
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Палеонтология и палеогеография
Палеонтология, развиваясь в процессе решения задач, выдвигаемых стратиграфией, своим развитием обусловила не только успехи стратиграфии, но и успехи другой не менее важной отрасли геологического знания - палеогеографии. Палеогеографические реконструкции, представляющие синтез данных геологических исследований, находят свое выражение в палеогеографических картах. Основной палеогеографической единицей является фация. Опыт составления палеогеографических карт показал недостаточность трактовки этого понятия как совокупности литологических и палеонтологических особенностей разностей осадочных пород. Такая трактовка понятия фация не требовала от исследователя выяснения обусловленности фациальных различий осадочных пород, ограничивая его задачи лишь установлением этих различий. Без выяснения же их обусловленности нельзя получить материал для палеогеографических выводов. Поэтому проблемы составления палеогеографических карт потребовали пересмотра содержания понятия фация и возвращения к исходному взгляду на это понятие как на среду отложения осадка, характер которой определяет литологические и палеонтологические особенности породы, возникшей из этого осадка, и, следовательно, может быть восстановлен на основе анализа этих особенностей. Области осадконакопления тесно связаны с областями разрушения и выноса, поэтому термин фация должен относиться не только к среде, где происходит осадконакопление, но и к среде разрушения и выноса. При взгляде на фацию только как на среду положительной седиментации нельзя составить полноценную палеогеографическую карту, так как на ней нельзя, например, показать древних рек, а можно показать лишь те их отрезки, где происходило накопление аллювиальных осадков. Области разрушения и выноса имеются не только на континентах, где они фиксируются корой выветривания или поверхностью размыва, но и в море, где течения размывают ранее отложившиеся осадки. Участки морского дна, эродируемые течением, не являются безжизненными. Уплотненное дно, лишенное наносов, представляет удобный субстрат для поселения прикрепляющихся и сверлящих морских животных и растений. Поверхности напластования с остатками прикреплявшихся организмов и следами сверлений дают материал для реконструкции таких фаций прошлого. Морские фации, являющиеся средой разрушения и выноса, не ограничиваются лишь участками морского дна, размываемыми течениями. Разрушению подвергаются также скалистые берега морей. Такие фации береговой зоны древних морей и тем самым береговая линия этих морей восстанавливаются также главным образом на основе находок остатков соответствующих древних организмов. "Нигде береговая линия давно исчезнувших морей так ясно не очерчена, - пишет Динер, - как там, где в древние времена существовало скалистое побережье. На гранитных скалах Богемского массива можно собирать приросшие к скалам балянусы, оставленные морем первого средиземноморского яруса (миоцен)" (1934, стр. 30). Находки раковин раннемеловых устриц, прикрепленных к основанию гранитного уступа, вскрытого денудационными процессами в ю.-з. части Кызыл-Кумов, позволили рассматривать этот уступ как участок скалистого берега раннемелового моря Ср. Азии (Кушнарь, 1937). Равным образом решающее значение имеют палеонтологические данные для суждения о климате и климатических зонах прошлых геологических эпох. Например, наличие годовых колец у ископаемых остатков древесины древних растений, связанное с неравномерностью жизненных условий этих растений, указывает на умеренный климате присущими ему сезонными изменениями (рис. 4). О том же свидетельствуют годовые кольца на раковинах беспозвоночных, например на раковинах пластинчатожаберных моллюсков и брахиопод (рис. 5), а также чередование Зон частых и редких днищ у табулят и других кораллов (рис. 6). Приведенные примеры, подчеркивая большое значение палеонтологических исследований для фациального анализа и решения вопросов палеогеографии, не исчерпывают их значения для этой цели. Почти нет таких вопросов палеогеографии, которые не могли бы в большей или меньшей мере быть освещены данными палеонтологического изучения.

Рис. 5. Годовые кольца на раковине современной Anodonta из Финского залива и франской брахиоподы Atrypa tanaica Nal. Девон Воронежской обл. (колл. Б. П. Марковского)
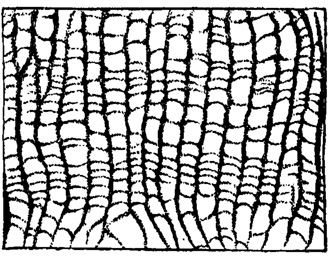
Рис. 6. Palaeofavosites mirus Sck. Чередование зон частых и редких днищ, отмечающее периодичность роста. Шлиф × 4. Силур Прибалтики (Соколов, 1951)
Поэтому проблема составления палеогеографических карт расширила требования, предъявляемые геологией к изучению палеонтологического материала, выдвинув необходимость изучения не только стратиграфического значения ископаемых остатков тех или иных групп древних организмов, но и их палеогеографического значения. Дать правильную оценку палеогеографического значения ископаемых остатков древних организмов можно только на основе изучения экологии этих организмов. Если раньше вопросам экологии древних организмов уделялось сравнительно ограниченное внимание и главным образом в специальных работах, посвященных этим вопросам, то требование оценки палеогеографического значения органических остатков наряду с освещением их стратиграфического значения поставили задачу изучения экологии древних организмов перед автором каждой палеонтологической монографии. Решение этой задачи еще более сблизило палеонтологию с другими отраслями биологического знания, потребовав знания закономерностей, установленных наблюдениями над современными организмами.
Краеугольным камнем биологии является положение о единстве организма и условий его существования. Данное положение требует изучения остатков древних организмов вместе с памятниками условий существования этих организмов. Однако остатки древних организмов не всегда сохраняются там, где обитали данные организмы. Нередко среда погребения отмерших организмов не совпадает со средой существования. Поэтому исследователю прежде всего надо решить, является ли толща, в которой найдены ископаемые остатки изучаемых организмов, памятником среды существования этих организмов или только погребения. Вопросы погребения остатков древних организмов и сохранения их в ископаемом состоянии вызвали новую отрасль знания - учение о закономерностях захоронения остатков древних организмов или тафономию. Данные тафономических исследований имеют большое значение для фациального анализа и палеогеографических реконструкций, давая материал для суждения о характере среды, где происходило захоронение данных органических остатков. Например, беспорядочное скопление разрозненных, частично перебитых створок брахиопод, местами вбитых друг в друга (рис. 7), дает материал для суждения, что данное скопление их связано с переносом волной, давая тем самым указание на небольшую глубину данного участка дна бассейна, поскольку поверхностное волнение обычно не распространяется на более или менее значительную глубину. Вместе с тем отмеченный характер захоронения дает указание на нормальный газовый режим придонных вод, так как волнение, перемешивая воды, обусловливает их хорошую аэрацию.

Рис. 7. Ракушняк, сложенный беспорядочно ориентированными, разрозненными, частично перебитыми, местами вбитыми друг в друга створками брахиопод Ladogia meyendorfii Vern Latnellispirtfer muralis Vern. и др. В. девон Ленинградской обл. (колл. Б. П. Марковского)
Кроме того тафономический анализ дает материал и для суждения об условиях существования древних организмов. Например, в рассмотренном случае данные тафономического анализа показывают, что брахиоподы, которым принадлежали перенесенные волной раковины, обитали также в зоне мелководья, иначе их раковины не могли бы подвергнуться переносу волнами, которые, как отмечено выше, не достигают больших глубин. Давая материал для выяснения экологии древних организмов, тафономическое изучение расширяет этим возможность применения их ископаемых остатков для решения вопросов палеогеографии.
Необходимость тафономического изучения ископаемых органических остатков при палеогеографичесих исследованиях подчеркивается тем, что остатки даже представителей одной и той же систематической и экологической группы организмов в зависимости от характера их ориентировки, сохранности, наличия или отсутствия следов механической сортировки могут указывать на различный характер среды их погребения (табл. I, фиг. 1-3).
Существование организмов определяется не всеми элементами среды, а теми ее факторами, комплекс которых необходим организмам для их жизни. Наследственные особенности организмов, определяемые условиями существования их предков, обусловливают неодинаковые требования различных организмов к количественной стороне этих факторов. Поэтому и реакция организмов на их изменения также различна. Эврифациальные организмы, обладающие широкой экологической валентностью, способны приспособляться к изменениям факторов среды, стенофациальные же узковалентные формы при их изменении погибают. Поэтому ископаемые остатки различных групп древних организмов имеют неодинаковую ценность для фациального анализа и палеогеографических обобщений. Более высокую палеогеографическую ценность имеют остатки древних стенофациальных форм, существовавших лишь в определенных условиях, показателем которых являются их ископаемые остатки.
Таким образом, палеогеографическая ценность ископаемых органических остатков и их стратиграфическая ценность определяются различными экологическими свойствами древних организмов. При использовании ископаемых органических остатков лишь в качестве материала для целей стратиграфии внимание исследователей уделялось главным образом изучению тех групп древних организмов, которые имеют для этих целей более высокую ценность. Изучение палеогеографического значения ископаемых остатков древних организмов прекратило деление последних на более и менее "важные" группы, расширив этим объем палеонтологических исследований и изучение органического мира в целом.
Узкая валентность стенофациальных организмов, исключающая возможность их существования в данной среде при изменении ее факторов, приводит к обеднению качественного состава биоценозов этой среды. Поэтому видовое однообразие служит указанием на специфичность среды, делающее ее пригодной для существования лишь сравнительно немногих видов животных и растений. Для водных организмов относительно высокая температура воды является благоприятным условием существования, поскольку в теплой воде развитие и все жизненные процессы протекают наиболее интенсивно. Поэтому органический мир тропических морей характеризуется чрезвычайным разнообразием. Наоборот, "общей характеристикой населения более холодных вод, в противоположность населению тропической области, является относительная бедность видами при богатстве представителей отдельных видов - иначе качественная бедность при количественном изобилии; это проявляется в населении как бентали, так и пелагиали" (Зернов, 1934, стр. 310). В Индийском океане и в тропической части Тихого океана общее число видов моллюсков достигает 6000, количество же видов моллюсков в Белом море не превышает 140 видов (Коробков, 1950). Для морских организмов не менее важным фактором является соленость воды. Отклонение солености от нормальной (35‰) влечет обеднение видового состава населения осолоненных и опресненных бассейнов. Общее количество видов, обитающих в нормально соленом Северном море около 1500, в восточной же части сильно опресненного Балтийского моря живет не больше 52 видов.
Обусловленность качественной бедности древних биоценозов выясняется на основе экологического анализа их компонентов с учетом отсутствующих экологических групп. Например, исследованиями М. Э. Ноинского установлено обеднение видового состава органических остатков, встреченных в верхнекаменноугольных отложениях Самарской Луки, и бедность их видового состава в нижней перми того же района. Останавливаясь на вопросе обусловленности этого явления, Ноинский пишет: "оскудение и измельчание фауны в верхах карбона Луки не просто лишь кажущееся или вторичное явление, а, действительно, изначальный факт, причину которого нужно искать в свойствах породившего эти осадки бассейна, т. е. мы должны... предположить, что уже вслед за отложением слоев с Spirifer jigulensis Stuck, в данном отрезке русского верхнекаменноугольного моря появились некоторые неблагоприятные для прежнего населения условия, которые затем накоплялись все более и более" (1913, стр. 602). Данные экологического анализа позволили Ноинскому связать эти неблагоприятные условия с обособлением бассейна и его осолонением. "За то, что вымирание... фауны, действительно, связано с обособлением бассейна и накоплением в нем солености, - пишет названный исследователь, - говорит очень определенно самый ход вымирания... Прежде всего и больше всего страдают те именно группы животного царства, представители которых принадлежат к типичным обитателям моря: корненожки, кораллы, брахиоподы и головоногие; напротив, пластинчатожаберные и брюхоногие..., легче приспособляющиеся к изменениям среды, сохраняются... до самого конца" (там же, стр. 624-625). Присутствие в нижнепермских отложениях гипса и ангидрита подтверждает вывод об осолонении бассейна на основе количественного учета видов и данных экологического анализа.
Обеднение видового состава может быть обусловлено также значительной глубиной, неблагоприятным газовым режимом, высокими темпами осадконакопления и т. д. Выяснение обусловленности видовой бедности комплексов ископаемых органических остатков дает поэтому ценный материал для палеогеографии, раскрывая характер среды существования соответствующих древних организмов.
Широкая экологическая валентность эврифациальных организмов не лишает их ископаемых остатков палеогеографического значения. Подвергаясь закономерным изменениям в итоге приспособления к изменяющимся условиям среды, эврифациальные организмы отражают характером своей изменчивости специфику данной среды. Например, раковины особей современного пластинчатожаберного моллюска Mytilas edulis L., живущих у западных берегов Англии, более чем вдвое тяжелее раковин особей этого вида, обитающих в более спокойной Кильской бухте (Книпович, 1938); величина раковины М. edulis L. в этой бухте при солености 15‰ - 100 мм, в Финском заливе - при солености 5‰ - 27 мм (Зернов, 1934). Особи байкальского брюхоногого моллюска Baikalia herderiana Lindh. (рис. 8), обитающие на камнях в прибрежной зоне, имеют раковину с отчетливо выраженной скульптурой, у особей же этого вида, живущих на некоторой глубине на мягком грунте, раковина почти гладкая (Жадин, 1952). Так как организмы зависят не только от абиотических факторов среды, но и от биотических, то изменение последних также накладывает на них определенный отпечаток. Например, раковины балянусов, живущих плотными поселениями в прибойной полосе, тоньше, чем у живущих менее скученно, величина же мягкого тела больше. "В условиях редких поселений, - отмечают В. В. Кузнецов и Т. А. Матвеева, - каждая особь в одиночку противостоит ударной силе волны, и поэтому значительная часть жизненной энергии затрачивается на усиление крепости соединения с субстратом и на повышение плотности домика. В условиях повышенной плотности поселений особи, кроме соединения с субстратом, соединены еще друг с другом; это уменьшает для каждой особи площадь сопротивления движущимся частицам воды и потому большая часть энергии идет на увеличение собственно живых частей организма, не принимающих непосредственного участия в сопротивлении разрушительной силе прибоя" (1949, стр. 415).
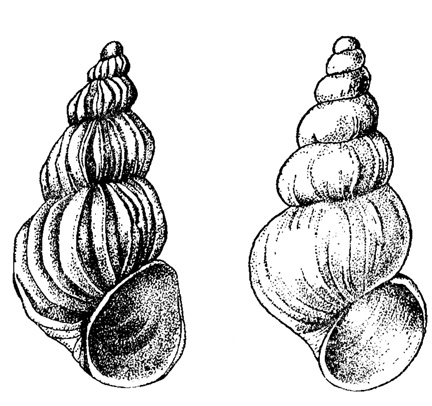
Рис. 8. Baicalia herderiana Lindh. Слева - раковина особи, обитающей на камнях открытых побережий, справа (var. laevis Kosh.) - живущей глубже на мягком грунте. Увел. × 8. Оз. Байкал. (Жадин, 1952)
Выяснение обусловленности экологических изменений древних организмов дает материал для суждения об особенностях данной среды и тем самым материал для палеогеографии. Например, раковины брахиопод, относящихся к виду Cyrtospsrifer archiaci Murch. из основания верхнедевонской соленосной толщи, вскрытой буровыми скважинами в западной части Русской платформы (Белорусская ССР) почти вдвое меньше раковин этого вида из морских отложений того же возраста центральной части платформы. Нельзя не видеть в этом проявления той же закономерности, в силу которой по М. Э. Ноинскому при осолонении позднекаменноугольного бассейна Самарской Луки происходило "оскудение и измельчение фауны". Псевдомонотисовые слои триаса севера СССР содержат многочисленные раковины пластинчатожаберного моллюска Pseudomonotis ochotica Keys, (в настоящее время родовая принадлежность этого вида пересматривается). Более крупные размеры раковин этого вида, по сравнению с раковинами близко родственной кавказской формы, можно рассматривать, с учетом других наблюдений (бедность видового состава сопутствующих форм, преобладание в составе вмещающей толщи сланцев, песчаников и грубообломочных пород), как проявление той общебиологической закономерности, в силу которой "в пределах одного вида особи, живущие в более холодной воде, в среднем крупнее (и часто гораздо крупнее), чем живущие в воде более теплой, или в близких родах или видах обнаруживается такое же различие, и формы, живущие в холодных морях и в холодных глубинных слоях, оказываются очень крупными или даже гигантами по сравнению с жителями более теплых вод" (Книпович, 1938, стр. 351). Таким образом, выяснение обусловленности более крупных размеров раковин рассматриваемого вида по сравнению с раковинами кавказской формы приводит к выводу о более низком температурном режиме триасового моря в северной части Евразии, т. е. к выводу о существовании в триасе климатической зональности.
Приведенные примеры подчеркивают, что изучение однородных явлений (изменений величины древних организмов) в их многообразных связях с другими явлениями (характер сопутствующих форм, состав вмещающей толщи и т. д.) позволяет устанавливать различную обусловленность этих явлений и делать на их основе неоднозначные выводы. Поэтому для правильной оценки палеогеографического значения ископаемых органических остатков, как и для выяснения их стратиграфической ценности, требуется, кроме палеонтологического изучения этих остатков, также геолого-литологическое изучение вмещающих отложений. Таким образом, расширение связи палеонтологии с другими отраслями биологического знания еще теснее связывает ее с геологией.
Палеогеографическое значение имеют не только ископаемые остатки древних организмов, но и сохраняющиеся в ископаемом состоянии следы жизнедеятельности этих организмов. Некоторые из них, как, например, следы передвижения, следы сверления и зарывания всегда автохтонны. Поэтому такие следы жизнедеятельности, указывая на тесную связь соответствующих организмов с данной средой, расширяют представления о составе населения этой среды и дают материал для суждения об ее характере. Например, следы ползания червей, окаймленные бахромой отпечатков параподий, свидетельствуя о принадлежности их многощетинковым червям, дают указание на отложение сохранившего эти следы осадка в морских условиях, поскольку многощетинковые черви принадлежат к числу морских организмов. Находки следов ползания древних морских беспозвоночных вместе со следами передвижения наземных животных служат материалом для установления низменных побережий морей прошлого. По указанию С. А. Зернова, илистые берега Каркинитского залива во время сгона воды ветрами "обнажаются на много сажен, нередко на несколько дней; ил подсыхает, растрескивается на типичные многоугольники; на нем ясно отпечатываются следы разных птиц; ясно видны ходы червей, моллюсков и кучки выкинутой червями земли" (1913, стр. 103).
Большое значение наблюдений над следами жизнедеятельности древних организмов для фациального анализа и палеогеографических выводов подчеркивают данные изучения следов сверления древних морских камнеточцев. В одних случаях они дают указания на течения, размывавшие морское дно, в других - служат материалом для установления скалистых берегов древних морей. Одним из примеров установления скалистого побережья прошлого на основе наблюдений над явлениями сверления является скалистый берег палеогенового моря Ферганы, сложенный в районе г. Ура-Тюбе палеозойскими известняками, источенными палеогеновыми литофагами (Геккер, 1955).
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'