
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Историческое развитие
Непосредственными предками фораминифер являются, по-видимому, корненожки с хитиноидной раковиной, а более отдаленными - голые формы, обладающие, подобно современному Тrichosphaerium sieboldi Schaudinn, сходным сложным циклом развития, включающим чередование поколений бесполого и полового, и, соответственно, процессов шизогонии и гамогонии. Более отдаленными предками фораминифер были, очевидно, жгутиконосцы (Mastigophora). На это указывает характер гамет большинства тех представителей подкл. Foraminifera, жизненный цикл которых известен. Наличие амёбоидных гамет - скорее ценогенетическая, чем унаследованная от предков, особенность.
Можно предполагать что примитивные скелетные образования фораминифер возникли в результате уплотнения пелликулярного покрова протоплазматического тела. Так возникли, по-видимому, хитиноидные (псевдохитиновые, тектиновые) раковины несохранившихся в ископаемом состоянии представителей отряда Allagromiida, среди которых следует искать предков всех прочих фораминифер (рис. 24).
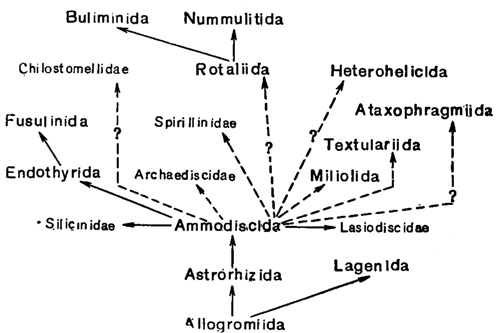
Рис. 24. Схема филогенетического развития отрядов и некоторых семейств фораминифер
В наиболее примитивном отряде фораминифер с минеральной раковиной - Astrorhizida - представлены те же типы строения раковины, что и в отр. Allogromiida, но в качестве отличительной и притом прогрессивной черты выступает способность к образованию жесткой раковины. Предполагается, что формы, агглютинирующие посторонние частицы, возникли независимо от секретирующих известковую раковину; как те, так и другие взяли начало параллельно от форм с хитиноидной раковиной (Galloway, 1933). В отр. Astrorhizida наблюдаются оба типа жестких скелетных образований - агглютинированные и секреционные известковые. Это позволяет выделить два надсемейства: Astrorhizidea и Parathuramminidea, отвечающие двум основным направлениям развития в пределах данного отряда. С выработкой у агглютинированных форм способности к выделению минерального цемента раковины, строго говоря, становятся одновременно и агглютинированными и секреционными.
Жесткая раковина явилась важным защитным приспособлением, но с ее появлением возникло противоречие между непрерывно растущим пропластом и препятствующей его росту раковиной; это противоречие разрешилось путем выработки способности раковины к росту, который на первых порах, у Astrorhizida в частности, осуществлялся либо вставочным (интеркаляционным) путем за счет включения в состав стенки новых агглютинированных песчинок, либо путем наращивания раковины вокруг постепенно смещающегося кпереди устья - надстроечный (суперпозиционный) рост. Вставочный рост наблюдается у Saccamminidae и отчасти у Astrorhizidae. Этот тип роста сам по себе весьма ограничен и не получил существенного развития в эволюции отр. Astrorhizida и вообще фораминифер. Наоборот, надстроечный рост открывает большие возможности, и в эволюции фораминифер он привел через стадию ритмичного надстроечного роста к прерывистому росту и многокамерности, с чем связано необычайное разнообразие строения раковины фораминифер. Среди Astrorhizida типично надстроечный рост наблюдается у Hyperamminidae и Caligellidae; у них же известен ритмичный надстроечный рост (соответственно, у Hyperamminoides, Paratikhinella), тогда как у Reophacidae и наиболее развитых Caligellidae (Tikhinella) имеет место многокамерность, связанная с прерывистым ростом.
Эволюция подкласса фораминифер шла, по меньшей мере, в двух направлениях (рис. 24). Одно из них привело к развитию отр. Lagenida. Это направление характеризуется образованием стенки, сложенной радиально ориентированными кристаллами кальцита. У более развитых представителей отр. Lagenida подобная текстура связана с наличием всегда очень тонких поровых канальцев. Развитие отряда начинается, по-видимому, в ордовике с однокамерных форм подсем. Umbellinae, родственных наиболее примитивным Parathuramminidae, и через стадию одноосных многокамерных, известных с девона, приводит к образованию различного строения спиральных форм, появляющихся, вероятно, также в девоне. В дальнейшем, в особенности, в мезозое, развиваются разнообразные гетероморфные, а также вторично одноосные и вторично однокамерные формы. Возможно, что предками Lagenida были не однокамерные Umbellinae, представляющие в таком случае слепую ветвь, а одноосные многокамерные Caligellidae. Этот вопрос, остается до сих пор неясным из-за неполноты геологической летописи.
Второе, более важное направление дало начало отряду Ammodiscida и, по-видимому, всем остальным. Это направление характеризуется развитием спирального типа строения, на первых порах очевидно, неправильно клубковидного. Возможно, что отр. Ammodiscida представляет собою сборную группу, куда включены формы, происходящие, с одной стороны, от агглютинированных Astrorhizidea (сем. Hyperamminidae), а с другой - от Parathuramminidea (сем. Calligellidae), с их секреционной раковиной. Развитие отр. Amodiscida начинается в силуре от неправильно клубковидных форм, раковина которых образована лишенной септации; трубчатой камерой (обособлена только начальная камера, как у Hyperamminidae), далее, в силуре - девоне развиваются такие же, но спирально-плоскостные формы. Последующая, эволюция Ammodiscida характеризуется постепенным переходом к многокамерным спиральноплоскостным формам путем сперва ритмичного, а затем прерывистого роста. Наиболее высоко организованные Ammodiscida (сем. Lituolidae) имеют сравнительно сложно построенную агглютинированную раковину, далеко уступающую, однако, наиболее сложным известковым скелетным образованиям, развивающимся у представителей некоторых других отрядов. Эти специализированные агглютинированные Lituolidae представляют собою, по-видимому, слепую ветвь.
От примитивных агглютинированных, возможно, еще неправильно клубковидных Ammodiscida, берут начало, вероятно, уже в силуре спирально-конические агглютинированные Ataxophragmiida (сем. Trochamminidae), из которых развиваются формы со спирально-винтовыми, нередко гетероморфными раковинами. Отряд. Ataxophragmiida является также одной из слепых ветвей в развитии фораминифер; наиболее специализированные его представители (из сем. Orbitolinidae и некоторые другие) отличаются относительно сложным строением раковины.
Отр. Textulariida, известный с девона, несмотря на преобладающий спирально-винтовой тип строения раковины, вряд ли является близко родственным Ataxophragmiida. На это указывает спирально-плоскостное строение начального отдела раковины многих Textullariida (в отличие от спирально-конического, трохоидного у Ataxophragmiida). Предков Textuariida следует искать среди примитивных Lituolidae в среднем палеозое, хотя возможно, что некоторые Textulariidae, не имеющие спирально-плоскостной стадии (подсем. Palaeotextulariinae), возникли раньше (в ордовике) и независимым путем; в таком случае отр. Textulariida (и, соответственно, сем. Textulariidae) представляет собою сборную группу, обнимающую две независимые ветви развития (Paleotextulariinae и Textulariinae). На это указывает и преимущественно секреционный характер стенки первой из названных групп.
Наиболее высоко организованные представители отр. Textulariida отличаются относительно простым строением раковины. Этот отряд, подобно предыдущему, является одной из слепых филогенетических ветвей фораминифер.
От Ammodiscida с секреционной известковой раковиной, имеющей примитивную еще септацию, во второй половине девона берет начало отр. Endothyrida. Необходимо отметить, что развитие септации среди Ammodiscida происходило неоднократно, в частности среди Tournayellidae в самом конце девона - в нижнем карбоне, т. е. и после появления Endothyrida. Характерные особенности, наблюдающиеся среди Endothyrida, - навивание раковины по спирали, сопровождающееся изменением ориентировки оси, так называемое плектогироидное навивание, наличие базальных образований дополнительного скелета (в особенности типа хомат), послойная дифференцировка структуры стенки. Эти особенности заставляют видеть в некоторых Endothyrida предков отр. Fusulinida, появившегося в начале каменноугольного периода. Мезозойскими потомками сем. Endothyridae являются представители сем. Mesoendothyridae, а также, возможно, сем. Spirocyclinidae.
В отр. Fusulinida достигается наивысшая для палеозойских фораминифер дифференцировка в строении раковины. Опорно-защитные функции раковины, связанные с увеличением ее механической прочности, разрешаются у Fusulinida несколькими способами путем выработки различных приспособительных особенностей: развитие складчатости септ, отложения различного рода дополнительного скелета, образования вторичных перегородок, изменения микроструктуры стенки.
Для отдельных направлений эволюции в пределах отр. Fusulinida характерно развитие тех или иных приспособительных особенностей или известного сочетания этих последних. Высокая специализация Fusulinida явилась причиной как их господства в верхнем палеозое, так и их полного вымирания ко времени завершения эпохи герцинского орогенеза.
Отр. Miliolida берет, по-видимому, начало от каких-то верхнепалеозойских Ammodiscida, имеющих секреционную известковую раковину с беспорядочным расположением слагающих ее кристаллов кальцита. Такая структура стенки присуща всем Miliolida, за исключением Peneroplis и др., у которых начальная камера прободенная, что является, возможно, ценогенетической особенностью - новообразованием. Развитие в пределах отр. Miliolida идет от неправильно завитых форм к спиральным. У наиболее специализированных (сем. Miliolidae) имеют место случаи правильного клубковидного навивания, или же их раковины спирально-плоскостные со сложной системой внутренних перегородок (сем. Alveolinidae), внешне напоминающие раковины Fusulinida. Отр. Miliolida - одна из слепых филогенетических ветвей фораминифер.
Весьма вероятно, что от Ammodiscida или от общего с ними предка взяли начало спирально свернутые Silicinidae, обладающие, в отличие от прочих фораминифер, кремневой раковиной.
С известным сомнением производятся от отр. Ammodiscida Spirillinidae с известковой спирально-плоскостной или трохоидной раковиной, со слабо выраженной септацией или совсем ее лишенные и занимающие совсем особое положение среди фораминифер в силу наличия амёбоидных гамет у современных представителей этого семейства.
Крупным стволом в эволюции фораминифер является отр. Rotaliida, представленный многочисленными семействами, общей особенностью которых является трохоидная пористая известковая раковина. Предками этого отряда, появляющегося в начале мезозоя, лишь весьма предположительно могут быть примитивные Ammodiscida, из числа обладающих секреционной раковиной. Развитие в пределах отр. Rotaliida шло в нескольких направлениях: от сравнительно просто построенных уже многокамерных Discorbidae к имеющим сложное строение перегородок и устья Ceratobuliminidae и Epistominidae, к имеющим систему каналов Rotaliidae, к Nonionidea, давшим, с одной стороны, сем. Nonionidae, с другой ряд групп, приспособившихся к сидячему и прикрепленному образу жизни (сем. Planorbulinidae, Homotremidae и др.). Также от Discorbidae берут начало в юрском периоде планктонные Globigerinidae, от которых происходят Globorotaliidae, появившиеся, по-видимому, в нижнемеловой эпохе. Сем. Elphidiidae связано, по ряду соображений, генетически с Rotaliidae и также характеризуется развитием системы каналов.
Отр. Buliminida, представители которого известны с юры и отличаются преимущественно спирально-винтовым типом строения, происходят от сем. Discorbidae. На это указывает строение начального отдела раковины некоторых Buliminida.
Наконец, отр. Nummulitida, появляющийся в верхнем мелу, обнимает наиболее высоко организованных, притом крупных по своим размерам фораминифер, происходящих, по-видимому от Rotaliida. У некоторых Nummulitida (сем. Discocyclinidae) начальный отдел раковины, особенно микросферической, сохраняет трохоидное строение. Раковина Nummulitida отличается сложной дифференцировкой частей, значительным развитием дополнительного скелета, и, как правило, системы каналов. Может быть, этот отряд представляет собою сборную группу, причем сем. Nummulitidae, с одной стороны, и остальные семейства - с другой, отвечают, по меньшей мере, двум самостоятельным направлениям филогенетического развития.
Если рассматривать развитие фауны фораминифер в целом как совокупности известных родов, семейств и отрядов, то можно отметить несколько основных этапов этого развития, достаточно отчетливо выраженных и в пределах СССР. Наиболее древняя - первая фауна фораминифер - происходит из отложений ордовика и силура, вторая характерна для девонских и нижнекаменноугольных отложений, третья развивается в верхнем палеозое, четвертая - в триасе - нижнем мелу, пятая - в верхнем мелу, шестая - в палеогене и седьмая - от неогена до современных морей (рис. 25). Каждая из перечисленных фаун отличается особенностями систематического состава с преобладающим развитием определенных систематических групп (семейств, отрядов). Каждая последующая фауна генетически связана с ей предшествующей, в которой имеет свои корни в виде еще редких представителей тех групп, которые в последующей фауне становятся господствующими.
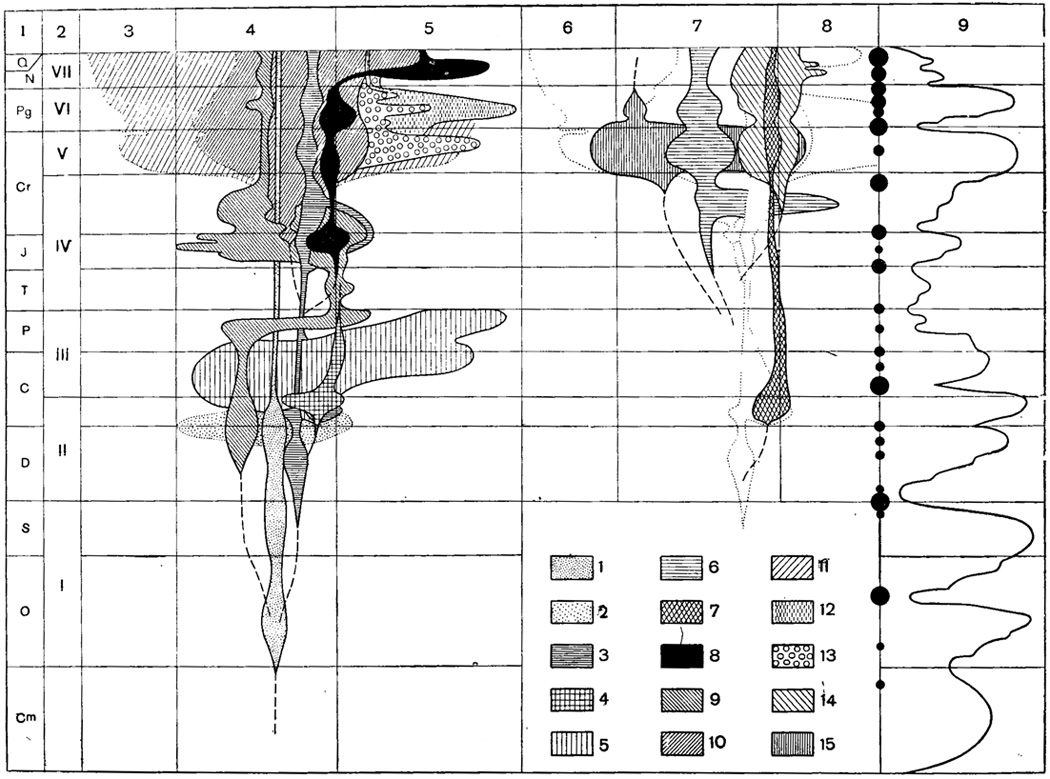
Рис. 25. Схема развития фаун фораминифер в геологическом прошлом. Левая часть схемы: 1-я колонка - геологические периоды; 2-я колонка - последовательные фауны фораминифер (I - ордовикско-силурийская; II - девонская - нижнекаменноугольная; III - верхнипалеозойская; IV - триасовая - нижнемеловая; V - верхнемеловая; VI - палеогеновая; VII - неогеновая - современная; 3-5-я колонки - относительное развитие фаун фораминифер; 3 - в областях океанических впадин (пелагиаль), 4 - эпиконтинентальных и 5 - геосниклинальных бассейнов; степень и область развития отдельных отрядов, а также филогенетические отношения между ними (пунктир - черточками) показаны приблизительно. Правая часть схемы; 6-8-я колонки - дополнения схемы относительного развития фаун фораминифер; 6 - пелагиаль, 7 - эпиконтинентальные и 8 - геосинклинальные бассейны; точечным пунктиром с целью совмещения с левой частью чертежа даны контуры развития отрядов Rotaliida и Ammodisclda; 9-я колонка - трансгрессии и регрессии моря (по Страхову, 1948); на границе между 8-й и 9-й колонками черными кружками различного диаметра показаны складкообразовательные движения различной интенсивности. Внизу справа условные обозначения для различных отрядов фораминифер: 1 - Astrorhizida, бентические формы; 2 - то же, планктонные формы (Parathuramminidae); 3 - Ammodisclda; 4 - Endothyrida; 5 - Fusulinida; 6 - Ataxophragmiida; 7 - Testulariida; 8 - Miliolida; 9 - Lagenida; 10 - Rotaliida, бентические формы; 11 - то же, планктонные формы; 12 - Nummulitida (сем. Nummulitidae); 13 - Nummulitidae - сем. Orbitoididae, Discocyclinidae, Miogypslnidae; 14 - Bullminida; 15 - Heterohelicida (Фурсенко)
Время существования отдельных семейств фораминифер показано в таблице (рис. 26). Естественно предполагать, что фораминиферы появились еще задолго до ордовика.
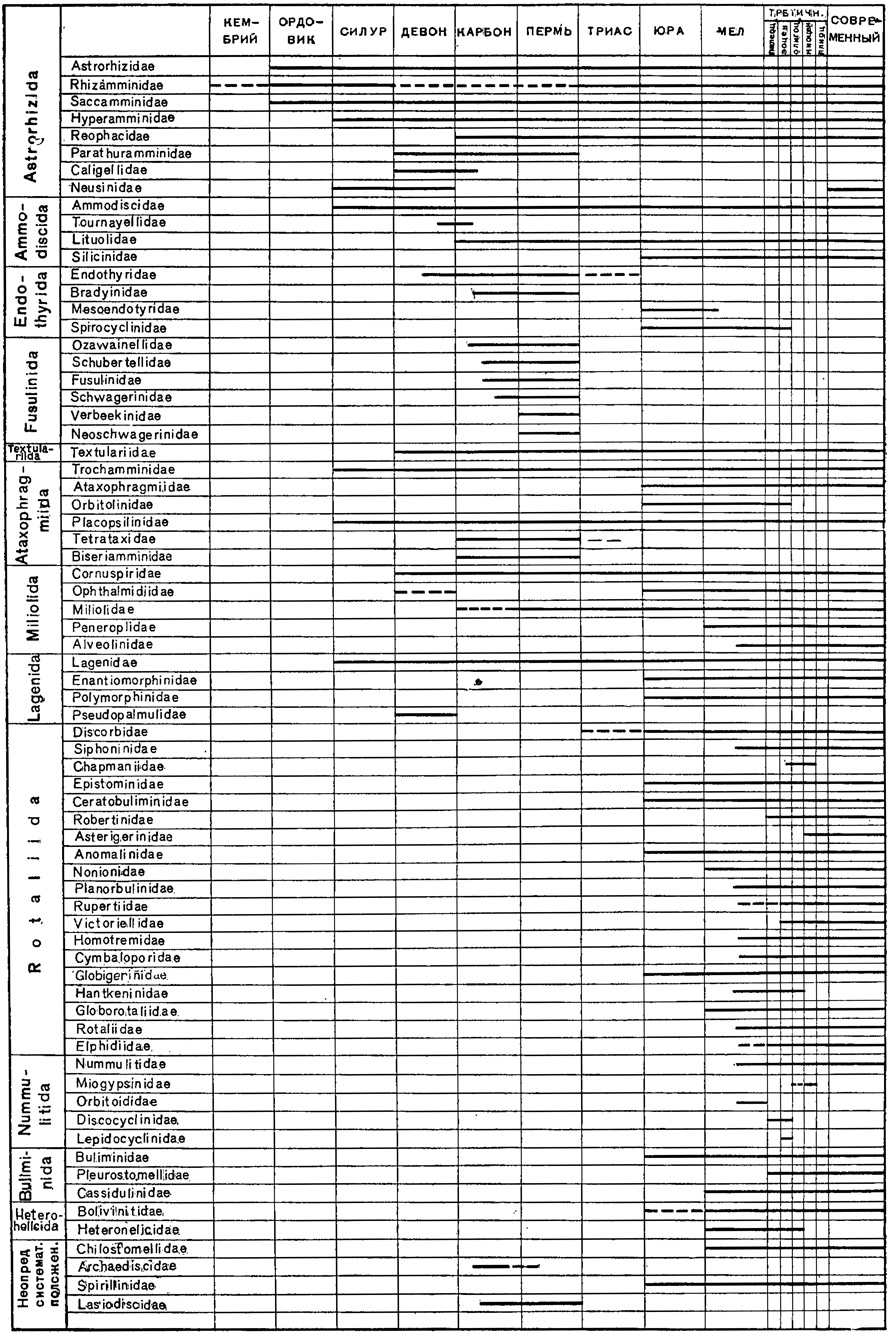
Рис. 26. Стратиграфическое распределение семейства фораминифер
Фауна фораминифер ордовика - силура Прибалтики представлена исключительно однокамерными формами, с одной стороны - агглютинированными из отр. Astrorhizida, с другой - примитивными формами, условно отнесенными к сем. Lagenidae (возможно, сборное подсем. Umbellinae; Быкова, 1956). Из ордовика и силура С. Америки, помимо представителей отр. Astrorhizida, известны первые примитивные Ammodiscidae. Фораминиферы ордовика - силура отличаются небольшими размерами и приурочены в основном к карбонатным мелководным фациям.
Для средне-верхнедевонской и нижнекаменноугольной фауны фораминифер СССР еще характерно присутствие ряда примитивных форм из отр. Astrorhizida. Значительного разнообразия достигают представители известных с девона сем. Parathuramminidae и Caligellidae. Роды этих двух примитивных семейств характерны в большинстве своем и для нижнего карбона; весьма важно, что в сем. Caligellidae можно проследить постепенный переход от неподразделенных трубчатых одноосных форм к родам с раковиной, имеющей септацию, как и у характерного для верхнего девона - нижнего карбона СССР сем. Tournayellidae. Впрочем, из верхнего девона СССР известны довольно многочисленные и подчас довольно высоко организованные многокамерные фораминиферы. Так, наряду с примитивными Umbellinae, широко развитыми в верхнем девоне, в этом же отделе присутствуют и такие высоко развитые одноосные Lagenidae, как Multiseptida, многокамерные Nanicella и др.
В нижнем карбоне достигают расцвета многокамерные Endothyridae, одна из ветвей которого дала в нижнекаменноугольную эпоху начало отр. Fusulinida. Отличительной особенностью нижнего карбона является присутствие играющих существенную роль в составе фауны Textulariidae и Bradyinidae. Lituolidae существуют, очевидно, с девона, но в нижнем карбоне они дают характерную ветвь - Haplophiagmellinae. В нижнем же карбоне появляется исключительно палеозойское сем. Archaediscidae, занимающее среди фораминифер особое положение.
Для верхнепалеозойского этапа эволюции фораминифер характерно значительное развитие отр. Fusulinida. Наряду с ним в верхнем палеозое встречаются представители таких появившихся в среднем палеозое семейств, как Endothyridae, Bradyinidae, Textulariidae, Tetrataxidae, Archaediscidae, но здесь они играют подчиненную роль. В верхнем палеозое, особенно в верхней перми, значительного расцвета достигают Lagenida, среди которых отмечается ряд специализированных форм (Pachyploia, Colaniella и др.). Сем. Lagenidae, достигающее значительного расцвета в верхнем палеозое, продолжает усиленно развиваться в последующие геологические периоды. Для верхнего палеозоя, особенно для перми, характерно своеобразное сем. Lasiodiscidae. В каменноугольном периоде появляются, по-видимому, первые Miliolidae.
Триасовый - нижнемеловой этап эволюции фораминифер характеризуется расцветом сем. Lagenidae, дальнейшим развитием агглютинированных многокамерных форм, особенно из Lituolidae, а также появлением Polymorphinidae, Trochamminidae, Ataxophragmiidae, Orbitolinidae, Discorbidae, Anomalinidae, Epistominidae, Ceratobuliminidae и др.
Фораминиферы триаса изучены недостаточно; их находки известны лишь на С. Кавказе, на севере Азиатской части Советского Союза и в некоторых областях З. Европы (Англия, Ю. Германия, Австрия, Венгрия). Фауна фораминифер триаса С. Кавказа носит смешанный характер: даже в норийском ярусе встречаются Tetrataxis, Robuloides и др., мало отличающиеся от верхнепермских представителей этих родов, - а представители рода Nodosaria в известной своей части напоминают пермские формы. Существенно новым является значительное развитие родов Ophthalmidium и Spirophthalmidium (А. Д. Миклухо-Маклай, 1952).
Значительно лучше известны фораминиферы юры и нижнего мела. Здесь развиваются разнообразные Lagenidae, в том числе спиральные формы; одноосные, начиная со средней юры, отступают на второй план. Продолжают развиваться Ophthalmidiidae. Для некоторых фаций юры и нижнего мела, в частности - Русской платформы и некоторых других областей, характерно развитие Epistominidae и нередко, сопутствующих им Ceratobuliminidae. Появляются Polymorphinidae и некоторые другие. Господствующими до конца нижнемеловой эпохи остаются Lagenidae и отчасти агглютинированные формы; последние в некоторых фациях преобладают.
Для нижнемеловых мелководных бассейнов области Тетиса характерно развитие сем. Orbitolinidae. В отличие от верхнепалеозойских, фораминиферы триаса, юры и нижнего мела даже в карбонатных фациях никогда не бывают породообразующими.
Характерной особенностью верхнемеловой фауны фораминифер является отмечающийся впервые в истории развития этого подкласса расцвет планктонных групп - сем. Globigerinidae и Globorotaliidae, появление крупных форм новых сем. Rotaliidae, Calcarinidae, Orbitoididae и некоторых других, господство из агглютинированных Ataxophragmiidae, а из имеющих известковую раковину - Anomalinidae, Bolivinitidae, Heterohelicidae, Buliminidae и др. Широко распространенные фации белого мела в пределах каждого данного яруса характеризуются значительной однотипностью.
В то же время отличия комплексов форм отдельных ярусов, а подчас и более дробных систематических подразделений, обычно весьма резки. Это связано в значительной мере с быстрыми темпами эволюции в отдельных группах фораминифер (Anomalinidae, Buliminidae и, особенно, Globorotaliidae).
Для верхнемеловой эпохи характерно весьма широкое расселение фораминифер как в эпиконтинентальных, так, в известной мере, и в геосинклинальных бассейнах. При этом фораминиферы встречаются в самых различных фациях и впервые в истории земли начинают играть весьма существенную роль в фауне полагиали.
Для фауны фораминифер палеогена характерно дальнейшее развитие ряда верхнемеловых групп. По-прежнему исключительно важную роль играют планктонные Globigerinidae и Globorotaliidae, но последние представлены другими родами - Globorotalia и ему родственными; глоботрунканы, вымирающие к концу верхнего мела, отсутствуют. Роль Heterohelicidae становится незначительной. Существенную роль в эпиконтинентальных бассейнах, помимо планктона, играют бентические Ataxophragmiidae, Anomalinidae, Buliminidae, Bolivinitidae и др.
Фораминиферы палеогена расселяются еще шире, чем верхнемеловые. При этом отмечается большое разнообразие фациальных типов фауны, каждый из которых представлен в ряде областей. Геосинклинальные области, в данном случае - Тетиса, становятся местами исключительного расцвета крупных фораминифер; в этом отношении начальный этап альпийского орогенеза по исключительному развитию отр. Nummulitida напоминает герцинский этап с характерным для него господством отр. Fusulinida.
Фауна фораминифер неогена отличается рядом особенностей, основной из которых является развитие групп, приспособившихся к особым условиям существования, возникшим благодаря образованию в некоторых областях Тетиса и с ними связанных более или менее замкнутых бассейнов. В других бассейнах, сохранивших связь с мировым океаном, развивалась фауна морского типа, сходная с палеогеновой по составу основных групп фораминифер, но все же значительно отличающаяся в систематическом отношении. Наиболее своеобразна фауна фораминифер Крымско-Кавказской области и с ней сопредельных территорий. Здесь развивались исключительно разнообразные и в значительной мере эндемичные формы из сем. Miliolidae, отчасти Elphidiidae, Nonionidae и другие. При установлении временных связей с западноевропейскими бассейнами фауна Крымско-Кавказской области обогащалась средиземноморскими элементами (Polymorphinidae, Rotaliidae, Cassidulinidae, Globigerinidae, Buliminidae и др.).
В некоторых сохранивших связь с океаном мелководных областях Тетиса и прилегающих к нему эпиконтинентальных бассейнах продолжают развиваться крупные формы, причем на смену палеогеновым Discocyclinidae приходят Miogypsinidae и некоторые другие. В известной мере самобытная фауна фораминифер развивается в области Восточной и Западной Тихоокеанских геосинклиналей: Lituolidae, отчасти Lagenidae, Polymorphinidae, Elphidiidae, в основном Cassidulinidae, Buliminidae, Nonionidae и др.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'