, 738, 1017]. Эти "кинематографические" тенденции [77] иногда рассматривают как прямое указание палеонтологии на факт существования эволюции [1137, 1194], хотя речь идет о весьма правдоподобной гипотезе [654, 664].
Постепенность эволюции хорошо согласуется с представлением о разрушительном действии на структуру больших порций кратковременно получаемой новизны. Любые формы макрогенеза сопряжены с большим риском вымирания. Это подтверждает и правило обратной зависимости ранга таксона и частоты возникновения таких таксонов в филогенезе. Постепенное и преемственное развитие, сохраняющее архетип таксона и направление его специализации, в соответствии с правилом "перемежающегося равновесия" составляет основной фон филогенетических событий. Градуализм нашел отражение в правиле градации [559] или постепенного прогрессивного усовершенствования [1130, 1382], правиле постепенного количественного нарастания разнообразия форм жизни от более древних отложений к более молодым [1256]. Правило возникновения филогенетических новшеств преимущественно на поздних стадиях онтогенеза [926, 928, 1178, 1776] - основа "биогенетического закона" Э. Геккеля [220] и закона К. Э. Бэра [129, 928, 1178] Терминальные изменения касаются более мозаичной стадии онтогенеза, в течение которой нарушение одной части меньше вредит развитию других частей целого [67]. Напротив, даже небольшое, но не регулируемое к норме уклонение ранних стадий развития влечет вследствие активного взаимодействия частей на стадиях детерминации цепную реакцию нарушения последующего хода индивидуального развития
Таким образом, правило постепенности и непрерывности эволюции - просто внешнее выражение преемственности, необходимое условие развития, и оно должно выполнять эвристическую функцию чем больше кратковременно получаемой новизны требует филогенетическая гипотеза, чем большего размера скачка требуют ее условия, тем менее при прочих равных условиях эта гипотеза правдоподобна.
Правило редкости переходных форм в палеонтологической летописи. Согласно Ч. Дарвину, это правило призвано объяснить несогласие массовых фактов палеонтологической летописи с правилом постепенности эволюции, которое в палеонтологии базируется на выборочных примерах. Чрезмерное увлечение значительными успехами полевой палеонтологии привело некоторых исследователей к выводу, что летопись стала в основном адекватной филогенезу, а наличие разрывов в палеонтологических рядах свидетельствует в пользу макрогенеза [433]. От столь оптимистических выводов удерживают факты отсутствия или крайней редкости ряда таксонов в ископаемом состоянии на протяжении всего или части их филогенеза, необходимость прибегать к понятию "практически вымершего" таксона [367, 368], а также требование преемственности, известной постепенности, филогенеза.
Факт избирательного выпадения именно переходных форм потому и питает идеи макрогенеза, что не может быть объяснен стратиграфической и тафономической неполнотой летописи как таковой. Если существует проблема переходных форм, то, значит, должны быть и продолжительные периоды относительного стасиса таксонов, когда архетип их сохраняется. Для такой фазы филогенеза неоднократно предлагались названия (рис. 10). О. Шиндевольф [2223, 2224] назвал ее типостазом. Чередование длительных периодов стасиса таксонов с относительно краткими периодами дестабилизации [314] является ядром правила "перемежающегося равновесия" [1034, 1550, 1666, 2318] Вероятность обнаружения группы в палеонтологической летописи, при прочих равных условиях, прямо пропорциональна всем основным критериям биологического прогресса А. Н. Северцова [928]: фактическому таксономическому разнообразию и численности группы, широте ее географического и стратиграфического распространения [457]. Неполнота летописи будет объяснять избирательное выпадение переходных форм, если они приходятся на фазы относительного биологического регресса и сравнительно скоротечны. Об этом говорит даже название "типолиз", данное этой фазе О. Шиндевольфом [2223, 2224]. На эти плохо представленные в летописи моменты филогенеза попадают начала важнейших морфологических перестроек, определяющие становление новых таксонов и смены направлений специализации (типогенез по О. Шиндевольфу). Позиции градуалистов и сторонников прерывистой эволюции отличаются лишь интерпретацией факта редкости переходных форм и, как можно видеть, в чем-то права каждая из сторон.
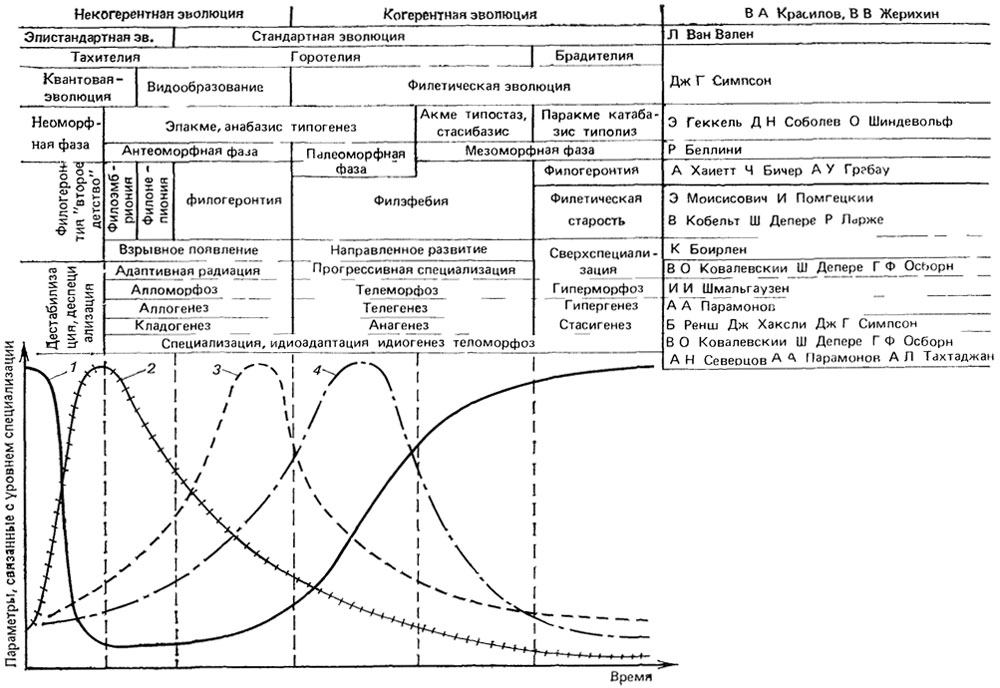
Рис. 10. Основные фазы полного филогенетического цикла (типичная смена форм адаптиогенеза), их свойства и классификация по разным авторам. Составил А. С. Раутиан. 1 - степень специализации устойчивости преемственности адаптации со стояний развивающейся системы и направленности ее развития снижение свободы выбора (изменчивости) и творчества, 2 - темп и уровень эманации (дестабилизации вымирания) снижение устойчивости преемственности адаптации развивающейся системы и направленности ее развития, 3 - темп диверсификации дивергентной эволюции (адаптивной радиации), 4 - темп специализации (филетической эволюции) и рост целостности
Правило "перемежающегося равновесия" (пунктуализма). Сформулированное С. Гоулдом, Н. Элдреджем [1550, 1666] и С. Стенли [2318], это правило гласит: в эволюции видов, как правило, чередуются длительные периоды стасиса, когда основные структурные черты вида сохраняются неизменными, и короткие периоды трансформации одной видовой формы в другую. В основу правила положен не столько массовый материал летописи, хотя он использован тоже, сколько ограниченное число интенсивно исследованных случаев видообразования в стратиграфически и палеонтологически непрерывных или почти непрерывных разрезах. Временные масштабы разрывов оценивались абсолютными методами датирования. С. Гоулд полагает, что этой закономерности подчиняется 95% случаев видообразования, а С. Стенли оценивает различия в длительности периодов стасиса и трансформации в несколько порядков, измеряя ее предполагаемым числом прошедших поколений [1034]. Авторы связывают свои воззрения с идеей Э. Майра [594, 596] о "бутылочном горлышке" низкой численности, которое проходит популяция в процессе видообразования.
Пунктуализм хорошо согласуется с теорией. Пока система существует, обладает индивидуальностью и выделенностью в среде, ее структура сохраняет большую и меньшую устойчивость (гомеостаз) по отношению к возмущениям. Следовательно, даже непрерывный градиент внешнего воздействия или внутреннего возмущения, по отношению к которому система организована (обладает умвельтом), вызовет более или менее дискретную реакцию системы по мере преодоления возмущающим фактором порога устойчивости [1193, 1194, 1196]. Краткие неустойчивые фазы эманации ассоциируются с переходными формами. Пунктуализм не противоречит постепенности филогенеза, настаивая лишь на значительной и закономерной неравномерности темпа эволюции [1034]. Это правило постулирует скоротечность и неустойчивость переходных форм и широкое распространение в филогенезе фаз относительной устойчивости архетипа таксона и направления его специализации.
Не всякая сальтация несет большой заряд новизны с ее разрушительными последствиями. Довольно резкие филогенетические перестройки могут быть связаны с неотенией и фетализацией [889, 1178, 1181], с переносом признаков с одного пола на другой [1193]. В этих случаях новое филогенетическое состояние имеет солидную исторически подготовленную базу, что в случае полноты материала легко опознать морфологически.
Правило "перемежающегося равновесия" было отчасти предвосхищено правилом происхождения нового наследственного признака В. Е. Руженцева, теоретически обоснованного М. А. Шишкиным [1178, 1180, 1181]. Это правило гласит: всякое новообразование в момент своего появления неопределенно и лишь пройдя испытание отбором, оно либо исчезает, либо становится полезным; изменчивость, отрицая наследственность, только при участии отбора сама становится наследственностью [887]. Формулировка В. Е. Руженцева указывает на исходную неустойчивость нового признака (а значит, до известной степени и его обладателя) и связь устойчивости с адаптацией. На значение и приоритет правила В. Е. Руженцева недавно указал Л. Ван Вален [2424].
Правило дестабилизации (эманации). Это правило сформулировал В. В. Жерихин [314], базируясь прежде всего на экспериментах Г. X. Шапошникова [1147-1150, 2282] по детальному анализу смены адаптивных норм. Понятие "адаптивная норма" ввел И. И. Шмальгаузен [1184, 1192]. Его можно определить как ансамбль жизненных циклов, онтогенезов или их стадий, возникающих как исторически оправданный, устойчивый (и в этом смысле - адаптивный и наследственный) ответ на воздействия исторически типичной внутренней и внешней среды [514].
В опытах Г. X. Шапошникова по эволюции тлей в условиях: смены кормового растения наблюдалась резкая дестабилизация многих морфологических признаков на фоне резко возросшего уровня элиминации, достигавшего в отдельные моменты опыта 80-90% численности популяции. За фазой дестабилизации последовала фаза стабилизации новой адаптивной нормы, сопровождавшаяся адаптацией к новому кормовому растению и понижением уровня элиминации до уровня контроля. Новая популяция имела морфологическую характеристику хорошего вида и практически полную репродуктивную изоляцию от контроля, т. е. предковой популяции. Смена адаптивной нормы, схематически изображенная на рис. 11, заняла 15 поколений тлей. Некоторые из результатов опыта недавно воспроизвел В. Б. Сапунов [912]; проведено еще несколько аналогичных экспериментов (см. 7.4).
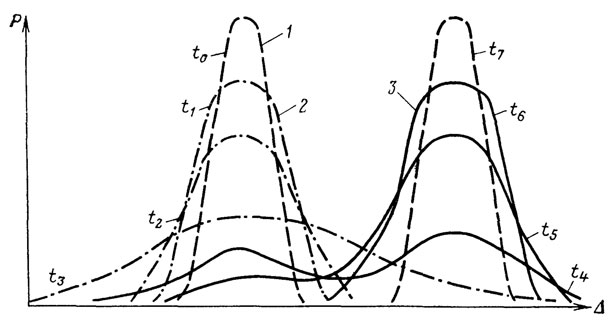
Рис. 11. Схема смены адаптивных норм в остром опыте Г. Х. Шапошникова. Составил А. С. Раутиан. 1 - старая и новая устойчивые адаптивные нормы; 2 - стадии эманации (дестабилизации) старой адаптивной нормы; 3 - стадии стабилизации (специализации) новой адаптивной нормы; t0-t7 - последовательные моменты времени; по абсциссе отложена величина (количественный показатель) признака (Δ); по ординате - частота соответствующих количественных значений величины признака (P)
Смена адаптивной нормы оказалась дискретной для популяции (частоты признаков) и градуальной для морфологических рядов переходных форм, преемственно связавших старую и новую нормы. Плавное смещение популяционной кривой изменчивости в область исторически нетипичных значений признаков (в духе классической схемы движущего отбора - рис. 12, а) должно вызвать дестабилизацию популяции. Это и наблюдалось в опытах Д. К. Беляева [69] и сотрудников по искусственному движущему отбору (рис. 12, б). Градуальная смена адаптивной нормы популяции противоречит исторической обусловленности устойчивости ее адаптивной нормы. Снижение эффективности стабилизирующего отбора прежней нормы привело к ее дестабилизации. Снятие популяционных и некоторых организменных ограничений прежней нормы привело к мобилизации резерва изменчивости [1192], т. е. к повышению свободы выбора. Возросшая свобода творчества в сочетании с жестким стабилизирующим отбором устойчивых в новых условиях онтогенезов формируют новую адаптивную норму.
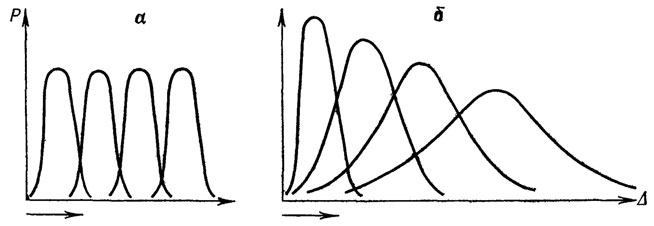
Рис. 12. Две версии движущей формы отбора: а - классическая схема (по Н. В. Тимофееву-Ресовскому, Н. Н. Воронцову и А. В. Яблокову); б - схема движущей формы искусственного отбора в острых опытах Д. К. Беляева и сотрудников: наблюдается дестабилизирующий эффект движущей формы отбора (составил А. С. Раутиан по данным Д. К. Беляева). Стрелкой показано направление вектора отбора и времени
Эффективность движущего отбора в дестабилизированной популяции должна быть исчезающе мала: 1) дестабилизация понижает наследуемость признаков и эффективность отбора по ним; 2) эффективный отбор признаков с низкой наследуемостью требует длительного времени [577], а затяжная дестабилизация - верная гибель; 3) чем сильнее движущий отбор смещает популяцию за пределы адаптивной нормы, тем слабее его направляющая роль. Область поисков новой адаптивной нормы ограничивается структурой обратимо мобилизуемого резерва изменчивости, определяющейся, в свою очередь, структурой прежней нормы, а не эффективностью движущего отбора.
Палеонтология не осталась в стороне от исследования дестабилизации, особенно трудного для нее по причине редкости переходных форм. Одно из филогенетических правил Э. Д. Копа [1457] гласит: перед моментом вымирания группа, как правило, повышает свое таксономическое разнообразие. Речь идет, очевидно, о неоднократных попытках вымирающей группы выйти из экологического кризиса путем смены направления специализации (немыслимого без дестабилизации). Но все попытки кончались лишь формированием короткоживущих таксонов. Это правило связывает процессы дестабилизации как с вымиранием, так и с типогенезом. Правило возникновения нового наследственного признака В. Е. Руженцева, концепции квантовой эволюции [952, 1150, 2282] и пунктуализма явились важными шагами в понимании чередования фаз специализации и эманации, а также неустойчивости и скоротечности последней. Наконец, само правила дестабилизации сформулировал палеонтолог В. В. Жерихин. На ископаемом материале продемонстрированы дестабилизации целых экосистем и связанные с ними волны вымирания и появления новых групп [367, 368, 457].
Правило прогрессивной специализации. Если в процессе развития устойчивость и защищенность системы от внешних и внутренних возмущений растет, т. е. идет процесс прогрессивного приспособления в избранном направлении, то мы встречаемся со специализацией [313]. Говорить о постоянстве направления специализации можно только применительно к избранному уровню рассмотрения, поскольку всякое изменение состояния эволюирующей системы требует известного нарушения устойчивости - дестабилизации, а следовательно, и нарушения направленности. Иерархия корреляционных систем организма и их координированных изменений в филогенезе [1192, 1196, 1197] - та биологическая реальность, которая стоит за этими уровнями рассмотрения, отраженными, в частности, в таксономической иерархии. Низшим уровнем специализации будет процесс стабилизации фенотипов определенного класса.
Прогрессивная, длительное время не прерывающаяся на данном уровне рассмотрения, специализация, сохраняющая архетип таксона соответствующего ранга и направление адаптивного совершенствования, является воплощением постепенного преемственного и наиболее безопасного в отношении вымирания развития, составляющего основной фон филогенетических событий. Напротив, резкое возмущение системы, крупное приращение структурной новизны ослабляет преемственность, форсирует забвение прежнего адаптивного опыта, нарушает устойчивость канала развития - филокреода [647], меняя направление специализации.
Особенно ярко прогрессивная специализация проявляется в явлениях параллелизма, когда независимые потомки общего предка сохраняют тенденцию приобретения гомологичных признаков и целых функциональных аппаратов. Эта тенденция нашла отражение в явлениях гомоплазии (Дж. Рей Ланкастер), понятиях латентной, потенциальной (Г. Осборн), косвенной или параллельной (В. Е. Руженцев) гомологии [291, 887]. Об этом же говорят правила гомологических и гетерологических рядов Э. Копа [1457], схематически показанных на рис. 13-15. Явления параллельной и итеративной эволюции, в частности правило параллельного достижения разными родственными филетическими линиями приблизительно одного уровня организации [1033, 1457, 2287], нашли отражение в представлении о вертикальной и горизонтальной классификации [883, 1033]. Эти эмпирические обобщения свидетельствуют о большой роли начальных условий развития и преемственности, ведущих к проявлению одних и тех же латентных потенций в параллельных линиях специализации [74, 592, 975]. Вероятный механизм действия начальных условий был рассмотрен Н. Н. Иорданским [444, 445] и будет описан при характеристике правила ключевого ароморфоза.
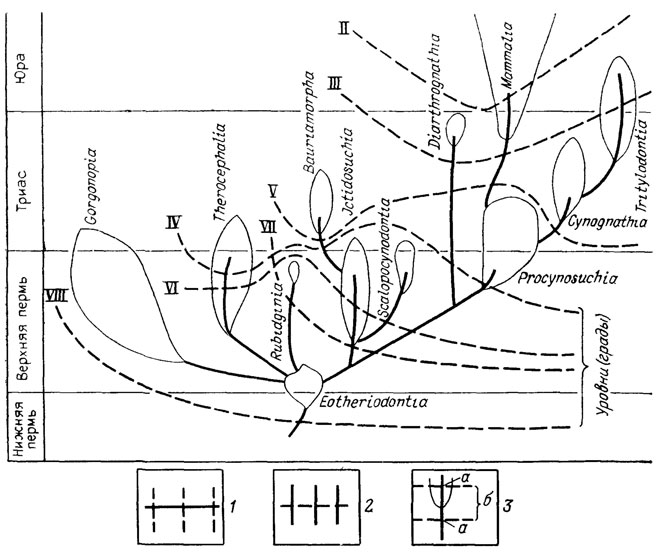
Рис. 13. Аристогенез в эволюции териодонтов. По Л. П. Татаринову. Кривые, пересекающие филетические линии, соответствуют уровням (градам), на которых приобретались характерные для млекопитающих признаки; II - звукопроводящий аппарат из трех слуховых косточек, III - челюстное сочленение между зубной и чешуйчатой костями, IV - мягкие, снабженные собственной мускулатурой губы, V - несколько расширенные большие полушария головного мозга, VI - зона усиленного кровоснабжения на губах, связанная, по-видимому, с формированием зачаточных вибрисс, VII - трехбугорчатые щечные зубы, VIII - верхние обонятельные раковины; ряды Э. Д. Копа; 1 - гомологические, 2 - гетерологические; 3 - сопряжения филетических линий с уровнями (градами): а - мутации В. Ваагена, ректиградации или аристогены Г. Ф. Осборна, б - аллойометроны Г. Ф. Осборна
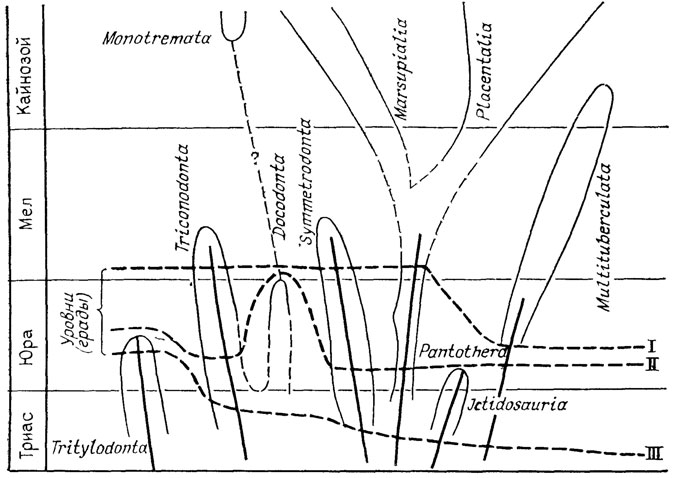
Рис. 14. Аристогенез в эволюции мезозойских млекопитающих. По Дж. Симпсону. I - телэнцефалический мозг; остальные усл. обозначения см. на рис. 13
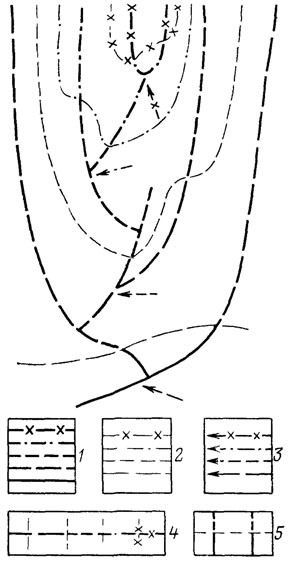
Рис. 15. Схема филогенеза в соответствии с концепцией аристогенеза. Составил А. С. Раутиан. 1 - филетические линии; 2 - границы уровней организации; 3 - моменты приобретения ключевых ароморфозов вышестоящих уровней организации; ряды Э. Д. Копа: 4 - гомологические; 5 - гетерологические
Можно указать направления специализации, характерные для всех стволов животного царства, удовлетворяющих правилу прогрессивного совершенствования [1130, 1382]. Эти направления отражены в следующих правилах: прогрессивной дифференциации, интеграции и разделения функций [1192, 1196, 1382]; автономизации и рационализации онтогенеза [1192, 1196, 1197]; цефализации - увеличения относительных размеров и концентрации центральной нервной системы [1919]; возрастания активности и чувствительности [250].
Сравним особенности прогрессивной специализации и дестабилизации (эманации) как основных модусов эволюции:
| Прогрессивная специализация | Дестабилизация (эманация) |
|---|---|
| Соотношение запоминания и забвения | |
| Запоминание превосходит забвение | Забвение превосходит запоминание |
| Надежность запоминания | |
| Возрастет | Понижается |
| Объем памяти | |
| Возрастает | Понижается |
| Преемственность (надежность наследования, наследуемость) | |
| Возрастает | Понижается |
| Устойчивость | |
| Возрастает | Понижается |
| Свобода выбора (изменчивости) | |
| Понижается | Возрастает |
| Типичная длительность | |
| Продолжительна | Очень коротка |
| Корреляции в онтогенезе | |
| Возникают | Разрушаются |
| Таксономический уровень изменчивости признаков | |
| Понижается | Возрастает |
| Степень приспособленности (адаптированности) | |
| Возрастает | Понижается |
| Направленность эволюции | |
| Возрастает | Понижается |
| Предел развития (эволюции) в соответствии с данным модусом | |
| Персистирование, или переход к дестабилизации (эманации) | Разрушение структуры (вымирание), или переход к специализации |
Третий из качественных режимов (модусов) развития (см. рис. 9), как отмечалось, является переходным и наблюдается в момент смены одного из двух основных модусов эволюции другим.
Онтогенез как прогрессивная специализация. Ярчайшим проявлением параллельной и итеративной специализации является повторяемость онтогенетических процессов - индивидуальной специализации организмов каждого вида. В высшем проявлении онтогенез многоклеточных абсолютно необратим и сопровождается программируемой смертью. У простейших онтогенез обратим, а особь потенциально бессмертна. Строго не ограниченные определенной стадией жизненного цикла процессы генетического обмена у прокариот принципиально допускают существование прокариотического бионта, развивающегося в разные моменты жизни на базе разных геномов. По этому важнейшему признаку у них оказывается неопределенной грань между индивидуальным и историческим развитием. Перечисленные типы организации (прокариоты - простейшие - многоклеточные) могут рассматриваться как узловые стадии разделения процессов онто- и филогенеза (рис. 16). Биологический смысл их разделения заключается в снятии противоречия между высокой организацией, требующей для своего осуществления высокого уровня устойчивости и автономности [1192, 1196], и требованием высокой лабильности, свободы творчества для продолжения развития. Снятие это осуществлено путем разделения проявлений жизни на популяционный и организменный уровни организации.
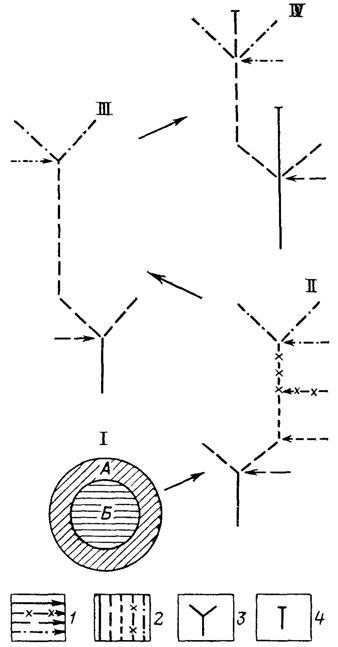
Рис. 16. Узловые стадии становления типичного онтогенеза метазоа и мета-фита как процесса тотальной специализации. Составил А. С. Раутиан. I - гипотетическая стадия разделения сомы (А) и гермы (Б); схемы изображения онтогенезов: II - прокариотического организма; III - эукариотического простейшего; IV - метазоа и метафита; 1 - вставки чуждой гермы в процессы онтогенеза; 2 - фазы онтогенеза, осуществляющиеся без вмешательства новой гермы; 3 - размножение; 4 - смерть
Популяция - низко организованная система с относительно слабыми связями между гомономно построенными, автономными и взаимозаменимыми элементами (особями), способная существовать продолжительное время, но медленно и обратимо специализирующаяся (обучающаяся), оказывается способной к эволюции. Организм - высокоорганизованная система с весьма сильными связями между гетерономно построенными, уникальными (незаменимыми), но подчиненными целому элементами (органеллами, органами, тканями), имеющая ограниченный срок существования вследствие совершенства памяти и необратимости процессов быстрого обучения (специализации). Сопряжение между обоими уровнями осуществляется благодаря процессу размножения, а обратная связь - благодаря процессам отбора (выбора) [1192, 1193, 1196]. Цена снятия отмеченного противоречия - регулярная эманация индивидуальной организации и опыта в процессах размножения, а в своих высших формах - смерть.
Правило специализации экто- и эндосоматических подсистем организма в филогенезе. Адаптация как выражение прогрессивной специализации носит характер прогрессивной оптимизации целого. Пути этой оптимизации сталкиваются с ограничением: оптимизация целостной системы по каждому параметру осуществляется в ущерб оптимизации по любому другому параметру в меру их взаимной сопряженности [754, 859, 1223]. Поэтому процесс специализации осуществляется тем легче и быстрее, чем меньшее число противоречивых структурно-функциональных подсистем в него вовлечено. Действительно, обычно процесс специализации удается особенно тщательно проследить на примере отдельных функциональных подсистем [19-21], например опорно-двигательного аппарата [215, 495, 1023].
Число главных функций, на которые делается адаптивная ставка, всегда невелико. Все прочие эволюируют в координационном порядке, обслуживая в конечном счете оптимизацию главных. Следуя духу, но не букве определений А. Н. Северцова [926], главные структурно-функциональные подсистемы целого можно назвать эктосоматическими, а подчиненные - эндосоматическими. Соответственно семофилезы (филогенетические изменения части или аспекта целого) первых можно назвать проталлаксами, а вторых - дейталлаксами.
Сравним особенности двух основных классов функциональных систем организма, отличающихся по характеру семофилеза в процессе прогрессивной специализации и роли в обеспечении адаптации целого организма:
| Эктосоматические системы | Эндосоматические системы |
|---|---|
| Типы семофилезов, осуществляющихся в процессе специализации | |
| Проталлаксы | Дейталлаксы |
| Количество в организме | |
| Малое | Очень большое |
| Роль в процессе адаптации к внешней среде | |
| Возрастающая | Понижающаяся |
| Роль в процессе коадаптации частей целого | |
| Понижающаяся | Возрастающая |
| Вклад в достигнутую приспособленность организма | |
| Преимущественно - устойчивость к внешней среде | Преимущественно - координация частей и поддержание целостности |
| Внутренняя дифференцировка функциональной системы | |
| Прогрессивная | Ограниченная, распространены редукции |
| Семофилез отношений к организму, как части к целому | |
| Прогрессивная эмансипация в рамках целого организма | Прогрессивное подчинение целому организму |
| Роль в стратегии эволюции организма | |
| Определяющая | Компенсаторная |
| Роль в поддержании устойчивости | |
| Поддержание направления специализации - гомеорезис филогенеза | Поддержание каждого состояния в их последовательном ряду - гомеостазис таксона |
Наиболее характерные формы специализация принимает, когда значительный и долгосрочный прогресс устойчивости (адаптации) достигается при участии минимума эктосоматических подсистем. При этом прогресс будет достигаться при минимуме новизны и опасности вымирания, а направление специализации будет в максимальной степени сохраняться.
Эктосоматические подсистемы по мере их прогрессивной специализации все в возрастающей степени берут на себя функции поддержания устойчивости целого во внешней среде. Это сопровождается дифференцировкой, ростом индивидуальности и эмансипированности в рамках целого. Специализация эктосоматических подсистем достигается ценой прогрессивного подчинения им специализации эндосоматических подсистем и ограничения возможностей независимой от них специализации последних. Такой путь ведет к нарушению целостности (понижению адекватности внутреннего умвельта), причем, как ни странно, в результате развития, направленного на достижение все большей устойчивости целого. Относительно бесконфликтный, направленный и безопасный путь специализации ведет к накоплению внутренних противоречий устойчивости (достигнутого уровня адаптации) и лабильности (свободы выбора и перспективы адаптиогенеза). Эти противоречия сначала останавливают, а затем и разрушают данное направление специализации. Основные фазы процесса прогрессивной специализации приведены на рис. 10. Пример такого процесса, исследованного Т. Уэстоллом [2482] для двоякодышащих рыб, показан на рис. 17.
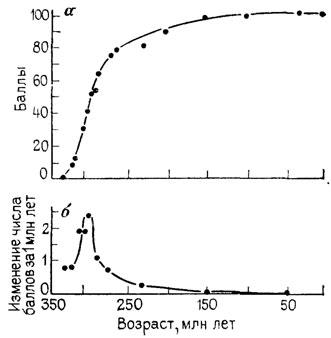
Рис. 17. Специализация и темп аристогенеза двоякодышащих рыб от силура (по реконструкции) до наших дней. По Т. С. Уэстоллу. а - уровень специализации - степень обновленности признаков (преобразования в направлении их современных модальностей), выраженная в условных средних баллах; б - темп аристогенеза - скорость обновления признаков, осуществлявшегося в нескольких параллельно развивавшихся филетических линиях
Правило адаптивной редукции. Оптимизация (адаптация) целого возможна не только за счет дифференцировки и усложнения структуры, при структурном превосходстве системы над средой, но и за счет упрощения структуры - сокращения затрат на поддержание частей (аспектов), функциональное значение которых в поддержании устойчивости не оправдывает себя. Возрастающая роль эктосоматических подсистем по мере специализации понижает относительную адаптивную ценность многих эндосоматических подсистем, для которых и характерны явления адаптивной редукции. В условиях специализации морфофизиологический прогресс протекает параллельно с редукцией; эти процессы как минимум компенсируют друг друга (телогенез) или даже редукция опережает морфофизиологический процесс (катагенез, гипогенез, рационализация онтогенеза [1197]).
Разнообразные формы вторичного упрощения и односторонний морфофизиологический прогресс немногих эктосоматических подсистем (наравне с эволюционными тупиками и вымиранием)- весьма характерные спутники специализации [805, 935, 1197]. Поэтому указания на роль специализации в процессах ароморфной эволюции появились лишь в самое последнее время [933-936], хотя, как мы уже знаем, общий рост уровня организации в конечном счете должен достигаться именно вследствие процессов специализации.
Правило мозаичной эволюции. Данное правило свидетельствует о широком распространении в филогенезе неравномерностей темпов специализации разных структурно-функциональных подсистем целого [594, 935, 1033, 1040]. Статическим его аналогом является правило гетеробатмии - разной эволюционной продвинутости разных частей целого [935, 936, 1040], а онтогенетическим - правило исторического гетерогенезиса стадий индивидуального развития [540, 887].
Эти правила обычно интерпретируются как свидетельство значительной свободы и независимости поведения отдельных признаков в филогенезе [594,596], принадлежащих разным координационным цепям [935, 936, 1040], низкой целостности филогенетических систем [2046, 2048, 2053], а следовательно, низкой преемственности их состояний как целого. В пользу такой позиции говорит лишь широкое распространение и наглядность явлений мозаичности при становлении ароморфных групп высокого ранга [1033], которому должна предшествовать глубокая эманация, а значит, и понижение целостности. Однако само становление есть выражение специализации - повышения устойчивости целого и взаимозависимости его частей. Эволюционная разноуровневость подсистем целого при гетеробатмии [935, 936, 1040] является следствием несинхронного появления признаков нарождающегося таксона. При этом организация переходных групп обычно совмещает признаки предкового и дочернего таксонов с чертами собственной специализации. Реальные предковые формы лишь в редких случаях бывают промежуточными по организации, как это обычно полагалось до начала палеонтологической эры в филогенетике [1033, 1034]. Сама возможность поляризации признаков по уровням говорит о направленности и преемственности несинхронных, но взаимообусловленных семофилезов целого.
Особенно ярко противоречие мозаичности и ортогенетической направленности эволюции выразилось в концепции аристогенеза Г. Осборна [291, 2046, 2048, 2053]. Однако все эти факты приходят в согласие, если предположить, что главной причиной мозаичности является запрет на синхронную оптимизацию многих разноплановых эктосоматических подсистем целого и большие порции кратковременно получаемой новизны, которых требует такая синхронная оптимизация. Это хорошо согласуется с мнением А. Л. Тахтаджяна, что при освоении новой адаптивной зоны меняются в первую очередь те подсистемы, которые определяют адаптацию к новым условиям; остальные подсистемы отстают в своем филогенетическом развитии [935, 936, 1040]. При этом эффект мозаичной эволюции представляет собой следствие не мозаичной организации, а, напротив, целостности филогенетической системы и согласованности составляющих его семофилезов. Таким образом, мозаичность эволюции - следствие комплексной оптимизации целого, которая возможна только как процесс последовательной (несинхронной) оптимизации отдельных его блоков, смены ролей экто- и эндосоматических подсистем, а значит, и смены направлений специализации.
Правило инадаптивной специализации. Правило отражает необходимую односторонность и противоречивость специализации целого. Накапливающиеся в этом процессе противоречия целого и частей (нарушение адекватности внутреннего умвельта), устойчивости и способности к изменению (ограничения свободы творчества) и т. п. в конечном счете начинают тормозить дальнейшую прогрессивную специализацию в данном направлении, затрудняя смену ее направления, ставя развивающуюся систему под угрозу гибели. Все эти оборотные стороны специализации могут рассматриваться как инадаптивные ее последствия, "плата" за приобретенную устойчивость - адаптацию [495].
Инадаптивная специализация, таким образом, не особая форма специализации, а термин для обозначения одного из имманентно присущих ей свойств. Интерпретация особенностей организации одного таксона как инадаптивных по сравнению с другим позволяет объяснить причину явления конкурентного вытеснения, биологического регресса инадаптивного таксона на фоне процветания экологически близкого, но эвадаптивного, по терминологии А. П. Расницына, таксона [859]. Экологически близкие таксоны, достигшие сравнимого уровня специализации, могут отличаться по инадаптивным последствиям и, следовательно, перспективам будущей специализации.
Д. Роза обратил внимание на относительную "скороспелость" инадаптивных и "запоздалость" эвадаптивных таксонов [291]. Действительно, тем легче и быстрее протекает прогрессивная специализация, а инадаптивность приобретает критические размеры, чем более односторонен этот процесс, чем меньшее число эктосоматических подсистем его определяет, чем более они индивидуальны и чем в большей степени определяют судьбу эндосоматических подсистем целого. "Запоздалые" таксоны, напротив, благодаря низкому темпу специализации могут обеспечить более гармоническое развитие целого с участием большего числа эктосоматических подсистем и более полной компенсацией их одностороннего развития эндосоматическими подсистемами, т. е. позволить менее противоречивое и более компромиссное развитие функциональных систем целого [859]. В процессе филогенеза крупного таксона первым биологического прогресса достигает обычно наиболее инадаптивный подтаксон, который вскоре последовательно сменяют все более эвадаптивные подтаксоны. Наличие двух таких смен Д. Роза считал правилом.
Правило итеративной эволюции. Если некоторое направление прогрессивной специализации исчерпало себя вследствие накопления груза инадаптивности (что завершается той или иной формой биологического регресса), то в сходном направлении может возникнуть новый цикл прогрессивной специализации, начальные этапы которого, естественно, будут уступать по уровню специализации конечным звеньям предыдущего цикла. Такой повторяющийся во времени процесс прогрессивной специализации в очень близком направлении, начинающийся каждый раз от родственных, часто персистирующих, но слабоспециализированных в данном направлении форм, составляет содержание итеративной эволюции [864]. Иногда итерация (возврат) осуществляется путем вторичной деспециализации (например, путем фетализации) инадаптивного филума [248]. Итеративная эволюция - особый случай широко понимаемого явления параллелизма. Повторение очень близких направлений специализации свидетельствует о большой специализации предка итеративных филогенезов, что хорошо согласуется с его персистированием. Сами итеративные филогенезы чаще всего являются гиперморфными линиями, завершающимися вымиранием или реже фетализацией, но не персистированием. Итеративная эволюция показывает, что порой проще создать новое, сходное с прежним, направление специализации, чем избавиться от накопленного груза инадаптивности.
Примеры итеративной эволюции известны в филогенезе двустворчатых моллюсков [24], аммонитов [887, 864], акул [248] и других групп.
Правило адаптивной радиации. Стабилизация новообразования на каждом уровне рассмотрения по отношению к итогу прежней стабилизации представляет собой в большей или меньшей степени смену направления специализации. Это хорошо согласуется с представлением И. И. Шмальгаузена об аллогенезе [805, 1197], при котором освоение таксоном новой адаптивной зоны начинается с одной или немногих субзон [952]. Напротив, при разделении первоначально широкой адаптивной зоны предка на субзоны [952] причиной адаптивной радиации является специализация каждого из подтаксонов, достигающаяся путем интенсивной эксплуатации части ресурсов адаптивной зоны предка. Эти два способа адаптивной радиации нельзя считать альтернативными: любая специализация до известной степени изменяет адаптивную зону, а любая смена адаптивных зон немыслима вне специализации.
Однако в пределах заданного направления прогрессивной специализации таксономический уровень, на котором осуществляется адаптивная радиация, будет убывать пропорционально достигнутому уровню специализации (рис. 18). После эманации, если не наступает вымирание, прежде всего происходит стабилизация структурного типа нового таксона, останавливающая дальнейшую эманацию. На базе еще достаточно широкого спектра изменчивости, но уже более совершенного механизма запоминания новообразований активизируется адаптивная радиация. Поскольку устойчивость нового таксона еще невелика, большинство радиирующих ветвей окажутся недолговечными. Более продолжительную историю будут иметь ветви, продолжающие процесс специализации в избранном направлении, а значит, приобретающие все большую ограниченность и устойчивость (адаптацию). Именно этот процесс, первоначально расчленяющий единую адаптивную зону на субзоны, со временем начинает постепенно замедлять дальнейшую адаптивную радиацию на данном таксономическом уровне и все больше стимулировать ее на следующем, более низком (см. рис. 9). Возрастающая устойчивость предыдущего иерархического уровня будет оказывать все более канализирующее воздействие на эволюцию нижележащего, вводя его в русло прогрессивной специализации.
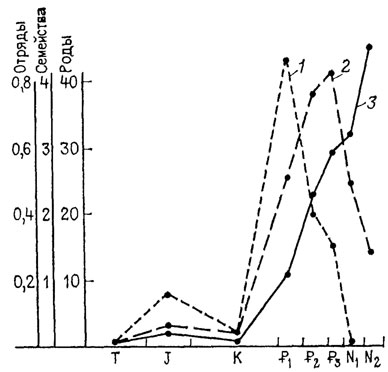
Рис. 18. Падение таксономического уровня, на котором протекала адаптивная радиация млекопитающих, по мере их прогрессивной специализации. По Дж. Г. Симпсону. Скорость появления (число соответствующих таксонов за 1 млн. лет - отложено по ординате), 1 - отрядов 2 - семейств, 3 - родов
Правила типичной смены фаз филогенетического цикла и адаптиогенеза отражают характерную последовательность событий процесса прогрессивной специализации (см. рис. 10). Ему всегда предшествует эманация (типолиз). Стабилизация основ организации нового таксона соответствует фазе типогенеза [2223, 2224]. Эти обычно плохо документированные краткие фазы часто рассматривают как одну [291, 952, 721, 1670]. Фиксация таксономического уровня рассмотрения специализации позволяет в начале этого процесса выделить фазу кладогенеза [1773] - адаптивной радиации таксона на подтаксоны следующего за ним уровня [2045, 2050]. Сменяющая ее фаза телогенеза [805, 1040, 1197] характеризуется угнетением адаптивной радиации и перемещением ее на более низкие уровни (см. рис. 18), а также развитием направленной филетической эволюции [952]. Прогрессивная стабилизация организации начинает постепенно останавливать и филетическую эволюцию. Если на этом пути груз инадаптивности быстро становится критическим, то телогенез сменяется фазой гипергенеза [805, 1197]. Неадекватность внешнего умвельта внутреннему в сочетании с высокой внутренней устойчивостью (константностью) организации делает гипергенную специализацию экологически неустойчивой и практически необратимой. Гипергенез часто бывает связан с резким снижением (некоторые островные формы; гипергенез перигамических признаков) или усугублением межвидовой борьбы. Фаза гипергенеза обычно бывает короткой и завершается вымиранием [805, 1197].
Если груз инадаптивности не слишком велик, а экологическая ситуация благоприятна, то телогенез может смениться стасигенезом [1771, 1772], для которого характерен очень устойчивый структурный тип. Эволюционируют очень немногие "поверхностные" признаки, направленность преобразований мала, некоторые признаки обратимы (брадителия [952]), характерно персистирование, иногда весьма продолжительное. Именно среди стасигенных форм обнаруживаются "живые ископаемые" [936].
Полный филогенетический цикл адаптиогенеза завершается эманацией, ведущей к вымиранию или смене направления специализации. Причиной этого являются издержки высокой устойчивости и инадаптивности, не обеспечивающие необходимый прирост или смену приспособленности в рамках прежнего направления прогрессивной специализации (эффект "черной королевы" Л. Ван Валена [2421-2423]).
Из всех онтогенетических стадий перспективы специализации: при прочих равных условиях максимальны для дифинитивной стадии [1178-1181]. Поэтому частичный или практически полный отказ от нее путем фетализации или неотении часто в достаточной мере понижает груз инадаптивности, порождая обычно близкое к прежнему, иногда итеративное, направление специализации. Примеры такого рода описаны среди аммонитов [887], акул [248] и других групп. Такое отступление на исторически в значительной мере подготовленные позиции и потому наименее опасное можно назвать гипогенезом [1197]. Особенно широко гипогенез распространен у форм с относительно слабоэмбрионизованным развитием, поскольку отступать легко только на свободно живущую стадию онтогенеза. Поэтому гипогенез характерен, например, для эволюции анамний, но не характерен для амниот. При чередовании активных и покоящихся стадий (членистоногие) даже значительная эмбрионизация - не помеха [560, 1061, 1062, 1063].
Итак, основными стадиями филогенетического цикла адаптиогенеза являются: типолиз (эманация, дестабилизация) - кладогенез (адаптивная радиация) - телогенез (прогрессивная специализация в самом типичном проявлении) - стасигенез (или гипергенез). Главная причина ограниченности, направленности и закономерности филогенетического цикла - издержки роста устойчивости - качественно подобна таковой и для онтогенеза. Следовательно, многократно формулированное [250] и столь же часто отвергавшееся [291] правило подобия онто- и филогенетических циклов имеет больше здравого смысла, чем принято считать.
Правило прогрессивной эволюции или ароморфоза. Эволюция не ограничивается только частным морфофизиологическим прогрессом немногих эктосоматических подсистем организма (специализация). Последовательно сменяющие друг друга направления специализации могут привести к более или менее комплексным преобразованиям организации, охватывающим разнообразные функциональные подсистемы целого, выводящие своего обладателя на новый более высокий уровень организации по сравнению с его предками. Обычно это сопровождается проникновением в ранее недоступную адаптивную зону или образованием новой адаптивной зоны путем принципиальной реорганизации отношений с ранее эксплуатировавшимися факторами среды. Такого рода изменения, связанные с общим прогрессом организации, А. Н. Северцов назвал ароморфозами [926]. Комплексность и взаимная согласованность прогрессивных изменений повышает общее организационное превосходство обладателя ароморфоза над средой, позволяющее, в силу теоремы о необходимом разнообразии [1163, 1223], более разнообразно, активно и, главное, избирательно относиться к факторам среды, дифференцированно оценивая их роль в собственном умвельте. Организационное превосходство- наиболее общая предпосылка выработки специфической адаптации к факторам среды, основанная на росте жизнеспособности, а не плодовитости в общем балансе приспособленности [1192, 1197].
Последнее обычно влечет рационализацию отношений организма со средой и рост коэффициента полезного действия эксплуатации ресурсов [689].
Возникновение ароморфозов наблюдается в немногих филогенетических ветвях на каждом данном уровне рассмотрения, но наличие известного количества ароморфных групп в любой момент геологической истории свидетельствует о характерности этого явления для филогенеза в целом. Значительные биологические достоинства ароморфоза, требующие высокого уровня структурной новизны, хорошо согласуются с наличием сильных ограничений на эволюцию в этом направлении. Общий прогресс организации не удается уложить в рамки одного направления специализации, поскольку либо уровень рассмотрения оказывается настолько общим, что данное преобразование выглядит достаточно частной специализацией, либо, при понижении уровня рассмотрения, оно оказывается настолько сложным, что сталкивается с запретом на синхронную оптимизацию многих параметров целого и на большие порции кратковременно получаемой новизны, необходимые для такой синхронной оптимизации.
Неопределенно длительный и направленный прогресс адаптации невозможен в силу возникающего при этом конфликта устойчивости (приобретенного уровня адаптации) и свободы творчества (перспектив будущей адаптации). Отсюда нами был сделан вывод: арогенез как специальная форма эволюции [1040], ведущая непосредственно (без смены модуса) к ароморфозу, оказывается теоретически невозможным. Необходимость смены направлений специализации говорит о риске вымирания на пути к достижению ароморфоза.
Правило ключевого ароморфоза. Н. Н. Иорданский [444, 445, 447] первый подытожил различные соображения о невозможности ароморфной эволюции: 1) путем параллельного синхронного приобретения всего синдрома ароморфных признаков; 2) путем случайного поиска каждого отдельного признака весьма сложного и взаимосогласованного синдрома ароморфных признаков; 3) при отсутствии априорной направленности такого процесса определенными начальными условиями. Главная проблема ароморфной эволюции, по Н. Н. Иорданскому, - вскрытие механизма, направляющего последовательное приобретение сложного комплекса взаимосогласованных признаков в филогенезе. Решать эту проблему он предлагает в концепции ключевого ароморфоза, явившейся развитием представления Э. Майра [594] о ключевых признаках.
Ключевой ароморфоз - это морфофизиологический признак или свойство, приобретение которого имеет не только важное значение для своего обладателя (благодаря чему он только и может стабилизироваться [887, 1178-1181] и выполнить свою ключевую роль), но также существенно изменяет взаимоотношения его функциональных подсистем (внутреннего умвельта). Обычно это происходит путем снятия одного из коренных инадаптивных противоречий организации его филогенетического предка, ведя к переоценке значимости, ценности, функциональных подсистем организма (эволюирующей системы) в целом [859]. Тем самым оно (снятие) открывает на базе прежней организации новое перспективное направление специализации, новые возможности функционирования и эволюции функциональных подсистем целого.
Наиболее проработанным примером ключевого ароморфоза является становление синдрома дефинитивных признаков рептилий на базе их амфибийных предков. В качестве ключевого ароморфоза предлагается рассматривать активизацию легочного дыхания путем оптимизации функции всасывания воздуха в легкие. Это обусловило освобождение кожи от присущей амфибиям функции интенсивного газообмена, что, в свою очередь, позволило сделать кожу сухой, ороговевающей, служащей механической и гидроизолирующей защитой. Освобождение венозной крови от функции переноса кислорода от кожи к сердцу дало возможность разделить венозный и артериальный токи крови, вплоть до формирования четырехкамерного сердца. На рис. 19 показано, что появление многих рептилийных черт и даже некоторых признаков птиц и млекопитающих обусловлено выявленным Н. Н. Иорданским [444, 445] ключевым ароморфозом. Им же приведено еще несколько примеров из разных групп животных [444]. Ключевая роль оперения в становлении синдрома дефинитивных признаков птиц [865] показана на рис. 20. Большинство рассмотренных в этих примерах признаков выводятся не непосредственно из ключевого ароморфоза, а из свойств, порожденных его наличием.
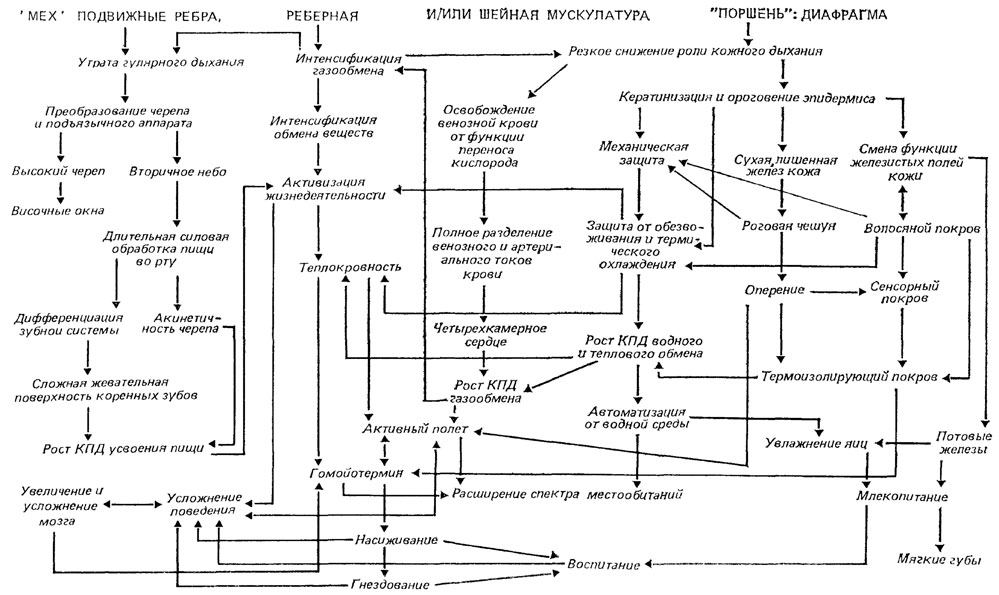
Рис. 19. Активизация функции легочного дыхания как ключевой ароморфоз синдрома дефинитивных признаков рептильного уровня организации (курсив) и некоторых признаков уровня организации высших позвоночных. Составил А. С Раутиан по данным Н. Н. Иорданского с дополнениями
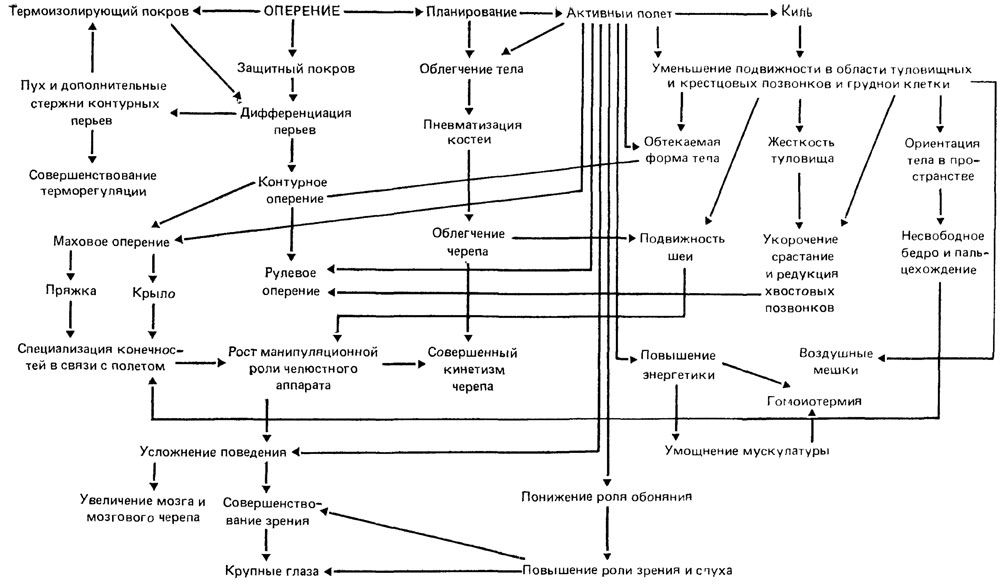
Рис. 20. Наличие оперения как ключевой ароморфоз синдрома признаков уровня организации птиц. Составил А. С. Раутиан
Ключевую роль в становлении млекопитающих сыграли оптимизация силовой обработки пищи во рту и особенности строения нижней челюсти их предков [12, 444], а также, вероятно, волосяной покров [1033]. Ключевым ароморфозом в становлении амфибий было, по-видимому, прикрепление конечностей к осевому скелету независимо от головы, поскольку легкие были уже у кистеперых и двоякодышащих рыб [1188, 1190], а голая лиссамфибийная кожа, очевидно, является поздним приобретением.
Любой признак или свойство, прошедшие филогенетическую стабилизацию, имеет ту или иную, большую или меньшую ключевую роль. Следовательно, термин "ключевой ароморфоз" выделяет не столько особую категорию признаков (хотя ключевая роль некоторых из них выражена необыкновенно сильно), сколько подчеркивает их роль в освобождении латентных потенций организации филогенетического предка, актуализация которых была затруднена или невозможна до возникновения ключевого ароморфоза. Специфика ключевого ароморфоза заключается в далекоидущих последствиях его приобретения и значимости этих последствий для их обладателя. Поскольку ключевой ароморфоз производит реальную (а не только ретроспективную) переоценку функциональных подсистем организма [859], его ключевая роль в принципе может быть установлена до актуализации его последствий в филогенезе [444].
Правила параллельной эволюции, гомологических и гетерологических рядов Э. Д. Копа. Представление о ключевом ароморфозе хорошо согласуется с широким распространением филогенетических параллелизмов, а применительно к проблеме прогресса - с феноменом параллельного достижения нового общего уровня организации несколькими независимыми филетическими линиями. Независимые потомки обладателя ключевого ароморфоза имеют реальную возможность более или менее полной параллельной актуализации его последствий. На рис. 13-14 приведен такой пример из истории зверообразных рептилий и ранних млекопитающих [594, 1032].
Ярчайшее проявление параллельной эволюции - описанные Э. Копом гомологические и гетерологические ряды признаков, отражающие процесс формирования гомопластических сходств в независимых филетических линиях [1457]. На рис. 13-14 показаны примеры таких рядов, а на рис. 15 - схема филогенеза, в котором они необходимо возникают (ср. с классической моделью филогенеза на рис. 21). Система гомологических и гетерологических рядов представляет собой внешнее выражение процесса закономерного развертывания последствий приобретения ключевого ароморфоза ближайшим общим предком всех филетических линий, демонстрируя общее направление специализации, характерное для всей группы в целом.
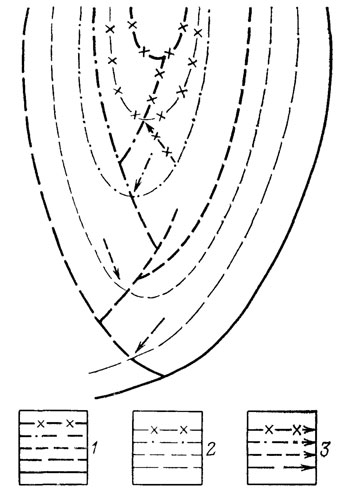
Рис. 21. Схема филогенеза в соответствии с классической монофилетической концепцией Ч. Дарвина. Составил А. С. Раутиан. Усл. обозначения 1 и 2 - см на рис 15; 3 - моменты арогенных преобразований организации, знаменующих переход на новый уровень организации
Наиболее общей причиной параллелизмов вообще и параллельного перехода на новый уровень организации, в частности, является сильный эффект преемственности, т. е. эволюционной инерции у потомков последствий приобретения разного рода ключевых признаков их предками. О правиле эволюционной инерции писали многие палеонтологи [291] и неонтологи [721]. Еще в 1878 г. К. А. Тимирязев [1048] дал верное объяснение феномена инерции через явления наследственности. Можно сказать, что наличие параллелизмов в эволюции следует считать правилом, а их отсутствие - исключением.
Правило монофилии. Независимое достижение несколькими филетическими линиями нового более высокого уровня организации считается наиболее распространенным и трудно снимаемым противоречием принципу монофилии. Поэтому мы остановимся только на этом аспекте проблемы. Более полный обзор проблемы моно- и полифилии опубликован в работе [110].
Кажущееся противоречие монофилии и параллелизма является следствием неправильно сформулированного вопроса. Параллелизм и гомопластическое сходство, возникающие независимо в различных филетических линиях потомков, можно объяснить через ключевую роль гомогенетических сходств, восходящих к общему предку всех линий развития потомков. Одновременно правило ключевого ароморфоза позволяет лучше понять процедуру становления ароморфоза. Направленность его начальными условиями делает его не только возможным, но и достаточно вероятным. Таким образом, явления параллелизма и независимого достижения нового более высокого уровня организации несколькими филетическими линиями не только не противоречат представлению о монофилетическом предке, но остро нуждаются в нем для своего объяснения.
Где же возникла ложная дилемма? Классическая концепция монофилетического филогенеза исходила из преувеличенной роли случайных явлений в эволюции [74, 590, 975, 1137]. Синдром признаков каждой независимой филетической линии считался удачным и потому неповторимым стечением обстоятельств. Параллельный переход на новый уровень организации казался чудом. Как мы знаем, лишь на первый взгляд это хорошо согласуется с правилами мозаичности эволюции и гетеробатмии. Понятия уровня организации и монофилетического таксона казались совпадающими (см. рис. 21): уникальный синдром ароморфных признаков "вырезает" на филогенетическом древе монофилетический таксон. Конкретное течение филогенеза при этом оставалось в стороне. Не случайно термин "ароморфоз" казался слишком морфологически односторонним, а заменяющие его более динамические термины "арогенез" и "анагенез" появились соответственно в 1951 и 1954 гг. [1040].
Однако попробуем выявить совместимые с классической гипотезой черты арогенеза. Становление ароморфоза должно быть продолжительным процессом, иначе трудно понять, как на основе случайного поиска можно быстро собрать сложный и согласованный синдром признаков. Собственно арогенез - переход на новый уровень - краток и связан с приобретением последнего недостающего для выхода на новый уровень признака. Ничего особенного нет ни в самом процессе арогенеза (всего лишь приобретение одного из признаков), ни в последнем признаке, если не считать, что именно его не хватало. Происходит переход количественных изменений в качественные без качественного изменения приводящего к этому процесса арогенеза. Именно поэтому термины для обозначения такого безликого процесса появились позже термина, обозначающего качественное изменение организации. Противники такого взгляда обвиняли "дарвинистов" (очевидно, что рассмотренная концепция не имеет отношения к ядру дарвинизма - теории отбора) в пропаганде "тривиальности прогресса" [590, 1137]. Для филогенеза с охарактеризованными выше свойствами явления гомоплазии, параллелизма, гомологических и гетерологических рядов должны быть нетипичны, что противоречит обширному классу фактов и эмпирических обобщений, в первую очередь палеонтологических.
Классической концепцией не учтено, что система с большой случайной составляющей поведения не может иметь ни совершенного механизма памяти, ни точного механизма запоминания. Следовательно, апелляция к длительности процесса случайного поиска не спасает концепцию: за большое время система успеет забыть приобретенное ранее. При допущении, что монофилетический таксон соответствует определенному уровню организации, любое расхождение уровня и монофилетического основания пучка родственных филетических линий должно рассматриваться как нарушение принципа монофилии. На самом деле это противоречило не принципу монофилии, а лишь филогенетической теории, построенной с его использованием. Монофилетическое основание таксона всегда древнее возникновения нового уровня и находится, иногда глубоко (см. рис. 13-14), в недрах предыдущего уровня организации. Это естественный ход вещей, поскольку ключевой ароморфоз должен предшествовать актуализации последствий его приобретения. Иными словами, проблема заключается не в предпочтении вертикальной или горизонтальной классификаций, а в обнаружении глубокого филогенетического смысла в некоторых горизонтальных объединениях и причинной связи гомопластических сходств с гомофилетическими. Оба типа классификации отражают дополнительные аспекты филогении, а их суперпозиция вносит в систему элемент комбинативности, о котором писал А. А. Любищев [592]. Возможный вариант их совмещения на филогенетических схемах продемонстрирован на рис. 13-15.
Интересно отметить, что несовпадение и самостоятельную ценность понятий уровня организации и монофилетического таксона принимали за нарушение принципа монофилии не только дарвинисты, сформулировавшие классическую концепцию монофилетической филогении, но и их противники, справедливо обратившие внимание на переоценку дарвинистами роли случайности [74, 307, 590,975, 1137].
Мысль об односторонности классической концепции вызревает и в рамках синтетической теории эволюции. Определение параллелизма как независимого развития сходных признаков в двух или более линиях, происходящих от общих предков, основанного на характеристиках этих предков или направляемого этими характеристиками [595], в зачатке содержит главную идею ключевого ароморфоза в приложении к явлениям параллелизма, а также мысль о причинной связи гомоплазии с гомофилией. Характерна эволюция взглядов Э. Майра, видевшего прежде в горизонтальной классификации лишь прагматические функции [598], но позже склонившегося к допущению элементов горизонтальной классификации в филогенетической системе [595]. Идеей компромисса вертикальной и горизонтальной классификаций проникнута и концепция широкой монофилии Дж. Г. Симпсона [110].
Правило аристогенеза. Это правило, сформулированное Г. Ф. Осборном [2052-2055], гласит: в системе родственных филетических линий, как правило, наблюдается процесс параллельного, ограниченного и направленного творчества - аристогенеза. Параллелизм Г. Ф. Осборн оценивал как внешнее выражение ограниченности и направленности филогенеза. Глубокое структурное единство параллельно и независимо возникающих гомопластических сходств свидетельствовало о существовании механизма латентной, или потенциальной гомологии, природу которого он видел во внутренних наследственных свойствах организмов. Дальнейшим развитием этого положения явилось правило ключевого ароморфоза. Представление об определяющей роли факторов внешней среды в параллельном происхождении гомоплазий Г. Ф. Осборн справедливо считал надуманным, противоречащим опыту и здравому смыслу. Внешние условия могли вызвать сходные формы реагирования только на базе общей наследственной, исторической основы. Единство латентных потенций естественно было связать с единством начальных условий для всей системы родственных, параллельно развивающихся филетических линий, т. е. с их монофилией. Значительное дальнодействие латентных потенций (правило эволюционной инерции) связано, очевидно, с сильным эффектом преемственности.
Гомопластические сходства, возникающие в системе параллельно развивающихся филетических линий, образуют гомо- и гетерологические ряды Э. Д. Копа (см. рис. 13-15). Приращение гомологического ряда новым членом - ректиградация (по терминологии Г. Осборна) - представляет собой на каждом данном уровне исследования филогенеза возникновение нового качественного признака. Ректиградации - сравнительно редкие эпизодические события, носящие характер очень небольшого направленного скачка, подобного мутации В. Ваагена [737, 2438]. Новый признак обычно появляется в очень слабом, едва заметном выражении и охватывает сразу если не всех, то многих особей популяции ("перечеканка" форм, по Л. С. Бергу [74]). Такая картина, очевидно, связана с тем, что начальные фазы появления, стабилизации и включения нового признака в адаптивную норму ускользают от наблюдения на ископаемом материале. В случае массовых ископаемых остатков некоторые из таких эпизодов могут быть восполнены. Так, исследуя ректиградации, появляющиеся на зубах акул (которые в некоторых местонахождениях образуют массовые скопления), Л. С. Гликман [248] наблюдал появление некоторых из них с частотой в популяции порядка 10-4. Этот уровень частоты обычен для уклонений (аберраций), не включенных в адаптивную норму у рецентных организмов [514, 886]. Подобные новообразования, называемые Л. С. Гликманом фенами, могут не только появляться, но и исчезать из популяции, так и не получив широкого распространения. Последнее не отмечается Г. Ф. Осборном, что, по-видимому, можно интерпретировать как результат обнаружения ректиградации уже на стадии специализации.
Признаки, появляющиеся в результате ректиградации, испытывают в дальнейшем очень постепенную количественную (аллойометрическую, по Г. Ф. Осборну) эволюцию, выражающуюся обычно в усилении выражения качественного признака и в его внутренней дифференцировке. Возникающие при такой дифференцировке элементы могут представлять собой ректиградации следующего, нижележащего иерархического уровня. По мере аллойометрической специализации признака возрастает наглядность его адаптивного значения в рамках целого. Это хорошо согласуется с представлением о семофилезе эктосоматических подсистем.
Плавный, постепенный и постоянно идущий аллойометрический семофилез лишь изредка прерывается ректиградациями. Это эмпирическое обобщение Г. Осборна, сформулированное применительно к семофилезу, хорошо согласуется с правилом возникновения нового наследственного признака В. Е. Руженцева и Правилом "перемежающегося равновесия".
Параллелизм ректиградаций, ведущий к появлению гомоплазий, свидетельствует об их закономерности и направленности. Гомологические ряды Э. Д. Копа образованы рядами гомопластических признаков, а моменты возникновения каждого из них представляют собой ректиградацию. Гетерологические ряды отражают последовательное прохождение одних и тех же качественных событий разными филетическими линиями одного монофилетического ансамбля. Каждый гетерологический ряд - своего рода уровень организации, независимо достигнутый несколькими филетическими линиями. Параллельное восхождение на более высокие уровни, связанные с приобретением значительных комплексных отличий (правило ароморфоза), осуществляется последовательно по ступеням гетерологических рядов. Восходящая последовательность таких рядов является филогенетическим выражением правила Ж. Кювье об известной градации форм в рамках большого структурного типа [470].
Правило аристогенеза в отличие от классической филогенетической концепции соединяет правила параллельной эволюции, гомологических и гетерологических рядов с правилом монофилии, отказываясь от представления о тождестве понятий уровня организации и монофилетического таксона. Все эти правила свидетельствуют о глубоком филогенетическом смысле и взаимной дополнительности вертикальной и горизонтальной классификации.
Саму возможность наблюдения аристогенеза в форме гомологических и гетерологических рядов (т. е. параллелизмов) Г. Ф. Осборн связывал с предварительной адаптивной радиацией предковой формы в систему независимых филетических линий [2045, 2050] - фаза кладогенеза. Аристогенез как ограниченный и направленный процесс градации гетерологических рядов особенно ярко проявляется в фазе телогенеза, в истории длительно сохраняющихся филетических линий, идущих но пути прогрессивной специализации. Угнетение адаптивной радиации при переходе к телогенезу Г. Ф. Осборн объяснял заполнением адаптивной зоны, доступной представителям данного таксона. Применительно к эволюции таксона в экосистеме этот тезис был значительно развит В. В. Жерихиным [367, 368]. Однако это объяснение, будучи верным, не является полным. Факт адаптивной радиации связан с относительно небольшой устойчивостью эволюирующей системы. Поэтому длительное существование и значительное обособление радиировавших ветвей в этом состоянии немыслимо. Длительную историю, а следовательно, и перспективу значительной актуализации латентных потенций, заключенных в разного рода ключевых признаках, получают лишь те филетические линии, которые идут по пути роста устойчивости - прогрессивной специализации.
Главным противоречием правилу аристогенеза в мировоззрении Осборна оказалось правило мозаичной эволюции, которое он толковал как свидетельство свободы и независимости элементарных биопризнаков в филогенезе. Биопризнаками, по Г. Осборну, являются те крупные или малые признаки, которые на основе данных онтогении (т. е. зоологии, эмбриологии), филогении (т. е. палеонтологии) или наследственности (менделизма) оказываются отдельными и независимыми друг от друга как единицы в процессах наследования, эволюции и индивидуального развития [291, 2048]. Казалось, что правило мозаичной эволюции подтверждало данные современной Т. Ф. Осборну экспериментальной биологии, особенно генетики. Преувеличенное представление о мозаичности организма полностью не изжито и из генетики сегодняшнего дня [1180, 1181]. Филогения представлялась Г. Ф. Осборну в большой степени суммой семофилезов, а казавшаяся ему неясной проблема происхождения видов заменялась более определенной проблемой происхождения признаков [291, 2047]. Однако мозаичность признаков, основанная, казалось, на неопровержимых фактах, вошла в противоречие с представлением о направленности, канализованности и ограниченности аристогенеза, вытекавшим из фактов параллелизма. Г. Ф. Осборн изобрел мозаичный направитель - аристоген, отдельный для каждого биопризнака, что, несомненно, было ошибкой и оставило аристогенез, по его собственным словам, совершенно необъяснимым и таинственным процессом [291, 2051, 2054]. Интерпретация фактов мозаичной эволюции с позиций целостности организма и эволюирующей системы разрешает это противоречие.
Неудачные рассуждения Г. Ф. Осборна о причинах аристогенеза вызвали сдержанное отношение большинства исследователей [270, 291, 297, 952, 2287]. Справедливые упреки в адрес концепции аристогенеза не умаляют, по нашему мнению, ее эмпирического содержания, обобщающего многие (если не большинство) наиболее характерные черты филогенеза, как они представляются палеонтологу.
Закон необратимости эволюции. Чтобы считать преобразование структуры запомненным, достаточно сохранение его последствий в течение одного элементарного звена развития (трех следующих друг за другом состояний системы). В длительном процессе развития эти элементарные звенья перекрывают друг друга: состояния системы, входящие в состав одного звена, могут образовывать с состояниями соседних с ними звеньев, предыдущим и последующим, четыре разных типа сочетаний, таких, чтобы каждое из них оставалось элементарным звеном развития.
Структура, по определению, обладает целостными свойствами. Значит, всякое запоминаемое новообразование опосредованно всеми прежними свойствами структуры, не забытыми в процессе операции выбора, а все сохранившиеся свойства структуры оказываются опосредованными новообразованием. Иными словами, новообразование в процессе запоминания становится неотъемлемым аспектом (а не частью) данного выбора - результатом синтеза старого и нового. Сохранение запомненного в течение одного элементарного звена развития влияет на исход, по крайней мере, двух актов выбора, не входящих в данное элементарное звено развития и следующих за ним. Эти соображения инвариантны для любого элементарного звена развития и последовательности перекрывающихся (на одно или два состояния системы) звеньев произвольной длины. Отсюда следует вывод: последствия данного акта запоминания не окажутся равными нулю, а данный, возникший в результате этого акта, аспект памяти не окажется полностью забытым, спустя любое наперед заданное конечное число элементарных звеньев развития (смен состояний). Последовательность таких элементарных шагов развития является собственным внутренним временем развивающегося объекта [167, 664.]
Противоположное равносильно разрыву преемственности между прошлым и будущим. Полное забвение последствий только одного акта запоминания равносильно разрушению соответствующего ему выбора - состояния структуры в прошлом развитии системы. Следовательно, существование развивающейся структуры в любой данный момент времени определяется не только преемственной связью всех предшествовавших ее состояний вместе взятых, но и каждого из них в отдельности, являющегося связующим звеном между предыдущим и последующим состояниями.
Таким образом, мы получили формальный вывод положения Л. Долло о неразрушимости прошлого [1514]. Прочие аспекты необратимости эволюции, вытекающие из рассмотренных ранее правил филогенеза, обладают меньшей силой и лишь дополнительно подтверждают вывод о неразрушимости прошлого. Однако перечислим и их. Дестабилизация, без которой невозможно возникновение нового, сопряжена, по крайней мере, с частичным забвением прошлого [183]. Если забвение коснется уникальной информации, не сохраняющейся после него в системе ни в одной копии, то в силу принципа невосполнимости полностью утраченной информации А. А. Ляпунова [593] такая утрата принципиально необратима. Поскольку количество доступной для приобретения системой новизны пропорционально количеству забытого ею, то чем больше нового получила система, тем более необратимым, при прочих равных условиях окажется приведший к этому процесс развития.
Всякий шаг в направлении прогрессивной специализации безопаснее, а следовательно, и более вероятен, чем любое изменение направления специализации. Изменение направления развития на противоположное не является исключением, хотя возможность широкого использования прошлого опыта облегчает смену направления специализации. Поэтому явления неотении и фетализации широко распространены в филогенезе. Однако использование прежнего опыта свидетельствует не об обратимости развития, а о его преемственности. Использовать можно только не забытый опыт. Правила ключевого ароморфоза, монофилии, параллелизма, гомологических и гетерологических рядов Э. Д. Копа заставляют задуматься над масштабами преемственной передачи латентных потенций и усомниться в справедливости утверждения о полной невосстановимости вполне атрофированных органов [925, 1024]. Достаточно вспомнить примеры атавизмов, среди которых найдутся и признаки, вполне утраченные в норме. Много ярких примеров такого рода (например, в костном черепе человека) приводит А. П. Быстров [126]. Что касается обратимости эволюции, то ее, несомненно, нет, так как в этих случаях не наблюдается реверсии даже структуры черепа, не говоря уже обо всей организации.
Л. Долло и его последователи полагали, что необратимость эволюции основана на очень низкой вероятности строго поэтапного обратного процесса [211, 957] - инволюции [592, 1106]. Полученный нами формальный вывод свидетельствует о детерминистической природе закона Л. Долло. Действительно, если развитие - процесс стохастический, то осуществленная уже траектория развития - единственная в своем роде незаменимая реальность, воплощением которой является сегодняшняя структура развивающейся системы. Это позволяет придать необратимости развития статус закона, выделив ее среди прочих эмпирических правил филогенеза.
Существуют три наиболее известные формулировки закона Л. Долло. Первая принадлежит Ч. Дарвину: вид, раз исчезнувший, не может появиться снова, если даже снова повторились бы совершенно тождественные условия - органические и неорганические [310]. Две другие формулировки принадлежат Л. Долло: 1) организм не может вернуться (хотя бы частично) К предшествующему состоянию, которое было осуществлено в ряду его предков [291, 1513]; 2) организм никогда не возвращается точно к прежнему состоянию даже в том случае, если он оказывается в условиях существования, тождественных тем, через которые он прошел; но вследствие неразрушимости прошлого он всегда сохраняет след промежуточных этапов, которые были пройдены им [291, 1514, 1516].
Практически все возражения против строгого применения [1024] принципа необратимости касались формулировок Л. Долло, а не Ч. Дарвина. Д. Н. Соболев [975] считал, что необратимости эволюции противоречат факты деспециализации. Очевидно, здесь речь идет об обратимости уровня специализации до известных пределов, а не развития. Не случайно в этой связи Соболев пишет об обратимости онтогенеза. У одноклеточных при этом речь идет о деспециализации, а у многоклеточных - о преемственности. Истинной обратимости онтогенеза нет. Об этом же говорят законы К. Бэра [129] и эквифинальности онтогенеза [336, 1180,. 1181]. Когда Д. Н. Соболев пишет о широком распространении обратного эволюционного развития и возвращении группы к исходному пункту развития, речь всякий раз идет о явлениях итеративной эволюции, неотении или фетализации. Очевидно, сам факт такой "обратимости" морфологически можно установить только в силу ее неполноты. Правда, Д. Н. Соболев, будучи палеонтологом, хотя бы в принципе мог судить об обратимости тождественных прямых и обратимых филогенетических рядов, используя их постепенность и независимо (точнее, достаточна независимо, особенно когда речь идет об аммонитах) данную шкалу геологического времени. Однако он никогда не полагается на чисто стратиграфическую последовательность, и все формы "обратного" ряда получают таксономические названия, а главное - диагнозы, отличные от таковых прямого ряда. В случае достаточно полной обратимости более последовательной была бы позиция В. П. Макридина [603], подвергавшего сомнению принцип необратимости в формулировке Ч. Дарвина. Больше того, полагая, что обратимость может быть скачкообразно-тождественных прямых и обратных филогенетических рядов, истинной обратимости и отвести гипотезу длительного персистирования предковой формы наряду с прогрессивно развивающимися потомками. Тем более полную обратимость не могли эмпирически обнаружить неонтологи, ряд которых высказывал аналогичные суждения [503, 929].
Часть соображений, подобных охарактеризованным, высказывал и П. П. Сушкин [1024, 1025]. Неслучайно он полагал, что эволюция обратима постольку, поскольку онтогенез похож на филогенез [1025]. Здесь опять имеется в виду преемственность, наблюдающаяся при гипогенезе, а не истинная обратимость. Будучи хорошим эмбриологом, П. П. Сушкин, несомненно, имел в виду неполную обратимость. Однако главная особенность позиции П. П. Сушкина, как и многих других исследователей, - противопоставление строгой необратимости организации целого организма возможной обратимости отдельных, особенно простых признаков. Даже для сравнительно несложных комбинаций простых признаков П. П. Сушкин считал обратимость менее вероятной. Эта позиция, где может фигурировать обратимость отдельного, произвольно выделенного из целого признака, вероятность обратимости которого как члена ансамбля понижается, очевидно, носит гносеологический характер. Вероятностный характер закон необратимости в этом случае получает не в результате вероятностной природы необратимости самой эволюции как онтологического явления, а вследствие вероятностной природы любых исторических реконструкций [654, 664] и вообще любых эмпирических суждений, основанных на ограниченном материале [835]. Такой подход к проблеме обратимости вполне оправдан, поскольку эволюция, как и любая реальность, не доступна для конкретного исследования во всей полноте и разнообразии своих проявлений. Мы вполне сознаем, что принципиальное решение проблемы необратимости развития оставляет множество гносеологических проблем (обратимости на определенном уровне).
В последнем случае речь идет не о самой эволюции и ее необратимости, а о возможностях, способах и пределах ее познания эмпирическим путем. Именно онтологическая сторона в дискуссиях вокруг закона Долло всегда была главной.
Неполноту чисто эмпирического обоснования своего закона чувствовал и сам Л. Долло, он даже пытался дать ему теоретическое дедуктивное обоснование [291]. Справедливо полагая его нестрогим, П. П. Сушкин стремился оставаться на почве надежных эмпирических фактов. На этом строилась его критика закона Долло. Но при этом П. П. Сушкин не заметил, что обсуждает уже не эволюцию, а нашу способность судить о ней на основе эмпирических данных. И действительно, эта способность оказалась ограниченной, что хорошо согласуется с мнением Л. Долло о неполноте эмпирического обоснования закона.
Если же обратиться к онтологическому аспекту значения термина "признак", а именно в этом смысле мы его употребляем, когда пытаемся судить о самой эволюции (а не способах ее познания), сразу же обнаруживается противопоставление части целому в подходе П. П. Сушкина. Очевидно, говоря об обратимости, предполагается сравнение двух состояний признака. Отождествление признаков означает установление их гомологии [650, 654, 664]. Для этого используют критерии гомологии [469, 2141]. Нас в данном случае интересует критерий положения признака как части или аспекта целого (мерона [650]) в его отношениях (коннексии) к другим признакам того же целого. Без использования этого критерия невозможно установить место мерона в архетипе, а значит, сравнивать соответствующие признаки как мероны. Фактически об этом писал еще Р. Оуэн [468, 469]. Такое отношение к признаку хорошо согласуется с представлением об организме как целом, в котором каждая часть или аспект опосредованы целым. Следовательно, противопоставление отдельного признака ансамблю признаков или целому (вполне допустимое как методический прием) при онтологическом подходе не только противоречит принципу целостности, но незаконна с чисто морфологической точки зрения: не имея возможности сравнивать признаки как таковые (установить их гомологию), мы не можем судить об их тождестве, а значит, и обратимости. С этой точки зрения оказывается справедливой первая из формулировок закона необратимости Л. Долло, встретившая наибольшее количество возражений.
Таким образом, основные возражения против закона необратимости эволюции сводятся к следующим положениям.
1. Повторение (обычно далеко не полное) признаков, в основе которого сами оппоненты видят преемственность, а не обратимость. Об этом свидетельствует интерпретация этих случаев как итеративной эволюции, неотении, фетализации или атавизмов.
2. Обратимость уровня специализации, а не филогенеза как такового.
3. Повторение организации или ее части только на данном, произвольно выбранном уровне рассмотрения (например, на данном уровне изученности). В действительности, повторение не является полным, что обычно следует из анализа методики, которой пользовался оппонент. Обычно сам факт обратимости не может быть эмпирически установлен при достаточной ее полноте.
4. Утверждение невозможности или крайней трудности эмпирической демонстрации необратимости каждого отдельного признака (что особенно важно для палеонтолога с его всегда дефектным материалом).
Поэтому предлагается считать, что правило необратимости эволюции имеет вероятностный характер, хотя в действительности речь идет лишь о вероятности наших суждений об эволюции на основе эмпирических данных.
Таким образом, первые три возражения имеют в основе методические просчеты на уровне интерпретации эмпирических данных; последнее основано на смешении онтологического и гносеологического аспектов проблемы. Не умаляя актуальности и сложности последних, мы считаем, что они не имеют непосредственного отношения к эволюции.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'