
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Скелет и его развитие
Химический состав. По химическому составу скелет радиолярий у различных отрядов неодинаков, вследствие чего их остатки неодинаково способны сохраняться в ископаемом состоянии.
Скелет спумеллярий и насселлярий по химическим свойствам, по растворимости в кислотах бесспорно признается кремнистым. Ископаемые радиолярии относятся исключительно к этим двум отрядам.
Скелетные части Sticholonchea состоят лишь частью из кремнезема, поскольку плавиковая кислота их растворяет неполностью. В ископаемом состоянии скелет Sticholonchea не известен.
Скелет феодарий также признается кремнистым, однако в данном случае еще требуются дополнительные исследования. Дело в том, что скелетные элементы представителей этого отряда не сохраняются в ископаемом состоянии. Их нет и в донных отложениях океанов в результате разрушения, разложения в воде при погружении на дно отмерших панцирей. Кроме того, Решетняк подметила своеобразную особенность панцирей представителей сем. Tuscaroridae, растворявшихся иногда в гвоздичном масле при изготовлении тотальных препаратов.
Наконец, в отношении скелета акантарий установился взгляд, что он состоит из целестина - сернокислого стронция (SrSO4). Однако и тут требуются дополнительные исследования в силу наличия существенных разногласий между разными авторами.
Стронциевый скелет у акантарий был установлен Бючли в результате химического анализа игл. аберрантной сидячей формы Podactinellius sessitis. Но Шевяков (1926) вполне обоснованно высказывает сомнение в принадлежности Podactinellius к акантариям (и вообще к радиоляриям). Полученные выводы Бючли распространил на несколько видов настоящих акантарий, только наблюдая у них фигуры травления, подобные отмеченным им для Podactinellius; спектральный анализ вещества скелета и здесь не был применен. В противоречии с данными Бючли стоят исследования Шевякова (1926). На основе тщательного химического анализа Шевяков в составе скелета акантарий установил наличие сложного алюмокальциевого силиката с содержанием 21,6% SiO2. Более новые данные X. Одума (Odum, 1951),на основании спектроскопического анализа и исследования в рентгеновых лучах, подтверждают наличие целестина в скелете акантарий; однако слишком малые количества исследованного материала (что признает и сам автор) еще не позволяют считать точно установленным целестиновый состав скелета акантарий.
В ископаемом состоянии скелет акантарий не сохраняется, так же как нет его и в донных отложениях океанов; Шевяков говорит, что скелет акантарий растворяется в воде после стояния в ней в течение нескольких часов*.
* (Надо сказать, что эти данные плохо увязываются с довольно слабой растворимостью кристаллов целестина минерального происхождения, повышающейся в водах, содержащих свободный хлор и углекислоту. В литературе имеются немногочисленные непроверенные сообщения о находках в ископаемом состоянии плохо сохранившихся остатков целестиновых скелетов акантарий например в морских третичных мергелях на С. Кавказе (Ферсман, 1945) (притч, редактора. - А. X.))
Элементы симметрии скелета. Наиболее распространенная у радиолярий шарообразная форма тела сочетается с многоосной симметрией скелета. У многих спумеллярий эта многоосность носит неопределенный (гомаксонный) характер (например в подотр. Sphaeroidea), сменяясь ограниченным числом осей (полиаксонностью), когда от шарообразного панциря отходит определенное число игл. Точно так же неопределенная многоосная симметрия свойственна тем феодариям, у которых шарообразное тело покрыто скелетной сеточкой (сем. Cannosphaeridae и Sagosphaeridae) или когда из него торчат многочисленные радиальные иглы (сем. Aulacanthidae).
Частой модификацией многоосного скелета у радиолярий является преобладание одной из осей, вдоль которой тело радиолярии или вытягивается, или сплющивается, или около которой по радиусам правильным образом располагаются части скелета - получается радиальносимметричный скелет всех акантарий, многих спумеллярий (особенно у тех, вроде Stytosphaera, Pipetta, Panarium, у которых сильно развиты две полярных иглы, одинаковых или разных по длине, становящихся осью тела), феодарий (сем. Castanellidae, Tuscaroridae) и насселлярий (род Cryptocalpis) с расположенным на одном полюсе скелетного панциря устьем. Наконец, на этой радиально-симметричной основе развивается и двустороннесимметричный скелет, когда получают преобладание два каких-нибудь противоположных радиуса (например Amphilonche среди акантарий и др.). Иной характер носит двусторонняя симметрия ряда феодарий (например в сем. Coelodendridae), развивающаяся на основе двустворчатого панциря сем. Concharidae.
Особый характер усложнения симметрии, редко проявляющийся у насселлярий (род Spirocampa), представляет винтовое скручивание панциря вдоль оси скелета. Подобное скручивание в одном направлении, перпендикулярно оси панциря, приводит к образованию плоских дисков спумеллярий (подотряд Discoidea) со спиральным расположением ячеек скелета.
Наконец, в ряде случаев скелет асимметричен. Первичная асимметрия свойственна скелетным иглам с несколькими отростками у примитивных спумеллярий и насселлярий. У последних на основе такого устройства скелета развивается асимметричный решетчатый панцирь с раструбом внизу. Вторичная асимметрия у высших представителей отр. Phaeodaria (род. Euphysetta, сем. Medusettidae) развивается на основе радиальной симметрии.
Наряду с проявлением различных ступеней симметрии в скелете одноосных радиолярий в сравнительно редких случаях развивается метамерное расположение камер скелетного панциря вдоль оси тела. Такую картину мы можем наблюдать как у спумеллярий (сем. Panartidae, Zygartidae, подотр. Prunoidea), так и у насселлярий (Eucyrtidium, Artocapsa, Stichocapsa и других представителей сем. Lithocampidae). Число камер, расположенных в ряд, начиная с двух, может достигать 10 и более. Экологическое значение различных форм симметрии у радиолярий рассмотрено В. Н. Беклемишевым (1952).
Общая конструкция скелета. Бесскелетное состояние, наблюдаемое в трех отрядах радиолярий (Spumellaria, Nassellaria, Phaeodaria) может быть первичным и вторичным. Чаще мы толкуем отсутствие скелета как первичное состояние, предшествующее развитию примитивного скелета в виде отдельных игл. (в подотр. Sphaerocoilidae среди спумеллярий, в сем. Caementellidae среди феодарий). С другой стороны, в сем. Medusettidae (отр. Phaeodaria) скелет именно исчезает; первичный скелет, редуцируясь, заменяется слабо развитым вторичным, исчезающим в свою очередь.
Если скелет отсутствует, поверхность тела радиолярии защищена посторонними частицами, налипающими на поверхность калиммы. Такое агломерированное состояние панцирями диатомей, мелких радиолярий, обломками их игл имеем у Phaeodaria в случае первичного отсутствия скелета (Caementelta) и у форм с вторичным исчезновением скелета (Miracella из сем. Medusettidae) (рис. 11). В этих случаях возникает известное экологическое подобие между такими формами мелких радиолярий и формами "песчаных" фораминифер.
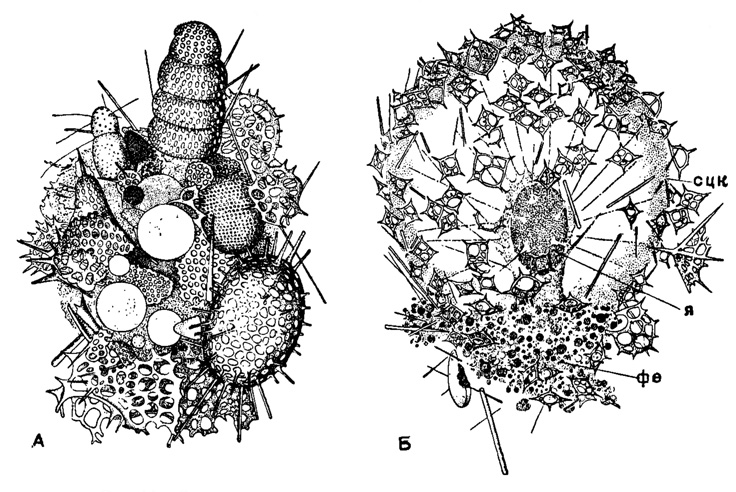
Рис. 11. Радиолярии с наклеенными посторонними частицами: А - Caementella ioricata Borgert (Phaeodarla, Caementellidae); современный вид из Атлантического океана, на поверхности тела - панцири радиолярий, × 175 (Borgert. 1909); Б - Miracella ovulum Borgert (Phaeodaria, Medusettidae); современный вид из Средиземного моря, на поверхности тела - обломки панцирей радиолярий и силикофлагеллят, × 175 (Borgert, 1913): сцк - стенка центральной капсулы; я - ядро; фе - феодни
Скелет из отдельных игл, лежащих в калимме, представляет сравнительно редкое состояние у радиолярий, характеризующее примитивные стадии их эволюции. Прямые, изогнутые, гладкие, шиповатые, простые или разветвленные иглы могут лежать свободно в теле радиолярии, или могут соединяться вместе в каком-то центре, скрепляясь проксимальными концами в один пучок (чаще) или в несколько пучков (значительно реже); или игольчатые элементы, уже в виде перекладин, соединяются обоими концами в различного рода поверхностные сети с крупной ячеей.
Формирование шаровидных продырявленных скелетов, весьма распространенных у радиолярий, может осуществляться двояко. С одной стороны, такие скелеты возникают у акантарий и спумеллярий (исторически и в онтогенезе) путем соединения концов боковых ответвлений (апофизов) радиальных игл. При наличии нескольких ярусов этих ответвлений может возникнуть скелет из нескольких концентрических, вложенных один в другой ажурных шаров (рис. 12). У акантарий это явление исключительно редко (формируется два таких шара один в другом), в то время как у спумеллярий оно очень распространено (пять шаров и более может быть вложено один в другой) (рис. 13).

Рис. 12. Staurocarium arborescens Haeckel (Spumellaria, Staurosphaeridae); современный вид из Индийского океана, скелетные шары, вложенные один в другой, × 130 (Haeckel, 1887)
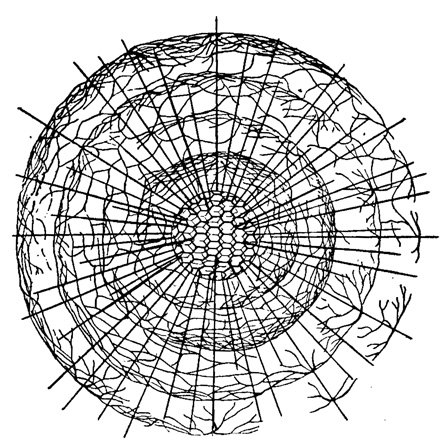
Рис. 13. Aracknosphaera myriacantha Haeckel (Spumellaria, Astrosphaeridae); современный вид из Средиземного моря; многоярусные апофизы на иглах, образующие скелетные шары, вложенные один в другой, × 150 (Haeckel, 1862)
Другой тип образования шарообразных скелетов-панцирей мы видим у феодарий, где они возникают из расположенных поверхностно слившихся вместе тангентальных игл. В пользу подобного образования шаров говорят во многих случаях сохранившиеся тангентальные иглы в стенках панцирей. Кроме того, в ряде случаев наблюдается развитие особой губчатой поверхностной обкладки, явно происшедшей благодаря спайке концов тангентальных игл. Точно так же и сплошная раковинка у Tuscaroridae (Phaeodaria) возникает из слоя тангентальных игл.
Иные формы панциря, весьма разнообразные, будут еще разобраны при рассмотрении скелета отдельных отрядов. Здесь уместно заметить, что для всего подкласса радиолярий в целом трудно наметить единый план строения скелета; но все же схожие, общие черты строения скелета можно найти в пределах отдельных отрядов.
Акантарии обладают исключительно астроидным скелетом: сплошные иглы скелета расходятся звездообразно от центра тела. Вторично в этом отряде может развиваться сфероидный скелет, его шарообразные формы.
Для спумеллярий типичен преимущественно сфероидный скелет, чаще в сочетании со сплошными радиальными иглами, но не сходящимися в центре тела, а берущими начало от внутреннего скелетного шара. Очень распространено у спумеллярий концентрическое вложение многих шаров один в другой. При сплющивании шаровидного скелета может получиться дисковидная форма скелета, со спиральным расположением камер.
Для насселлярий типичен скелет в виде башенки, сигары или колокола, открытого с одной стороны, развивающийся в результате разрастания ветвей скелетной иглы типа треножника.
У феодарий решетчатые шары всегда одинарные, с устьем (пиломом). Радиальная симметрия одноосной раковинки может замениться двусторонней симметрией вплоть до двустворчатой раковинки. Полые иглы располагаются радиально или слагаются в рыхлую поверхностную сеть.
Наконец, у Sticholonchea иглы сочленяются проксимальными концами, образуя несколько пучков, торчащих во все стороны из тела радиолярии.
Конструкцию скелета радиолярий подверг математическому анализу Д. Д. Мордухай-Болтовской.
Расчеты, приведенные в работе Д. Д. Мордухай-Болтовского, показывают, что у очень многих радиолярий шаровидная форма скелета или многогранная с двадцатью гранями - икосаэдра (у Circogonia из феодарий, у форм из акантарий с их 20 иглами) наиболее экономна по расходу строительного материала.
У многогранников с меньшим числом граней экономность затрат материала на построение панциря повышается искривлением граней (по дугам круга или, вероятнее, гиперболы). Еще большая экономия при этом достигается продырявливанием стенок скелета, наблюдающимся у громадного большинства форм. Ячейки на поверхности скелета, петли поверхностной сети, нередко принимают форму шестиугольников, поскольку на их формирование, по расчетам, расходуется меньше всего материала.
Однако у радиолярий существуют также и четырехугольные решетки; Мордухай-Болтовской доказывает, что разные формы скелетной решетки у радиолярий имеют механически различное назначение. Шестиугольная решетка, например, рассчитана на равномерное давление, четырехугольная - на удар, будучи более прочной. Могут быть приведены примеры (некоторое Astrosphaeridae из спумеллярий), когда у одной формы радиолярий имеются разные решетчатые сферы, вложенные одна в другую: внешняя, выдерживающая удары, с четырехугольными ячеями, и внутренняя, испытывающая равномерное давление, шестиугольная.
В случае многогранников, образованных из стержней, наименьшим и, следовательно, наиболее выгодным в смысле расходования материала периметром в одном и том же объеме обладает куб (что и осуществляется у Lithocubus из насселлярий, рис. 14). Полые стержни у феодарий тоже ведут к экономии материала при сохранении прочности трубчатых элементов скелета.
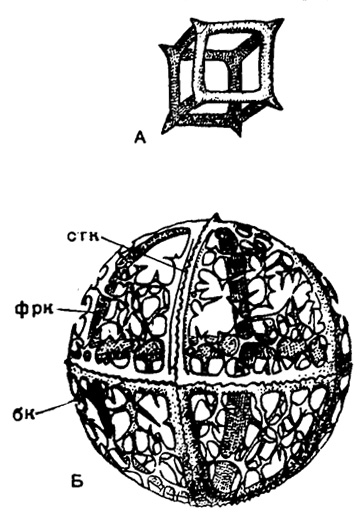
Рис. 14. Кубический скелет насселлярий: А - Lithocubus geometricus Haeckel (Nassellaria, Stephoidae), современный вид из тропической части Тихого океана, × 200. Б - Trissocyclus sphaeridium Haeckel Nassellaria,Stephoidae); современный вид из Индийского океана, × 300 (Haeckel, 1887); сгк - сагиттальное кольцо; фрк - фронтальное кольцо; бк - базальное кольцо
Анализируя конструкции панцирей, пронизанных стержнями, Мордухай-Болтовской доказывает, что тонкие иглы мало изменяют сферическую форму скелета, но при утолщении стержня общая форма панциря меняется; он вытягивается вдоль иглы, принимает эллипсоидальную и даже веретеновидную форму, что и наблюдается у ряда форм радиолярий. Несколько стержней влияют на скелет аналогично (например у Circospathis из Pheodaria).
Подвергая механо-математическому анализу способы образования узлов при соединении скелетных элементов в общую сеть, Мордухай-Болтовской расчетами по формулам механики подтверждает значение соединений для прочности всей системы в целом. Из формул механики следует, что осуществляемые у радиолярий соединения и спайки обеспечивают равномерное распределение нагрузки на каждый скелетный элемент, на перекладину, иглу.
Сходным образом Д'Арси Томпсон (Thompson, 1944) анализирует геометрическую форму скелета радиолярий, проводя параллели с конструкцией игольчатого скелета губок и коралловых полипов.
Физический анализ образования раковинок (у фораминифер) был дан еще Румблером (Rhumbler, 1911) чем объясняется механизм построения различных форм скелета и связывается с упрочением, достигаемым различными способами.
Роль скелета. Скелет радиолярий, столь разнообразный по форме, прежде всего имеет значение опоры, придавая определенную форму протоплазменному телу радиолярии. Скелетные иглы нередко бывают зазубрены, снабжены боковыми ветвями, нитевидными отростками со звездообразными бляшками на концах (спатиллами), затрудняющими сползание протоплазмы с игл. В ряде случаев скелет, одевающий радиолярию панцирем, сеткой, становится защитным образованием, предохраняя ее тело от механических повреждений.
Однако главное значение скелета у радиолярий - в приспособлении к плавучему пелагическому, планктонному существованию. Обилие игл, торчащих в разные стороны из тела радиолярии, вместе с псевдоподиями, значительно увеличивает общую поверхность тела.
Понятно, что различия в динамических условиях среды нередко вызывают отбор и приспособленность по механическим свойствам конструкций скелетов у совокупностей форм среди радиолярий. Так, имеются указания, что фауны радиолярий с преобладанием башенковидных и похожих на колокол форм характерны для условий, где развиты восходящие течения. Подобные фауны (с исключительным преобладанием насселлярий) описаны, в частности, из ископаемых фосфоритов, генезис которых связывается с такими условиями развития восходящих из глубин течений (сравните фауну радиолярий из валанжинских фосфоритов Камско-Вятского бассейна, по Хабакову 1937 г.; некоторые фауны из кремнистых толщ мезозоя на советском Дальнем Востоке, по Жамойда; некоторые мезозойские фауны из, Мексики и др.).
В. А. Догель и В. В. Решетняк (1953) специально обращают внимание на зависимость положения в воде флоттирующих насселлярий от формы их раковинки; например, виды, с растопыренными треугольником в стороны иглами скелета могут парить в воде в состоянии относительного равновесия только в том случае, если они будут обращены треножником книзу.
У акантарий (Diploconus) развиваются двойные парашюты за счет образования конусовидных футляров вокруг двух крупных, преобладающих над остальными игл. У Panartidae среди Spumellaria (подотр. Prunoidea) имеется система парашютов, вытянутых в одну линию (Мордухай-Болтовской, 1934).
Шип на вершине парашюта, вытягивающийся в иглу, поддерживает вертикальное положение панциря (рис. 38). При отсутствии шипа эту роль в поддержании устойчивости могут выполнять зазубрины и бахрома по краю колокола, сильно развитые именно когда нет других приспособлений, вызывающих устойчивость скелета в воде. Ребра на поверхности таких скелетов также способствуют повышению устойчивости. Наконец, появление пережимов на панцире насселлярий, формирование т. н. головного, брюшного и грудного отделов увеличивает боковую поверхность сопротивлении и вместе с тем опять-таки увеличивает устойчивость тела радиолярии в воде (рис. 15).
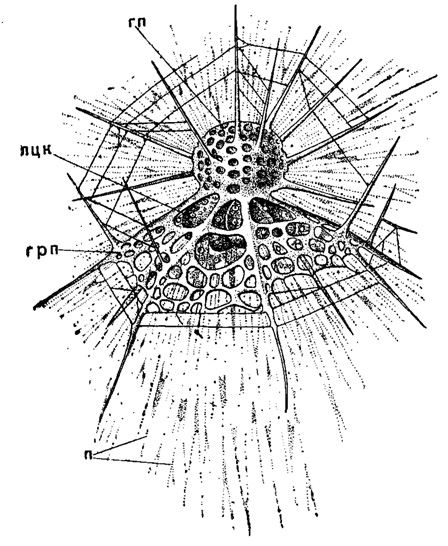
Рис. 15. Arachnocorys circumtexta Haeckel (Nassellaria, Cyrtoidae; живой экземпляр из Средиземного моря, X 400 (Haeckel, 1862): гп - головной отдел панциря; лцк - лопастная центральная капсула; грп - грудной отдел панциря; п - псевдоподии
Мордухай-Болтовской (1934) придает большое значение возможности использований радиоляриями течений воды на основе принципа планера. При этом может быть использовано неравномерное удлинение некоторых экваториальных стержней-игл; тело радиолярии тогда может становиться устойчиво наклонно по отношению к струям воды. Особенно показательны в этом плане Spumellaria Discoidea с крестообразным, пятиконечным или иным скелетом, в котором, как нередко бывает, одна лопасть развита значительно сильнее других. Нагибание шипа на вершине, часто наблюдающееся на колоколе насселлярий, ведет к усилению планирующей способности. Точно так же упомянутый автор на основе расчетов признает, что расположение игл у акантарий по закону Мюллера, когда иглы разных поясов лежат в разных меридианах, более целесообразно, чем то, когда они лежали бы в одном меридиане или были бы иначе приближены друг к другу. Площадь опоры в обоих случаях одна, но между близко лежащими иглами получается мертвая вода, увлекаемая телом радиолярии; расчет же необходимо вести на открытую воду, способную поддержать животное, и у акантарий иглы как раз и расположены в соответствии с подобным расчетом.
Уплощение тела у спумеллярий (подотр. Discoidea) в форме диска также ведет, к увеличению парящей способности. Наконец, отмеченное выше увеличение размеров глубинных форм, по сравнению с близкими поверхностными видами, связано с увеличением внутреннего трения (вязкости) воды при ее охлаждении, по мере продвижения вглубь. При увеличении же линейных размеров тела с сохранением той же формы поверхность тела будет увеличиваться относительно меньше (в квадратической пропорций), по сравнению с изменением объема и веса (кубической пропорции), и более крупные формы для флоттирования, следовательно, должны иметь более вязкую среду, т. е. более холодную, глубинную воду. Поэтому некоторые комплексы ископаемых радиолярий с преобладанием крупных форм можно считать приуроченными к более холодным водам (умеренных областей, области холодных течений или больших глубин), например, в мезозое (Хабаков, 1941; Жамойда, 1956).
Онтогенетическое развитие скелета. Сложные скелеты радиолярий претерпевают значительные изменений в процессе индивидуального развития. В ряде случаев удается заметить стадии онтогенеза, например, у насселлярий с разной степенью развития оторочки брюшного отдела. Однако стадии развития прослежены сколько-нибудь подробно только у представителей отряда акантарий, еще не найденного в ископаемом состоянии.
Онтогенетическое развитие скелета в зиготах у многих видов акантарий очень подробно и на большом материале проследил Шевяков (1926).
Онтогенетическое развитие у акантарий идет в совершенно особых условиях, отличных от существования взрослых радиолярий. Зиготы акантарий опускаются из поверхностных слоев моря на глубины до 400 м, падают на дно и в массе рыхлого детрита проделывают свое развитие в течение зимы и ранней весной (в условиях Средиземного моря).
Иглы скелета в полном количестве (10 диаметральных у отр. Holacantha и 20 радиальных у прочих акантарий) закладываются уже на самых ранних стадиях, когда размеры особи не превышают всего 0,02-0,04 мм (рис. 16). У видов с неодинаковой величиной игл последние могут закладываться или одинаковыми по размерам или, чаще, уже с самых ранних стадий развития более крупные иглы выделяются своими размерами. У форм, иглы которых имеют апофизы, вначале закладываются гладкие иглы, на которых по мере роста появляются отростки. При этом в онтогенезе наблюдаются стадии, соответствующие ряду более примитивных форм, описанных прежними авторами как самостоятельные роды. Прекрасный пример этому дает развитие видов рода Lithoptera, последовательно проходящих стадии, подобные родам Quadrilonche, Xiphoptera, Lithopteranna, Lithopterella, Lithopteromma (рис. 16).
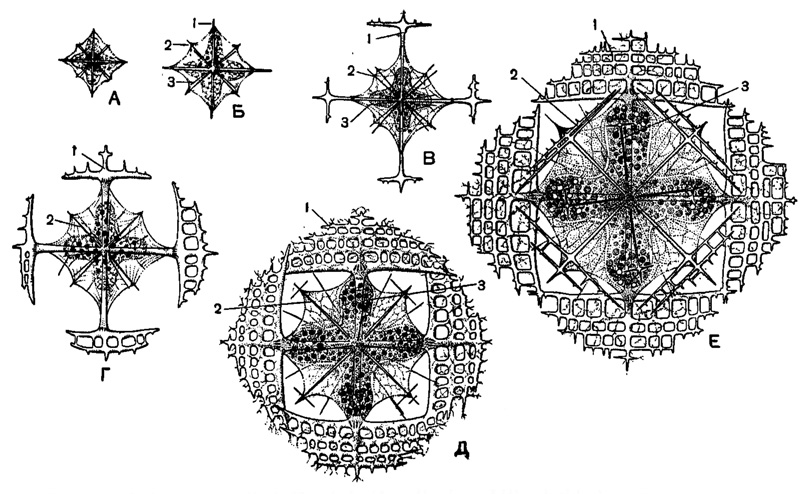
Рис. 16. Lithoptera mülleri Haeckel (Acantharia, Lithopteridae); онтогенез скелета в развивающихся живых зиготах из Средиземного моря: А-Б - очень молодые стадии; В - стадия, соответствующая роду Xiphoptera; Г - стадия Lithopteranna; Д - стадия Lithopterelia; Е - взрослая радиолярия, × 120 (Schewiakoff, 1920); 1, 2, 3 - иглы различных порядков
Точно так же скелет в виде решетчатого шара (в сем. Dorataspidae) закладывается в виде простых игл, на которых постепенно развиваются апофизы, смыкающиеся в конце развития в сплошной панцирь - шар (рис. 17). У Pleuraspis, например, проходятся последовательно стадии Lithophyllum, Phractacantha, Phracaspis.
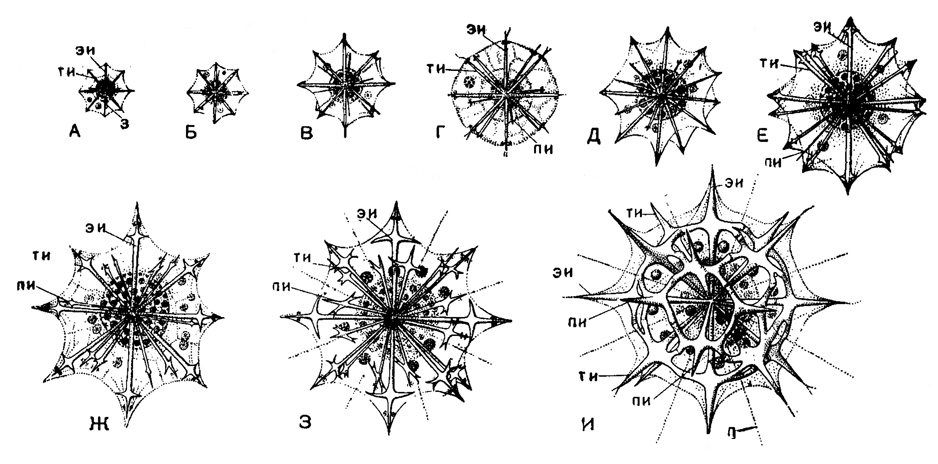
Рис. 17. Pleuraspis costqta J. Müller. (Acantharia, Dorataspidae); онтогенез скелета в развивающихся живых зиготах из Средиземного моря: А-Д очень молодые стадии; Е-Ж - стадия, соответствующая роду Lithophyllum, З - стадия Phractacantha; И - взрослые радиолярии, × 300 (Schewiakoff, 1026): эи - экваториальная игла; ти - тропические иглы; пи - полярные иглы; п - псевдоподии
У форм со вторичными иглами развиваются сначала гладкие пластинки панциря, на которых уже в конце онтогенеза появляются добавочные шипы и иглы.
Одним словом, в онтогенезе акантарий удается проследить формирование сложнейших форм скелета из заложенных первоначально одних и и тех же гладких игл.
Таков же, вероятно, процесс развития шарообразных панцирей и у спумеллярий, судя по фрагментарным данным О. Шрёдера (Schröder, 1909). Сначала закладывается самый внутренний шар, а остальные возникают позже, последовательно от внутренних к внешним. Таким образом, формирование скелетов радиолярий, составленных из концентрических шаров, там, где это изучено у современных форм, идет в центробежном направлении.
Однако у ископаемых форм известны случаи когда морфологически сходные по наружному виду шарообразные скелеты могут либо состоять только из одного шара, либо у них может иметься концентрический второй внутренний шар. Г, Дефландр (Deflandre, 1953) приводит в качестве примера виды спумеллярий из рода Stylosphaera.
Такие случаи дают возможность палеонтологам утверждать, что здесь имеет место развитие скелета, идущее в центрипетальном направлении, когда внутри первичного шара закладывается вторичный. Подобный способ вторичного образования внутренней сферы описывает и А. Шварц (Schwarz, 1931), приводя на своих рисунках с подвергнутых травлению шлифов ископаемых карбоновых спумеллярий сложные картины реконструкции новообразующейся внутренней сферы. Шварц изображает также и стенку центральной капсулы; считая, что она прекрасно сохраняется в ископаемом состоянии, он показывает особые способы впячивания ее при закладке новой сферы и радиальных игл и настаивает на центрипетальном росте скелета. Но данные Шварца требуют подтверждении на современном материале; известные сомнения возникают по поводу возможности сохранения центральной капсулы в ископаемом состоянии, а кроме того, они не согласуются с изложенными выше наблюдениями Шевякова и Шрёдера.
У насселлярий тоже удается установить в ряде случаев рост скелета (Popofsky, 1913): сначала закладываются иглы основного треножника, дополнительные же решетчатые элементы добавляются позже. Так же постепенно наращиваются ярусы брюшного отдела панциря.
У акантарий иглы наращиваются на дистальных концах, по мере роста тела. При этом в ряде случаев наблюдается концентрическое наслаивание материала иглы (Popofsky, 1904).
У феодарий Е. Геккер (Haecker, 1908) описывает закладку зачатка скелетной иглы в виде вытянутой вакуоли с тонкой оболочкой, наполненной студенистым веществом ("Gallertvakuole"). При образовании боковых ответвлений последние почкуются еще на мягкой стадии развития в соответствующих местах, и начинается процесс первичного окремнения, совершающегося в поверхностном слое закладывающейся иглы - возникает полый скелетный элемент. Далее, полость иглы заполняется зернистым содержимым, но так, что остается просвет в центре образующейся иглы, и наступает фаза вторичного окремнения, когда зернистый материал становится гомогенным и превращается в кремнезем. Иначе говоря, наблюдаются четыре фазы формирования игл: закладка студенистой вакуоли, почкование боковых отростков еще в мягком состоянии, первичное и вторичное окремнение.
У представителей рода Aulocleptes студенистая вакуоль может образоваться вокруг некоторых твердых посторонних тел, заглоченных в качестве пищи. Чаще других используются вытянутые панцири диатомовых водорослей (Rhizosolenia). И в данном случае окремнение начинается с поверхности (первичное окремнение, сочетающееся с откладкой зернистого материала вокруг инородного тела и вторичным его окремнением).
Закладка поверхностной скелетной сети (например у Aulosphaera) осуществляется в виде отдельных, самостоятельных первичных игл, вокруг которых выделяются студенистые вакуоли, окремняющиеся в дальнейшем и благодаря тому, что ячейки лежат довольно плотно, приводящие к образованию сплошного панциря. При этом тоже наблюдаются фазы первичного и вторичного окремнения окончательного материала. Может быть, существует даже фаза третичного окремнения. Таким образом, например, панцирь Castanellidae и поверхностная сеть в подотр. Phaeosphaeria возникают онтогенетически принципиально одинаково.
По сути дела, и стенка раковинки Tuscaroridae и Circoporidae закладывается в виде поверхностной сети первичных игл, которые сохраняются в толще стенки и во взрослом состоянии. Вторичное окремнение панциря сопровождается развитием мелкопористой структуры стенки.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'