
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Раковина фораминифер
Раковины образованы либо продуктами выделения протоплазмы как в виде органического вещества, так и минеральных солей, либо состоят в основном из различного постороннего материала, заимствованного из окружающей среды. В последнем случае выделяемые протоплазмой вещества служат лишь цементом, скрепляющим посторонние частицы. Соответственно различают раковины секреционные и агглютинированные (от лат. agglutinare - склеивать).
Общим для всех фораминифер является наличие хитиноидной или псевдохитиновой основы их раковин. Псевдохитин - особое органическое вещество, продукт выделения эктоплазмы. Из псевдохитина состоят скелетные образования многих простейших животных, в том числе не известные в ископаемом состоянии раковины фораминифер из отр. Allogromiida.
С. Аверинцев (Averinzew, 1903) указывает, что тектин - так называет он псевдохитин, - в отличие от хитина членистоногих, растворим в минеральных кислотах средней крепости (например в 10-15% соляной кислоте), а также и в щелочах. Милонов реактив окрашивает органическое вещество раковин фораминифер в бледный (благодаря тонкости окрашиваемого объекта) желтовато-красный цвет. По Догелю (1951), псевдохитин - гликопротеид, т. е. комбинация протеина с углеводом, тогда как хитин - вещество, близкое к углеводам.
Скелеты из псевдохитина (хитиноидные) сохраняются в ископаемом состоянии еще реже хитиновых. Хитиноидные скелеты фораминифер, а также их агглютинированные раковины с псевдохитиновым цементом были выделены из нерастворимого в соляной кислоте остатка некоторых известняков ордовика и силура Прибалтики (Eisenack, 1932, 1937, 1954).
Секреционные минерального состава раковины всегда, по-видимому, имеют псевдохитиновую основу, пропитанную минеральными солями; в ископаемом состоянии эта основа не сохраняется. Неорганическими веществами, пропитывающими псевдохитиновую основу, могут быть углекислая известь, обычно в форме кальцита, иногда с довольно значительной примесью углекислого магния и фосфата кальция и магния, либо, в очень редких случаях, - кремнезем. В состав цемента агглютинированных раковин могут входить те же соединения, а также в виде более или менее значительной примеси - сидерит, переходящий в ископаемом состоянии раковин нередко в лимонит. В других случаях водная окись железа входит, по-видимому, в состав раковины еще в прижизненном состоянии фораминифер.
Цемент у многих агглютинированных форм, возможно, и не имеет псевдохитиновой основы, равно как и столбики, грануляции, шипы, натеки и т. п. раковин различных известковых форм. В таких случаях углекислая известь является, по-видимому, лишь продуктом выделения протоплазмы, облекающей собственно раковину или проникающей в имеющиеся в этой последней пустоты.
С переходом раковин в ископаемое состояние часто происходит перекристаллизация и замещение минеральных составных частей скелета.
В состав агглютинированных раковин могут входить посторонние частицы самого различного характера. В большинстве случаев это минеральные частицы: зерна кварца, полевых шпатов, чешуйки слюды, подчас известковые частицы и т. п. Иногда это фрагменты скелета различных других организмов, особенно иглы (спикулы) губок, раковины других видов фораминифер. Так, например, некоторые гипераммины, обитающие в современных морях на поверхности глобигеринового ила,строят свои скелеты из пустых, падающих на дно моря после смерти животного раковин глобигерин. В состав раковины одного из видов рода Ammobaculites из верхнеюрских отложений Поволжья, наряду с многочисленными и разнообразными минеральными частицами, входят отдельные известковые раковины псевдоламаркин. Раковины Clavulina cyclostomata Galloway et Morrey, по данным Субботиной, могут быть нацело образованы мелкими раковинками Gumbelina (рис. 9).

Рис. 9. Clavulina cyclostomata Galloway et Morrey. Агглютинированная раковина, построенная из мелких Gümbelina. В. эоцен, белоглинская свита, С. Кавказ, р. Белая, × 47 (колл. Н. Н. Субботиной). А - вид сбоку; Б - вид со стороны устья
Относительное количество агглютинированных частиц и связующего их цемента у различных видов заметно варьирует. В одних случаях агглютинированные частицы соприкасаются одна с другой подобно камням хорошо сложенной мостовой; у других форм, наоборот, цемент может даже преобладать над посторонними частицами, образующими в таких случаях более или менее часто расположенные вкрапленники. Между этими крайними случаями имеются всевозможные переходы. Показателен в этом отношении морфологический ряд форм, приведенный А. Вудом (Wood, 1949). Так, например, у Clavulina cylindrica Hantken многочисленные различного типа обломки связаны тонкозернистым, известковым цементом. У Vulvulina capreolus Orbigny относительное количество цемента более значительно, а у Ammocibicides proteus Earland и Amm. pontoni Earland его еще больше; в последних двух случаях цемент уже явно преобладает, агглютинированные же зерна маленькие, не совсем равновеликие и с неровными краями. У Gaudryina pupoides Orbigny (в понимании этого вида Брэди) стенка сложена только лишь мелкими частицами углекислой извести, напоминающими образующие цемент частицы у вышеперечисленных форм. Можно предполагать, что у Gaudryina pupoides, а также у имеющих сходную структуру стенки Verneuilina pygmaea (Egger) и ископаемых Didyoconus, Coskinolina и Yaberinella агглютинация полностью замещена секреционным выделением кальцитовых зерен. Сходные отношения, когда и цемент в конечном счете становится в составе раковин преобладающим, отмечаются и в ряде других групп (Ammodiscida, Endothyrida).
Химический состав агглютинированных раковин некоторых современных видов показан ниже (см. Brady, 1879, Виноградов, 1935).

Значительное содержание кремнезема должно быть отнесено за счет агглютинированных кварцевых песчинок.
Необходимо отметить, что фораминиферы с агглютинированными раковинами обладают известной избирательной способностью по отношению к характеру частиц, идущих на построение скелета.
Природа избирательной способности остается до настоящего времени невыясненной. Несомненно, она связана с физико-химическими особенностями протоплазмы. С изменениями условий обитания характер избирательной способности тоже может изменяться, и состав раковины одного и того же вида оказывается в таком случае неодинаковым.
По данным Д. Слама (Slama, 1954), один из видов рода Ammobaculites, обитающий на илистом грунте (в солоноватоводном озере Пончартрен близ Нового Орлеана), строит свою раковину из очень мелких частиц неопределенной формы в соответствии с характером субстрата. В условиях культуры, на субстрате из песчинок 1/4-1/8 мм данный вид использовал этот материал, несмотря на то что отдельные песчинки занимали подчас всю поверхность отдельной камеры раковины. При добавлении к субстрату порошка карборунда, фораминиферы использовали и этот новый материал, причем зерна карборунда вклинивались между крупными песчинками.
Секреционные известковые раковины в подавляющем большинстве случаев состоят из кальцита и, как показали рентгеноскопические исследования, а также применение реакции Мейгена, в виде редких исключений - из арагонита (Hagn, 1953; Troelsen, 1954; Todd and Blackmon, 1956). Наличие арагонитовой раковины - родовой признак, связь которого с условиями внешней среды установить пока не удалось. Раковины смешанного кальцит-арагонитового состава у фораминифер не известны.
Химический состав секреционных известковых раковин нескольких современных видов приведен ниже (см., Brady, 1884, Clarke and Wheeler, 1922, А. П. Виноградов, 1935).
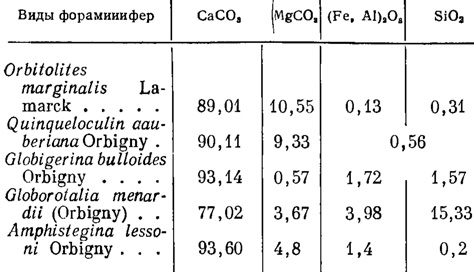
По данным И. Вальтера (Walther, 1893-1894), количество MgCO3 у Orbitolites complanata Lamarck может достигать 12,5%, а у миоценовой Nubecularia novorossica Karrer et Sinzov 26% (в последнем случае, возможно, за счет вторичного обогащения магнием при фоссилизации. А. Ф.).
Стенки известковых раковин могут быть либо видимо бесструктурными, либо зернистыми, либо обладать тонкослоистым, волокнистым или лучистым сложением. У некоторых фораминифер тонкое сложение (микротекстура) стенки раковины обнаруживает, при изучении в прозрачных шлифах, значительную степень дифференцировки, например в отряде Fusulinida. Наконец, у многих фораминифер стенка раковины пронизана многочисленными поровыми каналами, расположенными более или менее равномерно и перпендикулярно поверхности.
Зернистая структура стенки в ряде случаев является безусловно вторичной, связанной с перекристаллизацией; в других случаях зернистая структура, по-видимому, первичная. Подобное предположение подтверждается тем, что при удовлетворительной сохранности скелета лишь один наружный слой двухслойной стенки некоторых эндотирид, текстуляриид и других оказывается зернистым, тогда как внутренний слой в то же время представляется прозрачным и лучистом или вовсе бесструктурным, стекловатым; трудно допустить, чтобы перекристаллизация всегда затрагивала лишь один из слоев. Зернистая структура стенки или какого-либо из ее слоев объясняется беспорядочной ориентировкой плотно прилегающих один к другому мельчайших (до 3 μ) кристаллов кальцита (Липина, 1955). Бесструктурные или радиально лучистые стенки сложены, наоборот, ориентированными в определенном направлении кристаллами. Кристаллическая природа таких раковин фораминифер хорошо выявляется в поляризованном свете (Аверинцев, 1903). Изучение целых раковин фораминифер показывает, что они ведут себя как сферокристаллы или группы их. У Globigerina bulloides Orbigny, Hastigerina, Discorbis sp. и Planorbulina mediterranensis Orbigny каждая камера ведет себя как отрицательный сферокристалл, дающий весьма правильный черный крест при наблюдении коноскопического эффекта.
При наблюдении Hastigerina в поляризованном свете оказывается, что в соответствии с вытянутостью камер крест несколько искажен, лапы креста соединены попарно темными перемычками. При изучении известковых игл Hastigerina хорошо видно, что каждая из них ведет себя как одноосный отрицательный кристалл. Раковины Orbitolites и Peneroplis при разглядывании их с широкой стороны ведут себя как оптически положительные сферокристаллы (нерезкий или едва заметный темный крест), наоборот, при введении гипсовой пластинки оптический характер хорошо выявляется. У Orbitolites можно отчетливо наблюдать, что концентрические стенки, как и стенки плоских сторон, ведут себя оптически иначе в сравнении со стенками, ориентированными радиально; последние, по крайности в тонких шлифах, ведут себя как образованные хорошо выраженными положительными одноосными кристаллами, ориентированными радиально по отношению к центру раковины; концентрические и горизонтальные стенки ведут себя как образованные такими же кристаллами, расположенными тангенциально.
Наличие прободенной или, соответственно, непрободенной раковины - один из наиболее важных систематических признаков фораминифер.
Присутствие поровых каналов, открывающихся на наружной и внутренней поверхностях стенки раковины отверстиями - порами, является признаком относительно высокой степени дифференцировки стенки (рис. 10). Предполагается, что у большинства прободенных фораминифер протоплазма, выполняющая внутренние полости раковины, сообщается при помощи поровых каналов с той частью протоплазмы, которая облекает раковину снаружи.

Рис. 10. Декальцинированная раковина Cibicides lobatulus (Walker et Jacob). Видна слоистая псевдохитиновая основа раковины, пронизанная поровыми канальцами, имеющими каждый собственную выстилку (Rhumbler, 1911)
Очень тонкие поровые каналы (перфорации), диаметром от нескольких десятых долей микрона до 6 μ, наблюдаются у представителей сем. Lagenidae, Polymorphinidae, Buliminidae, Heterohelicidae и др. Более крупные перфорации, от 6 до 15 μ в диаметре, встречаются у многочисленных родов Rotaliidae, Anomalinidae и др., а также у некоторых видов Globigerina, Nonion, Bolivina, Virgulina (Wood, 1949; Le Calvez, 1953). Крупные поровые каналы наблюдаются у сравнительно специализированных форм; у относительно более примитивных фораминифер, по данным Ле Кальве, пористость, как правило, тонкая. По внешнему виду непрободенные известковые раковины фораминифер в наиболее характерных случаях напоминают фарфор или молочное стекло; это объясняется рассеянием света неодинаково ориентированными кристаллами. Прободенные же раковины при том же условии обычно более прозрачны и имеют стекловатый вид, что связано с единообразной ориентировкой кристаллических осей.
Иногда поры на одних частях раковины крупные, а на других относительно мелкие. У некоторых форм с трохоидной раковиной поры на спинной стороне крупные, хорошо видимые при малом увеличении микроскопа; на брюшной же стороне настолько мелки, что при том же увеличении неразличимы. У фузулинид, помимо частых равномерно расположенных тонких пор, иногда с трудом различимых под микроскопом, наблюдаются подчас редкие, значительно более крупные поры, неравномерно рассеянные на перегородках, - септальные поры. У представителей рода Peneroplis и некоторых других прободенной оказывается лишь одна начальная камера, что иногда рассматривается как указание на происхождение пенероплид от прободенных форм. Возможно, однако, что в данном случае имеет место возникновение нового прогрессивного признака на ранней стадии индивидуального развития.
Очень существенно то, что у некоторых фораминифер (Rotaliidae, Anomalinidae, а также, возможно, у представителей других семейств) под прободенным обызвествленным слоем раковины находится хитиноидная перепонка, которая иногда не бывает пронизана порами насквозь. В таких случаях полностью исключается возможность непосредственного сообщения внутренних полостей раковины с внешней средой через поры. У высокоорганизованных фораминифер это сообщение осуществляется более или менее сложно построенной системой каналов.
Кремневые секреционные раковины, если отбросить нечастые случаи образования псевдоморфозов кремнеземом по кальциту, встречаются исключительно редко: у представителей одного сем. Silicinidae, самостоятельность которого оспаривается рядом исследователей, и у некоторых особняком стоящих форм (Silicotextulina). Можно предполагать, что, подобно известковым, кремневые раковины образуются в результате пропитывания псевдохитиновой основы кремнеземом. Но имеется и другое предположение - что кремневые раковины представляют собой только лишь продукт выделения протоплазмы (Le Calvez, 1953). Вопрос о происхождении кремневых фораминифер, в силу невыясненности природы их раковины, остается открытым. Учитывая важное таксономическое значение особенностей биохимической деятельности организмов, в фораминиферах с кремневыми раковинами можно видеть представителей особого филогенетического ответвления. В приведенном ниже систематическом обзоре фораминифер Silicinidae с кремневой раковиной лишь условно отнесены к отр. Ammodiscida.
Под типом строения раковины понимается известная закономерность в относительном пространственном расположении ее частей, что придает раковине в той или иной мере определенную характерную форму. Основные типы строения раковин: неправильного строения (в основном с лучистым или иногда с ветвистым расположением частей), одноосные, спиральные, в том числе неправильно клубковидные, спирально-плоскостные, трохоидные или спирально-конические, спирально-винтовые и правильно клубковидные; циклические раковины представляют собою видоизмененные спиральные.
Закономерное расположение частей раковины отражает в первую очередь закономерности ее онтогенетического, т. е. индивидуального развития. Это последнее идет у фораминифер, как известно, путем последовательного наращивания новых частей.
Иногда, вследствие изменения закономерности нарастания раковины в течение ее индивидуального развития, в одной и той же раковине сочетаются два или более типов строения - образуются так называемые гетероморфные раковины. Так, например, раковина, нарастающая на ранних стадиях по спирали, к концу своего развития образует прямой одноосный отдел и т. п. В одной раковине может сочетаться два, три и более типов строения: биморфные, триморфные и т. д. раковины. Гетероморфность распространяется не только на основные типы строения раковин, но иногда на характер устья, а также и на второстепенные ее особенности, например на число и форму камер по отдельным оборотам, на закономерность возрастания этих последних и т. д.
Тип строения раковины в наибольшей степени отражает специфические, выработавшиеся в филогенезе закономерности совершающихся в протоплазме процессов, которые обусловливают закономерный ход индивидуального развития скелетных образований.
Различные типы строения раковины могут быть охарактеризованы с неизбежной долей схематизации, исходя из общеизвестных геометрических понятий, хотя степень приложимости этих последних к органическим скелетам совершенно иная, чем для геометрически правильных тел неживой природы. Попытка Румблера (1911-1912) установить подобные закономерности в типе строения раковин фораминифер имеет относительную цену, поскольку постоянство угловых величин, как и соотношений размеров, не только подвержено известным отклонениям, но и оказывается переменным в зависимости от стадии индивидуального развития. Необходимо отметить, что Румблер исходил в своих построениях из физического закона постоянства краевых углов для вязких жидкостей, смачивающих определенного рода поверхность. Он ошибочно предполагал, что образование новых камер в индивидуальном развитии фораминифер происходит путем простого истечения протоплазмы из устья на поверхность ранее сформировавшихся частей раковины. К тому же, объяснение особенностей формы камер за счет одних лишь сил поверхностного натяжения оказывается недостаточным, и действительные биологические и физико-химические явления, лежащие в основе морфогенеза раковины, значительно сложнее, чем предполагал Румблер. Несмотря на это, схемы Румблера хорошо иллюстрируют одну из основных особенностей строения раковины, - выраженное в той или иной мере геометрическое подобие слагающих ее последовательных камер (рис. 11). Заслуживает упоминания, что при регенерации поврежденных камер образуется грыжевидный выступ, краевые углы которого оказываются специфическими, теми же, что присущи нормальным камерам данной формы.
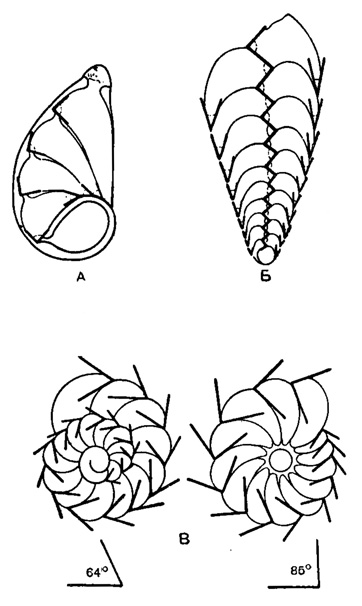
Рис. 11. Схемы, иллюстрирующие 'закон постоянства краевых углов' при образовании раковин различных фораминифер: А - спирально-плоскостная раковина Lenticulina; краевые углы у периферического края больше, чем по краю начальной камеры; Б - спирально-винтовая раковина Bolivitia; краевые углы по краям раковин меньше, чем в осевой ее части; В - спирально-коническая раковина Streblus; краевые углы на спинной стороне меньше, чем на брюшной (соответственно 64° и 85°) (Rhumbler, 1911)
Различные типы строения раковин сочетаются как с агглютинированными, так и с секреционными стенками.
Раковины неправильного строения наблюдаются лишь у некоторых примитивных фораминифер, в частности в сем. Astrorhizidae, Hyperamminidae, а также у ряда форм из других семейств, вторично видоизмененных в результате перехода к сидячему, прикрепленному образу жизни, - некоторые представители сем. Reophacidae, Silicinidae, Polymorphinidae, Ophthalmidiidae, Planorbulinidae, Homotremidae и др. У Astrorhizidae, обладающих раковиной неправильного строения, обычно имеется более или менее хорошо выраженное центральное тело и отходящие от него лучи или руки (например у рода Astrorhiza); лучи раковины в рассматриваемом случае представляют собою скелетный панцирь основания ложноножек. Эти последние выходят наружу через зияющие отверстия на концах лучей.
Неправильные, стелющиеся по грунту амёбовидные по очертаниям раковины наблюдаются у представителей рода Iridia, относимого к сем. Saccamminidae.
У Hyperamminidae, наряду с иного строения раковинами, известны скелетные образования, либо древовидно ветвящиеся, как, например, у Syringammina, либо образованные системой ветвящихся и анастомозирующих трубок. У древовидных форм развивается подчас особая подошва, при помощи которой раковина прикрепляется к субстрату.
Вопрос о том, насколько раковины неправильного строения представителей трех названных семейств являются филогенетически первичными, в собственном смысле слова примитивными, остается в настоящее время неясным. Не исключена возможность, что наиболее примитивные фораминиферы обладали уже раковиной правильного, в частности одноосного, строения. В таком случае все перечисленные примеры относятся к раковинам, вторично приспособившимся к прикрепленному образу жизни.
Одноосные раковины могут рассматриваться в целях наглядности изложения, как тела вращения, приближающиеся то к шару, то к овоиду, то к эллипсоиду, то к цилиндру и т. п. Само собой разумеется, что сказанное имеет чисто геометрический смысл, так как на самом деле нарастание раковины происходит обычно от некоторой начальной точки в направлении вдоль оси (рис. 12).
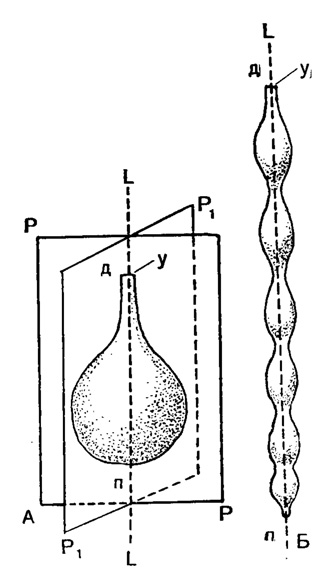
Рис. 12. Схема одноосных раковин: А - однокамерная - типа Lagena; Б - многокамерная - типа Nodosaria; LL - ось симметрии; РР, P1P1 - плоскости симметрии; п - проксимальный конец; д - дистальный конец; у - устье (Фурсенко, 1933)
В простейшем случае, когда любое, перпендикулярное оси сечение раковины - круг, ось симметрии будет бесконечно большого порядка; при повороте "тела вращения" вокруг такой оси на любой произвольный угол будет наблюдаться полное совмещение всех точек тела с их исходным положением.
Нередко раковина одноосного типа строения имеет в своем поперечном сечении эллипс или многоугольник: треугольник, четырехугольник, пяти-шестиугольник и т. д. Соответственно, и ось симметрии таких раковин будет 2-го, 3-го, 4-го, 5-го, 6-го и т. д. порядка (рис. 13, 14). Воображаемая ось симметрии раковины является геометрическим местом точек пересечения плоскостей симметрии раковины; при наличии оси симметрии бесконечно большого порядка, можно представить себе также бесконечное количество плоскостей симметрии.
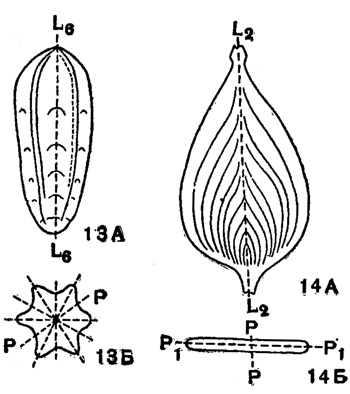
Рис. 13-14. Одноосные раковины. 13 - Nodosaria (шестигранная): А - сбоку; Б - со стороны устья; L6L6 - ось симметрии шестого порядка; РР - одна из шести плоскостей симметрии (Фурсенко). 14. Frondicularia (двусимметричная): А - сбоку; Б - со стороны устья; L2L2 - ось симметрии второго порядка; РР, Р1Р1 - плоскости симметрии (Фурсенко)
Одноосные раковины лишь в исключительных случаях имеют одинаковые окончания. Такие раковины обладают, естественно, еще одной плоскостью симметрии, перпендикулярной в данном случае оси. Поскольку у подобных раковин на обоих противоположных концах расположены отверстия - устья, такие раковины называют двоякоротыми или амфистомными (греч. амфи - дву-, двояко и стома - рот). Двоякоротые, амфистомные раковины, строго говоря, не одноосны, поскольку у них имеется бесконечно большое количество осей симметрии 2-го порядка, расположенных в плоскости симметрии, перпендикулярной продольной оси. В большинстве случаев, однако, одноосные раковины разнополюсны - гетерополярны. Один из концов, с которого начался рост раковины, носит название проксимального; иногда его обозначают как аборальный (противоротовой) или апикальный. Другой конец, удаленный от начала роста, обозначается как дистальный, устьевой или оральный.
В зависимости от характера роста, в одних случаях непрерывного) либо вставочного, либо происходящего путем наращивания новых частей скелета со стороны устья), а в других - прерывистого, ритмичного, одноосные раковины могут быть однокамерными или многокамерными. В промежуточных случаях, при наличии лишь колебаний в скорости роста, на раковине отмечаются только перетяжки или пережимы, обозначающие собою замедления в отложении скелетного вещества. В такой форме в филогенезе ряда групп фораминифер обычно намечается переход от однокамерного строения к многокамерному.
На проксимальном конце многокамерных одноосных форм находится начальная (эмбриональная или примордиальная) камера.
Однокамерные одноосные раковины иногда шаровидны или почти шаровидны; при этом положение оси симметрии определяется подчас лишь по местонахождению устья. В исключительных случаях, когда устье отсутствует, шаровидные раковины, строго говоря, уже не могут рассматриваться как одноосные. В качестве примера можно привести раковины некоторых Saccamminidae, а также раковины представителей пелагического рода Orbulina; у последних, правда, строго сферической и в то же время лишенной устья является только последняя камера, полностью объемлющая все сформировавшиеся ранее и образующие спираль. Чаще, при относительно незначительном удлинении раковины в направлении оси, одноосные раковины обладают яйцевидной (овоидной) или эллипсоидальной формой; нередки раковины в виде плоской фляги (например Entosolenia). При заметном удлинении раковины по оси, она может приобретать веретеновидную, эллиптическую или даже палочковидную или трубчатую форму. Часто раковина заметно расширяется к несущему устье окончанию и приобретает коническую или удлиненно-коническую форму. Благодаря наличию перетяжек многокамерные одноосные раковины могут иметь четковидное строение.
Принято различать длину одноосных раковин, измеряемую в направлении роста, т. е. по оси, их ширину и перпендикулярную последней толщину; толщина и ширина неодинаковы лишь в том случае, если раковина двусимметрична (имеет ось 2-го порядка); тогда под шириной понимается наибольшее из двух измерений, перпендикулярных длине.
Всякое сечение, ориентированное в направлении оси раковины, будет продольным. Продольное сечение называется осевым (аксиальным), если оно проходит строго по оси раковины. Близкие к поверхности раковины продольные сечения носят название тангенциальных. Сечения, перпендикулярные оси, обозначают как поперечные. Сечения, ориентированные по отношению к оси раковины под некоторым произвольным углом, не равным 0° или 90°, принято называть косыми.
Одноосные раковины присущи в основном донным - бентосным фораминиферам, приспособленным к неподвижно лежачему или ползающему образу жизни. Исключение представляют планктонные Orbulina, шаровидные раковины которых, как это уже отмечалось не являются в строгом смысле одноосными.
В результате наблюдающегося иногда перехода к прикрепленному образу жизни одноосные формы приобретают неправильное строение и образуют раковину, змеевидно стелющуюся по субстрату. В качестве примера можно привести некоторых Reophacidae, род Placopsilina и ему близкие и другие. У подобных прикрепленных форм обращенная к субстрату сторона становится плоской, а поперечное сечение раковины - приближающимся к полукругу.
Спиральные раковины можно представить как результат видоизменения одноосных в силу скручивания оси. Такое определение имеет не только геометрический смысл и удобно в целях наглядности, но и в известной мере отвечает действительности, поскольку в процессе филогенетического развития фораминифер спиральные раковины происходят от одноосных. Бывает, правда, в некоторых случаях и обратный переход: от спиральных форм к одноосным. При этом спиральная раковина сперва образует более рыхло свернутый конечный отдел, затем в процессе эволюции происходит дальнейшее раскручивание этого отдела. Принято различать: неправильно клубковидные раковины, когда навивание происходит в беспорядке или в некоторой более или менее непрерывно меняющей свое положение плоскости; раковины спирально-плоскостные, спирально-конические (трохоидные) и спирально-винтовые, являющиеся по существу лишь крайним выражением спирально-конического типа строения. Особо следует выделять раковины сем. Miliolidae, отличающиеся в типичном случае правильно клубковидным навиванием. У милиолид наблюдается навивание и по спирально-плоскостному типу.
Неправильно клубковидные раковины по своему строению наиболее примитивны; их можно представить как результат беспорядочного навивания одноосного скелетного образования в нескольких направлениях. Подобные раковины, по-видимому, и генетически, наиболее примитивны, поскольку наблюдаются в таком относительно низко организованном и в то же время древнем семействе, как Ammodiscidae (пример - род Glomospira). У некоторых представителей названного рода наблюдается, однако, известный переход к более правильному расположению оборотов - спирально-винтовому, как у Gl. charoides, или к охватывающему часть клубковидных оборотов спирально-плоскостному типу.
Особенно характерны неправильно клубковидные раковины для сем. Endothyridae, где наблюдаются и многочисленные случаи выраженного в более или менее чистом виде спирально-плоскостного типа строения. Неправильно клубковидное навивание многокамерных раковин представителей этого семейства послужило основанием для введения термина плектогироидное навивание (от названия Pledogyra). Плектогироидное, или, как его называли прежде, эндотироидное, навивание наблюдается и в начальных оборотах некоторых представителей отр. Fusulinida, что рассматривается как дополнительное указание на происхождение этого семейства, вообще говоря обладающего спирально-плоскостными раковинами, от эндотирид.
Спирально-плоскостными раковинами принято называть такие, спиральная ось которых расположена в одной плоскости; они являются двусторонне-симметричными. Термины плоскоспиральная, планоспиральная или наутилоидная менее удачны, так как спиральноплоскостные раковины не всегда плоские. Раковины этого типа строения могут быть как многокамерными, так и двухкамерными, состоящими из шаровидной начальной камеры и второй трубчатой спиральной.
Плоскость, в которой располагается воображаемая спиральная ось, является одновременно и плоскостью симметрии раковины (рис. 15); это так называемая срединная, медианная или экваториальная плоскость. Располагающиеся по сторонам экваториальной плоскости симметричные половины раковины носят название боковых (латеральных) сторон раковины. Часть раковины вблизи срединной области называется срединной. Перпендикуляр к экваториальной плоскости, проходящий через центр спирали, принято обозначать как ось навивания (завивания) или просто ось раковины (не смешивать со спиральной осью раковины). Область вблизи точек выхода оси навивания называется осевой, или пупочной. Отрезок спирали, конечная точка которого удалена от начальной на 360°, носит название оборота. В противоположность центральным частям раковины, наиболее удаленные носят название периферических.
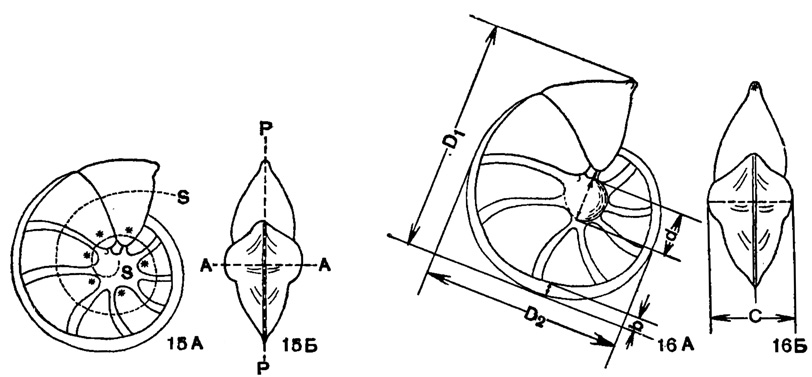
Рис. 15-16. Схема соотношения частей и элементов симметрии и измерений спиральноплоскостной раковины Lenticulitia: 15 - Элементы симметрии: А - вид сбоку; Б - вид с периферического края; АА - ось навивания; РР - плоскость симметрии (экваториальная или медиальная); SS - спиральная ось; * - пупочные окончания камер, ограничивающие пупочную область раковины (Фурсенко). 16. Схема измерений: А - вид сбоку; Б - вид с периферического края; d - диаметр пупочного диска (ограничен окончаниями стрелок); D1 - больший диаметр; D2 - малый диаметр; С - толщина раковины; b - ширина киля (ограничена окончаниями стрелок) (Фурсенко)
При изучении раковин фораминифер спирально-плоскостного типа строения практически приходится иметь дело не с воображаемой спиральной осью, а с внешними очертаниями последовательных оборотов, лишь в некоторой мере отражающими направление спиральной оси. Внешний, видимый при разглядывании раковины сбоку контур наружного оборота носит название внешнего, или периферического, края. Характер этого края - существенный диагностический признак при определении и при описании спирально-плоскостных (а также и спирально-конических) форм. Периферический край при рассматривании раковины сбоку может быть ровным или лопастным. В сечении он бывает закругленным, округло-усеченным, усеченным, заостренным, острым, килеватым (с одним, двумя или тремя килями).
При изучении в шлифах нуммулитов приходится обращать внимание не на внешний контур оборотов, а на сечение периферической стенки оборотов, имеющей известную толщину. На экваториальных сечениях эта стенка выражена в виде так называемой спиральной полосы.
В большинстве случаев спирально-плоскостные раковины образованы несколькими оборотами, но иногда имеется лишь один полный или даже неполный оборот (например раковины некоторых Lagenidae - Lenticulina, Dentalina).
Наибольший поперечник спирально-плоскостной раковины, лежащий в ее срединной плоскости, называется наибольшим диаметром или просто диаметром. Чтобы правильно провести диаметр, необходимо мысленно наметить две точки: центр спирали и точку, находящуюся на наибольшем удалении от центра на периферическом крае, т. е. на окончании спирали. Через эти две точки следует провести прямую до пересечения с внешним краем на противоположной стороне раковины (рис. 16). В некоторых случаях полезно ввести также понятие малого диаметра или поперечника - перпендикуляра к наибольшему диаметру в плоскости симметрии раковины. Отношение наибольшего диаметра к малому дает представление о степени удлиненности раковины. Отрезки диаметра, отвечающие размерам отдельных оборотов, носят название высот оборотов. Под толщиной спирально-плоскостной раковины принято понимать измерение по оси навивания. У вытянутых в направлении этой оси раковин из отр. Fusulinida для подобного измерения употребляется наименование - длина; это наименование распространяется обычно и на сжатые по оси (линзовидные, шаровидные и т. п.) раковины данного отряда.
Необходимо указать, что по своей форме раковины спирально-плоскостного типа строения в достаточной мере изменчивы: у одних родов или видов они дисковидные; в таком случае боковые стороны оказываются более или менее плоскими, толщина - незначительной. У линзовидных или чечевицеобразных раковин боковые стороны - низкие конусы; диаметр раковины превышает по своим размерам толщину (рис. 17). Встречаются шаровидные или субсферические (почти шаровидные) раковины. У веретеновидных раковин толщина (в данном случае длина) превышает диаметр, а сама раковина оттянута к концам. Сильно вытянутые по оси раковины могут быть почти цилиндрическими или субцилиндрическими - вальковидными, как это имеет место у некоторых высокоорганизованных представителей отр. Fusulinida.
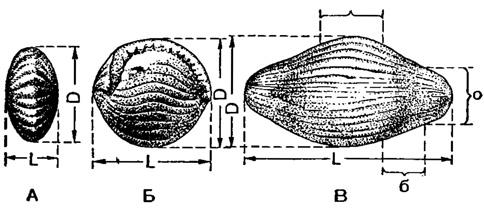
Рис. 17. Спирально-плоскостные раковины фузулинид: А - Staffella (чечевицеобразная); Б - Schwagerina (шаровидная); В - Fusulinella (веретеновидная); DD - диаметр; LL - длина раковины в направлении осн; С - срединная область; б - бока раковины; о - осевые или полюсные концы (Раузерчерноусова, 1937, по Меллеру, 1880)
Степень удлиненности раковины в направлении оси имеет для Fusulinida важное систематическое значение и определяется отношением длины (практически иногда - толщины) раковины к ее диаметру (L:D). Систематическое значение имеет также характер изменения в отношении длины к диаметру в процессе индивидуального развития.
На общей форме спирально-плоскостных раковин сильно сказывается степень возрастания высоты последовательных оборотов. При незначительном возрастании высоты раковины приближаются по своим очертаниям к кругу; наоборот, при резком возрастании раковина приближается по форме к рогу изобилия. Между этими крайними случаями имеются всевозможные переходы.
При описании спирально-плоскостных раковин, удлиненных в направлении диаметра, приходится вводить понятия о спинном и брюшном краях. Понятия "спинной" и "брюшной" у спирально-плоскостных форм имеет совершенно иное значение, чем у спирально-конических (у них - спинная и брюшная стороны).
При наличии нескольких оборотов они могут либо просто налегать один на другой; в таком случае снаружи видны все обороты, называемые необъемлющими. Раковины с необъемлющими оборотами называются эволютными. Нередко каждый последующий оборот полностью охватывает с боков предыдущий; обороты такого типа называют объемлющими, а раковины с объемлющими оборотами - инволютными. При изучении нуммулинтов термины эволютный и инволютный употребляются применительно к оборотам и даже содержащимся в этих последних полостям камер.
Видимая снаружи часть завитка носит название пупка раковины. У эволютных форм снаружи видна большая часть завитка - пупок у них широкий. У инволютных он может быть либо узким, либо вовсе отсутствовать. Пупок может быть открытым или, наоборот, заполненным веществом дополнительного скелета, кроме того, глубоким или неглубоким.
Наименования сечений спирально-плоскостных раковин в основном те же, что и для одноосных, причем учитывается ориентировка сечения по отношению к оси навивания; присущее лишь спиральноплоскостным раковинам сечение, отвечающее плоскости симметрии, носит название экваториального, или медианного.
Спирально-конические, или трохоидные (от названия моллюска Trochus), раковины в большинстве случаев многокамерные, но известны двухкамерные - трубчатые.
Спирально-конические раковины отличаются от спирально-плоскостных тем, что их спиральная ось расположена не в одной плоскости, а на некоторой воображаемой конической поверхности. Отношение высоты конуса к его основанию может быть различным и достигать 1:1. Раковины, у которых это отношение превышает единицу, по существу также спирально-конические, но их принято выделять в особую группу спирально-винтовых. Спирально-конические раковины совершенно несимметричны; этим они отличаются от спирально-плоскостных. Понятие оси навивания вполне, однако, применимо и в данном случае. Стороны спирально-конических раковин не могут быть равноценными, как у спирально плоскостных. Обычна одна сторона более выпуклая, другая-менее. Однако иногда, при наличии спирально-конического строения и несимметричном строении сторон, раковина оказывается двояковыпуклой и внешне симметричной: расположение спиральной оси оказывается замаскированным.
Принято называть спинной (дорзальной) или спиральной стороной ту, на которой снаружи видны все обороты спирали или, во всяком случае, большее их количество, чем на другой стороне; обороты на этой стороне не объемлющие или же менее объемлющие. Брюшной (вентральной), или пупочной, стороной спирально-конических раковин называют ту, где, напротив, снаружи виден лишь один последний оборот или меньшее число оборотов (рис. 18). У одних родов или даже видов выпуклее может быть спинная сторона, а у других - брюшная. У ползающих по грунту фораминифер со спирально-конической раковиной более плоской бывает обычно брюшная сторона. У других форм наблюдаются обратные отношения; такой особенностью отличаются преимущественно неподвижно лежащие на грунте или прирастающие к субстрату. С прикреплением к субстрату спинной стороной в филогенезе фораминифер происходит постепенная утрата правильного спирально-конического расположения частей - камер раковины (Planorbulinidae, Homotremidae). Спирально-коническое, унаследованное от предков строение сохраняется у таких форм на ранних стадиях индивидуального развития. В процессе развития спирально-конических известковистых фораминифер появляются, кроме того, формы, приспособленные к планктонному образу жизни - к свободному парению в толще воды (сем. Globigerinidae и Globorotaliidae). У этих форм выработались своеобразные особенности строения: шаровидные или килеватые камеры, облегченная стенка с ячеистой поверхностью (рис. 19).
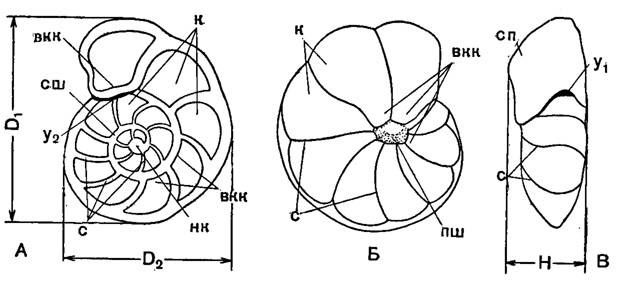
Рис. 18. Схема строения спирально-конической раковины Anomalina: А - со спинной (спиральной) стороны; Б - с брюшной (пупочной) стороны; В - с периферического края; D1 - наибольший диаметр; D2 - меньший диаметр; H - высота (толщина) раковины; вкк - внутренние концы камер; к - камеры; нк - начальная камера; пш - пупочная шишка; с - септальные швы; сп - септальная поверхность; сш - спиральный шов; y1-y2 - устье (Василенко, 1954)
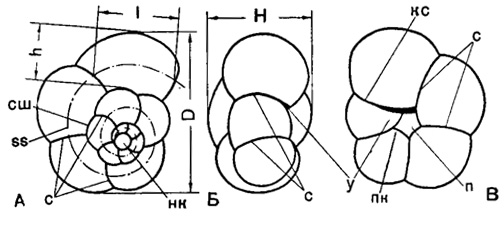
Рис. 19. Схема строения спирально-конической раковины Globigerinidae: А - со спинной (спиральной) стороны; Б - с периферического края; В - с брюшной (пупочной) стороны; D - диаметр (наибольший); H - высота; п - пупок; у - устье; h - ширина (высота) камеры; l - длина камеры; ss - спиральная ось раковины; кс - краевой септальный шов; нк - начальная камера; пк - пупочное окончание камеры; с - септальные швы; сш - спиральный шов (Субботина, 1953)
Основные измерения спирально-конических форм те же, что спирально-плоскостных; высотой называется измерение в направлении оси навивания. Наименования сечений также сходные, но сечения, перпендикулярные оси, у спирально-конических раковин носят название спиральных.
Спирально-винтовые раковины в подавляющем большинстве случаев многокамерные. Высота или длина раковины превышает основание конуса (ширину, толщину) раковины (рис. 20).
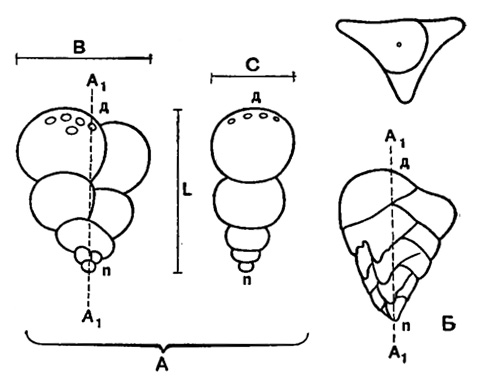
Рис. 20. Схема строения спирально-винтовых раковин: А - Cribrostomum (двурядная); Б - Tritaxia (трехрядная); A1A1 - ось навивания; L - длина; В - ширина; С - толщина; п - проксимальный конец; д - дистальный конец
В зависимости от числа камер, образующих отдельные обороты, различают двухрядные, трехрядные, четырехрядные, пятирядные спирально-винтовые раковины. Число камер в обороте в данном типе строения лишь в исключительных случаях превышает пять. Взаимное расположение камер в оборотах спирально-винтовых форм можно определить также из того угла, который образуют отдельные камеры одна по отношению к другой. Так, в двухрядных раковинах камеры ориентированы под углом 180°, а в трехрядных располагаются одна по отношению к другой под углом 120° и т. д.
В онтогенезе спирально-винтовых форм количество камер по оборотам может меняться: гетероморфная раковина, начавшая свое развитие как трехрядная, может на более поздней стадии стать двухрядной (например Gaudryina) и т. д. Спирально-винтовое строение нередко сочетается в одной и той же раковине и с другими типами строения - одноосным или спирально-плоскостным.
Камеры спирально-винтовых раковин могут прилегать друг к другу, либо, как, например, у Polymorphinidae, охватывать одна другую. У некоторых представителей этого семейства, камеры настолько сильно заходят своими проксимальными окончаниями к одноименному концу раковины, облекая при этом предыдущие, что почти охватывают начальную камеру. Для раковин двухрядного строения принято различать, боковые стороны, с которых видны все камеры - оба их ряда, и край раковины, или периферический край, со стороны которого, естественно, можно видеть только камеры одного из рядов. Для трехрядных раковин и раковин, обороты которых образованы числом камер большим трех, приходится говорить лишь о боковых сторонах раковины, соответствующих по существу всей, ее поверхности, за исключением дистальных поверхностей последних камер.
При измерениях спирально-винтовых раковин принято называть длиной расстояние по оси от проксимального до дистального конца, шириной - наибольший поперечник раковины, перпендикулярный оси. Особое понятие толщины приходится вводить только при изучении двухрядных раковин; для них ширина - измерение в той плоскости, в которой видны два ряда камер (эта же плоскость является плоскостью симметрии); толщина двухрядных раковин - измерение в направлении, перпендикулярном к названной плоскости, и к оси раковины.
Кроме общепринятых сечений для двухрядных раковин приходится различать среди осевых сечений сагиттальное, проходящее в плоскости симметрии через оба ряда камер, и фронтальное, проходящее также строго по оси раковины, как и сагиттальное, но ориентированное к этому последнему под прямым углом.
Правильно клубковидные раковины сем. Miliolidae по типу своего строения стоят несколько особняком. Камеры, составляющие раковину этих фораминифер, располагаются в наиболее типичных случаях в нескольких взаимно пересекающихся плоскостях, образуя свернутый клубок; при этом обычно каждая камера занимает половину оборота (см. рис. 327). Ось навивания раковин Miliolidae закономерно меняет свое положение в плоскости, называемой главным сечением. Обычно это сечение перпендикулярно наибольшему измерению раковины - ее длине - и совпадает с направлениями, которые принято обозначать, как ширина и толщина.
Необходимо заметить, что у некоторых Miliolidae, таких, как представители родов Spiroloculina и Pyrgo (Biloculina), раковины имеют спирально-плоскостное строение. В главном сечении видно, что камеры располагаются одна по отношению к другой, под углом в 180°, т. е. лежат в одной плоскости (рис. 324). У некоторых Miliolidae наблюдаются раковины гетероморфного строения. Они сочетают в себе два или более типов строения, отвечающих различным стадиям индивидуального - онтогенетического развития.
Наиболее характерен для Miliolidae так называемый пятерной (квинквелокулиновый - от названия рода Quinqueloculitia) тип строения раковины. В главном сечении у таких форм наблюдается следующая картина (рис. 325). По отношению к расположенной в центре раковины начальной камере первая камера будет располагаться в плоскости, условно принятой за нулевую, вторая, при отсчете угловых расстояний против часовой стрелки - в плоскости, ориентированной под углом 144°, третья - под тем же углом по отношению ко второй, а стало быть, в плоскости, ориентированной под углом 288° к первой камере. Нетрудно заметить, что если вести отсчет по часовой стрелке, третья камера по отношению к первой располагается под углом 360° - 288° = 72°. Продолжая счет против часовой стрелки, четвертая по порядку возникновения камера располагается под углом 144° по отношению к третьей, т. е. под углом 72° по отношению к первой, попадая посредине между первой и второй. Пятая камера, располагаясь под углом в 144° по отношению к четвертой и подобно этой последней, попадает в промежуток между двумя камерами, в данном случае - второй и третьей. Наконец, шестая по порядку возникновения камера оказывается расположенной в той же нулевой плоскости, что и первая. При последующем нарастании камер картина повторяется: седьмая камера располагается в плоскости второй, восьмая - третьей и т. д.
Для пятерного - квинкелокулинового - строения характерно расположение последовательных по порядку возникновения камер под углом в 144°, а смежных по расположению под углом в 72°. Благодаря такой закономерности раковины подобного типа строения в известной мере приближаются по своей форме к многограннику, правда, неправильному, благодаря прогрессивному возрастанию размеров последовательных камер. У наиболее типичных представителей рода Quinqueloculitia снаружи бывает видно пять камер, при этом с одного бока - четыре, а с другого три (рис. 328, А, Б), из-за того, что две последние камеры видны и в том и в другом положении.
В некоторых случаях пятерное расположение камер нарушается на более или менее ранней стадии онтогенеза раковины - камеры продолжают нарастать под углом в 120°; раковина в результате этого приобретает трехгранное - тройное или трилокулиновое строение. Формы с подобным расположением камер прежде относят к особому роду Triloculina (рис. 326).
У представителей рода Sigmoilina нарастание последовательных камер происходит в плоскостях, расположенных одна по отношению к другой под углами, большими 144° и меньшими 180°, в результате чего в главном сечении получается характерная S-образная фигура, откуда и название Sigmoilina (рис. 327).
У раковин Miliolidae различают устьевой (апертурный) конец, на котором находится устье последней камеры раковины, основание раковины - ее противоположный конец - и стороны: многокамерную, где у Quinqueloculina видно снаружи четыре камеры, или, вообще говоря, большее число камер, и малокамерную, где у Quinqueloculina видно три камеры, или вообще меньшее число камер. Кроме концов и сторон раковины, у Miliolidae в большинстве случаев удобно различать периферический край, разграничивающий стороны.
Основные направления измерений раковин Miliolidae: длина - измерение от основания раковины до ее устьевого конца; ширина - наибольшее измерение между наиболее удаленными точками периферического края в перпендикулярном длине направлении; толщина - наибольшее расстояние между наиболее выдающимися точками малокамерной и многокамерной сторон или вообще наименьший поперечник.
Циклические раковины с камерами, расположенными по концентрическим окружностям в одной плоскости или по некоторой воображаемой конической поверхности, наблюдаются у фораминифер относительно редко. При этом для большинства родов, отличающихся подобным строением, установлено, что начальный отдел раковины имеет спиральное строение; особенно часто проявляется эта особенность у форм микросферического поколения. Исходя из допущения рекапитуляции, можно рассматривать циклический тип строения как результат видоизменения спирального.
Циклическое строение наиболее ярко выражено у представителей сем. Orbitiodidae и ему близких. Их раковина, имеющая в большинстве случаев дисковидную или чечевицеобразную форму, заключает в центре начальную камеру, обнаруживающую сложное строение: обычно она бывает поделена на несколько вторичных камерок. Стенка начальной камеры, как правило, толстая, значительно толще перегородок между вторичными камерками. В данном случае скорее можно говорить о начальной раковине, чем о начальной камере, и эта по существу эмбриональная раковина обозначается нередко как нуклеоконх. По предположению Ле Кальве (1953), нуклеоконх представляет собою раковину зародыша - мерозоита, образованную им еще во время пребывания внутри материнского шизонта; последующие, характерные для Orbitoididae, Discocyclinidae и Miogypsinidae камеры образуются, по мнению этого исследователя, уже после того, как зародыш перешел к свободному образу жизни. При этом экваториальные камеры располагаются в один, реже в два слоя концентрическими рядами в средней плоскости раковины и образуют в своей совокупности экваториальную пластинку. Обычно экваториальная пластинка однослойная (Orbitoides, Discocyclinidae и др.), иногда в периферической зоне наблюдается ее удвоение, а между образовавшимися двумя слоями может в еще более периферических частях вклиниваться третий (Omphalocyclus).
Помимо нуклеоконха и камер экваториальной пластинки, у большинства Orbitoididae, Discocyclinidae и др., за исключением Omphalocyclus, имеются еще многочисленные так называемые латеральные (боковые) камеры, образующие как бы подушки по сторонам экваториальной пластинки.
Расположение латеральных камер либо приближается к циклическому, либо более или менее беспорядочное.
Несколько иначе построены циклические раковины у некоторых представителей сем. Peneroplidae.(Orbitolites и близкие к нему роды). Здесь вслед за обычного типа начальной камерой располагаются камеры спирального отдела, не выраженного обычно у мегасферических форм. Благодаря непомерному возрастанию оборотов спирали в высоту, последующие камеры приобретают кольцевую форму. Дальнейшее нарастание новых камер происходит циклически. Характерной особенностью таких форм является наличие в камерах вторичных радиальных перегородок, обеспечивающих подразделение основных камер раковины на вторичные камерки.
Устье. Простым устьем (апертурой) называется отверстие, при помощи которого внутренние полости раковин фораминифер сообщаются с внешней средой. Устье называется сложным или множественным, если это сообщение осуществляется через несколько отверстий. При образовании новой камеры раковины устье предыдущей камеры становится отверстием, соединяющим полости этих двух камер между собой, - устье превращается в так называемый форамен (откуда и название Foraminifera - буквально от латинского foramen - отверстие, дыра; дырочники). У некоторых фораминифер (Epistominidae) при образовании новой камеры первичное устье - протофорамен (Hofker, 1950, 1951) - зарастает, а сообщение между последовательными камерами осуществляется при помощи вторичного, вновь образовавшегося отверстия - дейтерофорамена.
По своему положению устье может быть конечным (терминальным), например у одноосных форм, когда оно располагается на дистальном окончании раковины (рис. 21). Оно может быть основным (базальным), если находится у основания септальной поверхности последней камеры и при этом у внутреннего края этой поверхности (интерно-маргинальная апертура, Hofker, 1953). У спирально-плоскостных форм устье при этом занимает, как правило, срединное (медианное) положение. Основное, или базальное, устье при трохоидном строении раковины может оказаться срединным, спинным или брюшным; наблюдаются и другие случаи, например устье срединное, но заходящее на спинную или брюшную сторону. Иногда устье может быть расположено на самой септальной поверхности, посреди нее или ближе к одному из ее краев (ареальная апертура, Hofker, 1950, 1951). Наконец (это касается лишь простого устья спирально-плоскостных и трохоидных форм), оно может располагаться у внешнего края септальной поверхности на периферическом крае раковины - периферическое устье (некоторые Lagenidae, Epistomina).
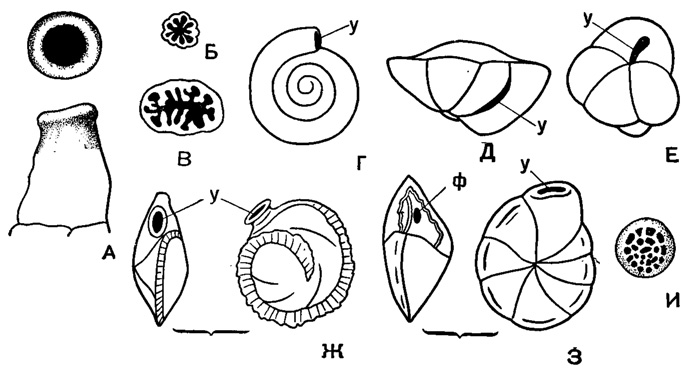
Рис. 21. Различные типы устья у фораминифер: А - конечное круглое, отороченное губой; Б - расчлененное на лопасти; В - расчлененное на разветвленные лопасти (приближающееся к древовидному); Г - конечное у спирально-плоскостной формы (зияющее отверстие трубчатой камеры); Д - основное (базальное, интерно-маргинальное) устье; Е - петлевидное; Ж - устье, расположенное на септальной поверхности (ареальное), на конце шейки, отороченной губой; З - периферическое щелевидное устье; у - (протофорамен); ф - форамен (дейтерофорамеи) - отверстие, при помощи которого сообщаются полости последовательных камер; на стенках камер (кроме последней) видны зарубцевавшиеся устья (протофорамены); и - сложное - ситовидное устье (Богданович, 1953; Мятлюк, 1953)
По своей форме простое устье может быть круглым, почти круглым (округлым),эллиптическим, щелевидным, полулунным, серповидным, петлевидным. Благодаря образованию неровностей по краям устья, оно может быть лопастным; в подразделении на лопасти могут играть роль выросты различного строения, как, например, у Ataxophragmiidae, Miliolidae и др. У некоторых фораминифер из сем. Peneroplidae устье древовидное - сложно разветвленная щель. У большинства представителей отр. Lagenida устье лучистое - круглое отверстие, осложненное радиально отходящими от него щелями. Строение устья усложняется особыми связанными с ним образованиями. У форм с конечным устьем нередко, например, наблюдается устьевая шейка в виде короткой трубочки, подчас отороченной губой; иногда такая шейка вместо того, чтобы выдаваться наружу, оказывается ввернутой внутрь полости раковины или последней ее камеры. У некоторых Lagenidae и Polymorphinidae устье открывается не непосредственно в полость последней камеры, а в особую устьевую камерку; последняя, в свою очередь, при помощи простого круглого отверстия открывается в полость камеры. У ряда фораминифер (Epistominidae, Ceratobuliminidae, Buliminidae и др.) от края устья в полости камеры отходит особая зубная пластинка (Hofker, 1950, 1951; Hoglund, 1947), часто желобчато изогнутая; внутренним своим концом эта пластинка прирастает обычно к краю устья (или дейтерофорамена) предшествующей камеры. У форм с первичным зарубцовывающимся впоследствии устьем (протофораменом) зубная пластинка скрепляет края того и другого отверстия. Как самый факт наличия зубной пластинки, так и ее характер, форма, способ прикрепления - важные, систематические признаки. У некоторых фораминифер по краю устья, внутри него, развиваются различного строения выросты - зубы (в частности у Miliolidae и др.). Устьевой зуб может быть палочкообразным, пластинчатым, двухраздельным, кольцевидным; в последнем случае устье неизбежно становится сложным, состоящим из нескольких отверстий; дальнейшее расчленение зуба может привести к образованию ситовидного устья (рис. 22). Иногда, при наличии по сути дела простого устья, оно оказывается осложненным наличием добавочного отверстия. Так, например, у рода Robulus, помимо лучистого периферического устья, имеется еще щелевидное отверстие на септальной поверхности.
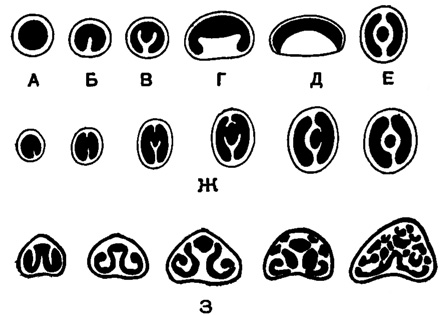
Рис. 22. Различное развитие устьевого зуба у Miliolidae: А-Е - зубы различных видов Miliolidae: (А - беззубая форма); Ж - развитие кольцевидного устья в онтогенезе; З - развитие ситовидного устья в онтогенезе (Богданович, 1953)
Сложное устье чаще всего бывает ситовидным, состоящим из ряда небольших отверстий, расположенных на септальной поверхности, иногда на конце одноосной раковины - терминально. У некоторых форм (Nonionidae, Elphidiidae) сложное устье, состоящее из нескольких отверстий, расположено базально - у основания септальной поверхности. Ситовидный характер устья у агглютинированных форм нередко сочетается с губчатым строением стенки.
В онтогенезе устье может менять свое положение: устье ранних камер может быть, например, базальным, а в дальнейшем ареальным - смещаться на септальную поверхность, или вначале простым, а затем ситовидным и т. п. Устьевые зубы в процессе онтогенеза в ряде последовательных камер могут претерпеть изменения от простого палочковидного, через ряд промежуточных стадий; (два палочковидных, ориентированных навстречу один другому, двураздельный и палочковидный, два встречных двухраздельных) до кольцевого и т. п.
Различные усложняющие строение устья морфологические структуры типа зубов, пластинок и г. п. играют, по-видимому, важную роль в передвижении фораминифер, обеспечивая прикрепление основания выходящего из устья пучка ложноножек; кроме того, все эти образования увеличивают прочность раковины в наиболее подверженной излому ее части.
Не выяснено физиологическое значение свойственных некоторым фораминиферам прикрывающих устье пластинок, имеющих характер отворота или козырька края устья (Globigerinidae, Globorotaliidae). У Janischewskina устье сложное, причем каждое из составляющих его отверстий полностью прикрыто круглым козырьком, прирастающим одной своей стороной к стенке раковины у края устья.
Система каналов. Наличие системы каналов, находящихся внутри раковины, является особенностью наиболее высоко организованных фораминифер из сем. Rotaliidae, Calcarinidae, Elphidiidae и Nummulitidae. Система каналов у представителей названных семейств была изучена первоначально Карпентером и др. (Carpenter, Parker and Jones, 1862), относительно недавно Хофкером (Hofker, 1927, 1930), а в самое последнее время В. А. Крашенинниковым (1956).
У Rotalia polystomelloides Parker et Jones (рис. 23) система каналов имеет следующее строение: основными ее элементами являются спиральный и межсептальный (внутрисептальный) каналы. Спиральный канал один; он постепенно увеличивается в диаметре по направлению к более поздним оборотам, завернут в глубине брюшной части раковины в роталоидную - коническую спираль; начинается он слепо у начальных камер раковины и открывается небольшим отверстием на ее поверхности близ устья с нижней стороны последней камеры. Этот канал имеет, кроме того, сообщение с брюшной (пупочной) лопастью каждой из камер раковины. В промежутках между камерами спиральный канал дает ответвления - межсептальные каналы, которые, в свою очередь, подразделяются на четыре-пять расходящихся веерообразно канальцев, открывающихся в швах; отдельные отверстия этих канальцев разграничены друг от друга известковыми перемычками.
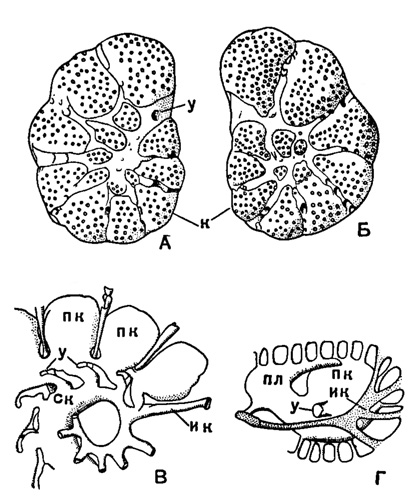
Рис. 23. Rotalia polystomelloides Parker et Jones. Система каналов: А - брюшная (пупочная) сторона; Б - спинная (спиральная) сторона; В - отливка внутренних полостей камер и каналов, в экваториальном плане; Г - то же, в акснальном плане; ик - интрасептальный (внутрисептальный) канал; к - камеры; пк - полости камер; пл - пупочная лопасть; ск - спиральный канал; у - устье (форамен) × 60 (Hofker, 1930, из Sigal, 1952)
У представителей семейства калькаринид система каналов изменена в связи с развитием вторичных наслоений дополнительного скелета, в том числе характерных для данного семейства радиальных шипов. Имеется сеть анастомозирующих пупочных каналов, пронизывающих пупочный диск и открывающихся наружу посреди брюшной поверхности. От этой сети каналов отходят ответвления, при помощи которых каналы сообщаются с брюшной частью камер; кроме того, в направлении к краю раковины и к ее спинной поверхности отходят многочисленные радиальные каналы, разветвляющиеся и открывающиеся на спинной поверхности. Шипы Calcarina также пронизаны каналами. У нонионид система каналов присутствует не всегда; ее нет у наиболее примитивных форм с незаполненным пупком и плоскими (поверхностными) швами, таких, как Melonis pompiliodes (Fichtel et Moll). У форм, подобных Florilus asterizans (Fichtel et Moll) с пупком, заполненным вторичными образованиями, и с углубленными швами, система каналов имеется, но строение ее несложно: с каждой стороны симметричной спирально-плоскостной раковины наблюдается по спиральному каналу, затем имеются пупочные и короткие межсептальные каналы, едва выходящие за пределы пупочной области.
У представителей семейства эльфидиид система каналов имеет более сложное строение. Так, у рода Elphidium наблюдается два спиральных канала, имеющих вид низких конусов, обращенных вершинами друг к другу. От этих каналов к пупку отходят не образующие анастомоз пупочные каналы. Межсептальные каналы имеются на каждой из сторон раковины; у периферии они соединяются попарно. Межсептальные каналы дают, кроме того, боковые ответвления, открывающиеся в области швов септальными порами; у микросферических форм эти ответвления, в свою очередь, раздваиваются, в результате чего образуются парные септальные поры.
У представителей нуммулитид система каналов имеет различное строение. Так у Nummulites она представлена спиральным каналом, проходящим под периферическим краем раковины, и межсептальными разветвленными каналами, находящимися между двумя скелетными листками, образующими двойную септу. У Operculina, по данным Карпентера, периферический канал в более поздних оборотах подразделяется на сеть канальцев, заполняющую всю периферическую область стенки камеры. У Heterostegina, система межсептальных (интерсептальных) каналов отсутствует, а взамен нее развивается сетка каналов, окружающих каждую вторичную камерку.
Швы. Последовательные камеры раковин (у многокамерных форм) отделяются одна от другой септальными швами, которые отвечают линиям сочленения септ с наружной стенкой раковины. Линия соприкосновения смежных оборотов у спиральных форм называется спиральным швом.. Швы могут быть в той или иной мере углубленными (при вздутых в различной степени камерах), поверхностными или выпуклыми. Швы бывают либо линейными, в виде не имеющей практически толщины границы между смежными камерами, либо каемчатыми, обозначенными на поверхности раковины как двухконтурные полоски, либо натечными, когда вдоль них отлагается, дополнительное скелетное вещество. По своим, очертаниям септальные швы могут быть прямыми, дуговидно или сигмоидально изогнутыми и т. п. У Nummulitidae септальные швы - в данном случае так называемые "следы перегородок" - могут быть радиальными и при этом прямыми или дуговидно изогнутыми, волнистыми, вихреобразно закрученными, меандрирующими и, наконец, сетчатыми (благодаря слиянию окончаний смежных септ). У одноосных форм по отношению к оси симметрии или к периферическому краю у спиральных форм швы могут быть ориентированы перпендикулярно или наклонно под различным, характерным обычно для того или иного вида углом. У некоторых форм (Elphidiidae) швы бывают осложнены более или менее многочисленными поперечными скелетными перемычками, соединяющими выпуклые поверхности смежных камер: это либо плотные септальные, либо имеющие полость межсептальные мостики, в которые заходят особые так называемые ретральные отростки протоплазмы. В промежутках между мостиками расположены, соответственно, септальные или межсептальные ямки. Так называемые пресептальные и постсептальные пластинки сем. Bradyinidae располагаются у швов, но внутри - в полости камер, эти пластинки являются образованиями, увеличивающими прочность пришовных областей раковины.
Характер швов имеет, как правило, значение видового или родового признака.
Дополнительный скелет. Под дополнительным скелетом у фораминифер понимаются все те образования, которые не представляют собою ни стенок раковины, ни перегородок между камерами, т. е. основных частей скелета. Дополнительный скелет образуется в онтогенезе, по-видимому, несколько позже основного скелета, в качестве вторичных отложений. Дополнительный скелет может быть как внутренним, так и наружным. К образованиям внутреннего дополнительного скелета относятся различные базальные и иные образования у Endothyrida и в частности хоматы у некоторых Fusulinida; парахоматы, псевдохоматы, осевые уплотнения и т. д. у различных Fusulinida;
К образованиям наружного дополнительного скелета относятся различные известковые натеки на поверхности раковины, ребра, шипы, иглы. Наблюдающиеся у ряда фораминифер грануляции - бугорки - представляют собою обычно выходы на поверхность стенки окончаний столбиков дополнительного скелета, пронизывающих раковину и имеющих обычно коническую форму (Nummulitidae, Discocyclinidae).
У многих спиральных фораминифер дополнительные скелетные образования выполняют пупочную область, образуя либо известковую втулку - затычку, более или менее свободно сидящую в пупочном углублении, либо так называемые пупочные диски, являющиеся, строго говоря, выходами на поверхность раковины конических по форме выполнений пупочных углублений (некоторые Lagenidae, Rotaliidae, Elphidiidae, Nummulitidae и др.). Нередко пупочные диски бывают пронизаны поровыми канальцами или даже выводными трубками внутренней системы каналов. Пупочные диски, как и большинство образований дополнительного скелета, обычно стекловидны из-за единообразной (нормальной к поверхности диска) ориентировки слагающих их кристаллов кальцита.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'