
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Особенности строения скелета отдельных отрядов
Скелет акантарий астероидного типа с известными вторичными его изменениями у высших представителей отряда.
У всех акантарий имеются прямые, сплошные (не полые) иглы, пронизывающие тело радиолярии и проходящие через его центр или сходящиеся в нем, прободая стенку центральной капсулы. Примитивным акантариям (подотр. Holacantha) свойственны 10 диаметральных игл, сплетенных вместе (но не спаянных) в центре тела. Эти иглы могут быть совершенно ровными и гладкими или посредине каждая игла, там, где она соприкасается с другими, может быть скручена или утончена, с явной тенденцией в этом месте к разрыву. У громадного же большинства (свыше 90% видов) акантарий осуществляется наметившийся у Holacantha разрыв и получается 20 радиальных игл, сходящихся к центру тела радиолярии, соединенных различным образом, так, что все равно получается 10 диаметров (радиальные иглы комбинируются попарно, располагаясь в одну линию по диаметру). Чаще иглы соприкасаются друг с другом своими слегка утолщенными и срезанными пирамидально (конусовидно) проксимальными концами. В иных случаях иглы бывают спаяны друг с другом посредством шарообразного центрального тела (сем. Pseudolithidae) (рис. 21).
Расположены иглы совершенно правильно, подчиняясь известному закону Мюллера (Müller, 1858). Будет ли это 10 диаметральных игл с выступающими из. тела наружу 20 радиальными концами, или это будет 20 радиальных игл, торчащие концы игл располагаются в пять параллельных кругов, по четыре иглы каждый. По аналогии со сферой земного шара, эти круги игл названы: один экваториальный (ему соответствуют четыре экваториальных радиальных иглы), два тропических (им соответствуют восемь тропических радиальных игл, по четыре в каждом полушарии) и два полярных (им соответствуют восемь полярных радиальных игл, по четыре в каждом полушарии (рис. 18 и 19). Эти круги отстоят друг от друга на 30° по окружности (т. е. располагаются по 0°, 30° и 60° по широтам). Экваториальные иглы расположены в одной плоскости под углом 90° друг к другу; условно концы их можно считать выступающими под 0°, 90°, 180° и 270° к окружности экватора. Концы полярных игл выступают строго над и под экваториальными. Если их спроектировать на плоскость экватора, то они лягут на те же градусы. Концы же тропических игл располагаются в промежутках между экваториальными. Проекции их концов на плоскость экватора сместятся на 45° по отношению к концам экваториальных (следовательно, и полярных) игл и займут соответственно положение 45°, 135°, 225° и 315°.
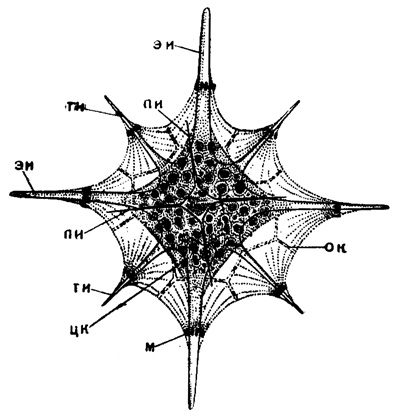
Рис. 18. Acanthostaurus purpurescens Haeckel (Acantharia, Phyllostauridae); современный вид из Средиземного моря. Вид с полюса, × 300 (Schewiakoff, 1926): эи - экваториальные иглы; ти - тропические иглы; пи - полярные иглы; цк - центральная капсула; м - мионемы; ок - отверстия в калимме, через которые выходят наружу аксоподии
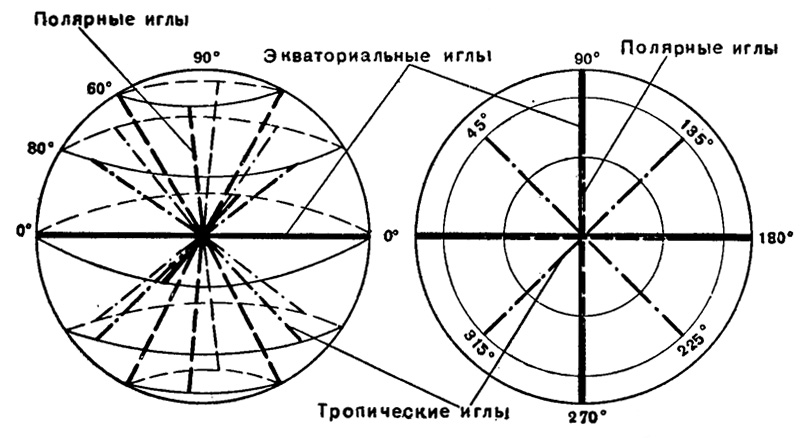
Рис. 19. Схема расположения скелетных игл у акантарии: слева - вид сбоку, справа - вид с полюса
Иного расположения игл у акантарий не бывает (Шевяков, 1926). Упоминаемые в литературе законы Геккеля, Брандта и др. Шевяков сводит к модификации закона Мюллера. Расположение 20 игл по закону Брандта свойственно Rosetta elegans, оказавшейся стадией развития типичной акантарии Hexalaspis heliodiscus; этот закон, следовательно, теряет право на существование в качестве самостоятельной категории. По Шевякову, исключения из закона Мюллера, указанные другими авторами, не относятся к акантариям (сем. Trizonidae с 18 иглами, расположенными в три круга по шести игл каждый, и сем. Actinastridae с 32 иглами). Точно так же сотни радиальных игл у представителей сем. Astrolophidae (роды Actinellius, Astrolophus, сидячая форма Podactinellius) принадлежат таким радиоляриям, которых нет оснований включать в отр. Acantharia.
Иглы акантарий массивные, в примитивных случаях цилиндрические, заостренные дистально. Чаще иглы могут быть сплющены (овальны в сечении), и очень часто вдоль иглы развиваются два или, обычно, четыре ребра, иногда с зазубринами или шипами. Ребра всегда прямые, никогда не идут по спирали (винтом) (рис. 20).
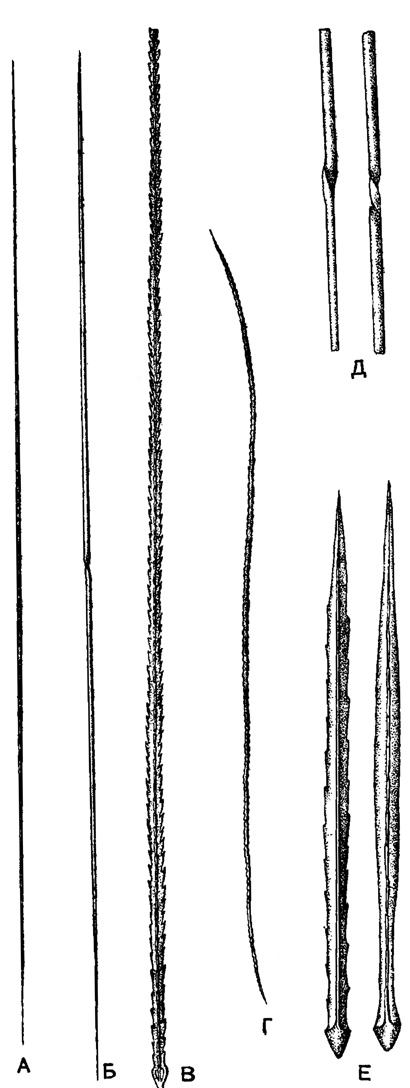
Рис. 20. Скелетные иглы различных акантарий; живые экземпляры из Средиземного моря: А - Acanthoplegma krohnl Haeckel (Acanthdplegmldae) - гладкие диаметральные иглы, пронизывающие все тело, × 100; Б - Acanfhosplra spiralis Haeckel (Acanthoplegmidae) - гладкая диаметральная игла, пронизывающая все тело, перекрученная посередине, × 200; В - Slauracon pallidus Clap. (Stauraconidae) - ребристая зазубренная радиальная игла, × 300; Г - Acanthohlasma serrulala Schewlakoff (Acanthochiasmidae) - зазубренная диаметральная игла, × 200; Д - Acanthosplra spiralis - центральная часть диаметральной Иглы с перекручиванием, × 750; Е - Conacon foliacens Haeckel (Conacopidae) - ребристые радиальные иглы с зубцами на двух противоположных ребрах (Schewiakoff, 1926)
Степень развития игл может быть неодинакова. Чаще у акантарий (у 65% видов) иглы одинаковой величины. В ряде случаев все экваториальные иглы превосходят по своему развитию все остальные роды AcanthoeoUa, Stauracen, A canthostmrus, все представители сем. Lithopteridae. Или же больших размеров достигают только две экваториальные иглы, развитые одинаково (роды Atnphilonche, Amphistaurus, Diploconus) или неодинаково (род Amphilithium). Наконец, известны такие формы акантарий, у которых полярные иглы недоразвиваются и бывают значительно мельче тропических (роды Acanthostaurus, Lonchostaurus, представители сем. Lithopteridae) (рис. 21).
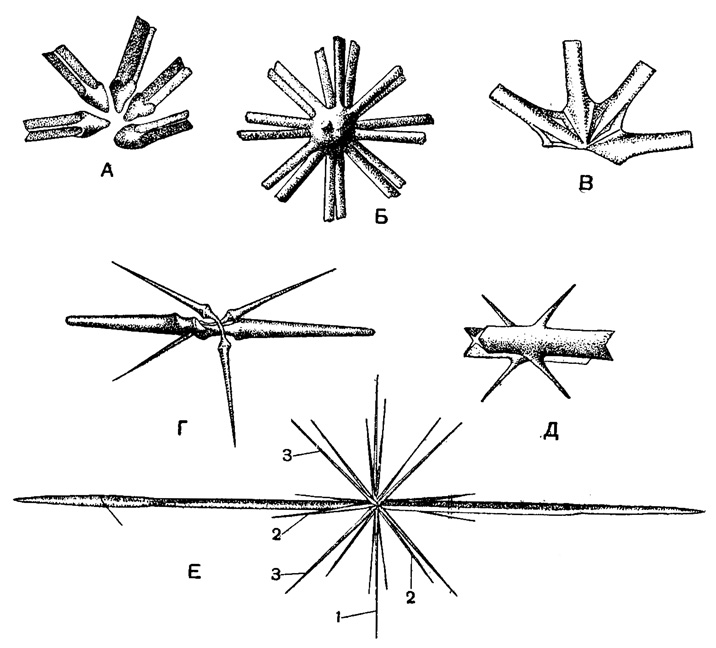
Рис. 21. Скелетные иглы акантарий, живых радиолярий из Средиземного моря: А - Heteracon biformis Popofsky (Gigartaconldae) - сочленение игл внутри тела, × 1550; Б - Heliolithium aureum Schewiakoff (Astrolithidae) - иглы спаяны воедино в центре тела, × 360; В - Acanthometra peliacida J. Müller (Acanthometridae) - иглы сочленены в центре тела помощью крыловидных расширений их проксимальных концов, × 900; Г - Acanthocolla cruciata Haeckel (Acanthoplegmldae) - диаметральные иглы истончены каждая посредине, × 350; Д - Xiphacantha quadrilineata J. Müller (Stauracanthidae) - ребристые радиальные иглы с шипами, × 300; Е - Amphilonche elongata J. Müller (Acanthometridae) - две экваториальные радиальные иглы развиты значительно сильнее всех остальных, × 150 (Schewiakoff, 1926); 1, 2, 3 - иглы различных порядков
Усложнение строения игл происходит, как уже было упомянуто, путем возникновения на них боковых отростков - апофизов (рис. 22). В простейшем случае это 2-4 шиповидных выроста, отходящих более или менее на одном уровне вблизи проксимального конца каждой иглы (Xyphacantha), или этих шипов может быть много (Pristacantha). В сем. Lithopteridae апофизы, наоборот, развиваются у дистального конца каждой иглы; на сильно развитых первичных апофизах, отходящих от игл перпендикулярно, развиваются поперечные перекладины, образующие в конце концов сеть, характерную для всего семейства (рис. 17). Проксимальные апофизы могут также развивать на себе поперечные отростки и образовывать сетевидные пластинки, в результате сближения которых с соседними иглами могут формироваться решетчатые шарообразные панцири, свойственные наиболее высоко организованным акантариям (например сем. Dorataspidae). Можно проследить линию весьма постепенного развития такого панциря-скелета, начиная с рода Haliommatidium, где решетчатые апофизы, изогнутые по сфере, лежат еще изолированно (рис. 23), через род Dictyacantha, у которого апофизы смыкаются краями и частично спаиваются, образуя либо редкую (род Pteuraspis), либо густую сеть (род Lychnaspis).
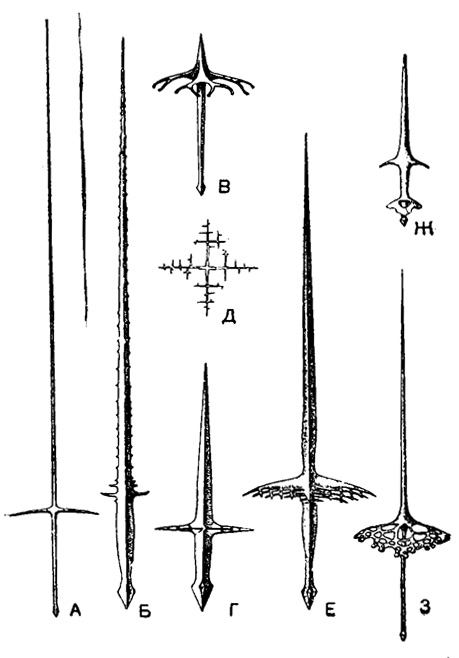
Рис. 22. Развитие апофизов на скелетных иглах акантарий. Живые радиолярии из Средиземного моря: А - Hallommatidium mulleri Haeckel (Pseudolithldae), × 150; Б - Pristacantha multidentata (Stauracanthidae), × 75; В - Stauraspis stauracantha Haeckel (Dorataspidae), × 350; Г - Sfauracanthus orthostaurus Haeckel (Stauracanthldae, × 110; Д - то же, вид сверху; E - Dictyacantha tabulata Haeckel (Dictyacanthldae) - апофизы образуют решетчатый шар, × 150; Ж - Phractopeita dorataspis Haeckel (Phractopeltidae) двухярусные апофизы, × 210; З - Isocaspis elegans Haeckel (Dorataspidae) - апофизы образуют решетчатый шар, × 150 (Schewiakoff, 1926)
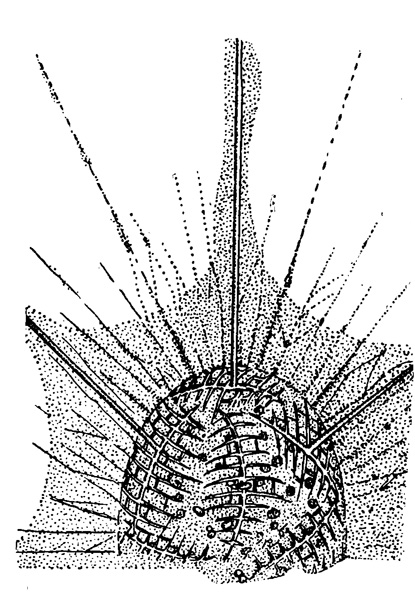
Рис. 23. Hallommatidium mulleri Haeckel (Pseudolithidae); живой экземпляр из Средиземного моря; решетчатые апофизы, не образовавшие еще сплошного шара, × 230 (Schewiakoff, 1926)
Утолщение ветвей апофизов приводит к тому, что панцирь становится сплошным, с небольшими порами в его стенке (род Dorataspis), и в ряде случаев сохраняется шов, указывающий на неполное слияние пластинок возникшего таким образом сплошного панциря (рис. 24). Наконец, изредка у акантарий апофизы возникают одновременно на разных уровнях игл и приводят к образованию двойных шаровидных решетчатых панцирей (род Phradopelta), вложенных один в другой. При этом апофизы на дистальных концах игл, развитые слабо, показывают тенденцию к образованию и третьего концентрического шара.
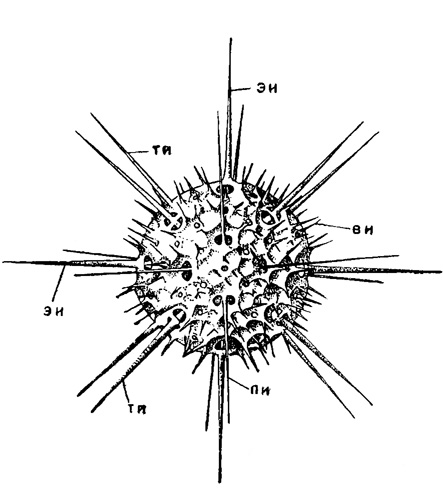
Рис. 24. Histrichaspis dorsata Haeckel (Acantharia, Dorataspidae); из Средиземного моря, вид с полюса, × 180 (Schewiakoff, 1926): эи - экваториальные иглы, ти - тропические иглы, пи - полярные иглы, вт - вторичные иглы
Образование подобного панциря сочетается нередко с развитием вторичного скелета: появляются вторичные (добавочные) радиальные иглы, гладкие или зазубренные, отходящие от пластинок (род Coleaspis). Эти добавочные иглы могут спаиваться вместе, приводя к образованию дополнительной обкладки - футляра вокруг концов радиальных игл, торчащих из панциря. Особенно сильно эта обкладка развивается у форм, у которых сильнее развиты две экваториальные иглы (сем. Diploconidae), что создает двуконическую форму скелета.
Скелет Spumellaria. Низшие, примитивные спумеллярий имеют шарообразную форму. Некоторые из них лишены скелета полностью (ряд родов подотр. Sphaerocollidea). Точно так же бесскелетны и некоторые колониальные формы (Collazoum, Myxosphaera из того же подотряда). Громадное же большинство спумеллярий обладают скелетом, достигающим невероятной сложности строения и неисчислимого разнообразия форм (см. систематическую часть).
В простейшем виде скелет мы видим также у низших спумеллярий в виде отдельных игл, не соединенных и не спаянных друг с другом. Эти иглы могут быть простыми (одноосными), слегка изогнутыми, шиповатыми, трех- и четырехосными одинарными и двойными, когда: две четырехосных иглы спаяны воедино одной общей осью (рис. 25). Концы игл могут быть прямыми, загнутыми, гладкими, шиповатыми, ветвящимися. Эти иглы могут быть рассеяны во всей толще калиммы, могут концентрироваться у ее поверхности в виде скелетного слоя (Thalassoplanda) (рис. 26) или у колониальных форм могут охватывать, как плетенкой, своим скоплением каждую центральную капсулу (Sphaerozoum). Многие колониальные Sphaerocollidea (сем. Collosphaeridae) характеризуются спаянностью игл в подобных скоплениях в маленькие ажурные скелетные сфесы весьма разнообразной и причудливой формы, в ряде случаев с трубчатыми продолжениями краев отверстий (пор); изредка отвороты трубчатых пор становятся столь значительными, что встречаются с таковыми соседних трубочек и, соединившись, приводят к образованию двуслойной сферы. Подобный сетчатый, неправильной формы панцирь развивается и у одиночных представителей сем. Orosphaeridae. Особняком стоят среди Sphaerocolidea одиночные представители сем. Thalassothamnidae с одной крупной иглой, вдоль которой от центра тела расходятся более мелкие иглы (Thalassothamnus, Cytocladus) (рис. 27).
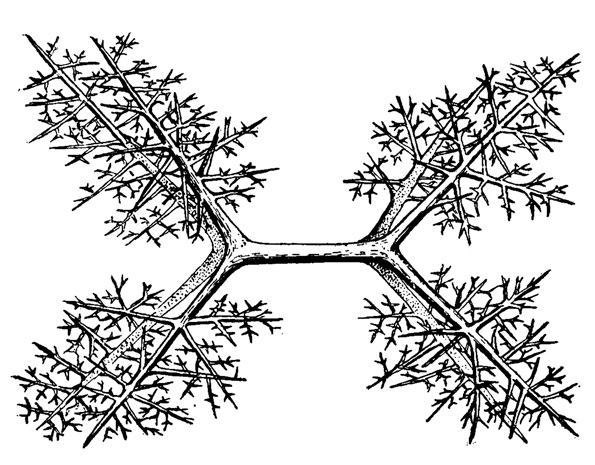
Рис. 25. Sphaerozoum arborescens Haeckel (Spumellaria, Sphaerozoidae); современный вид из южной части Атлантического океана, скелетная игла, спаянная из двух четырехосных, × 200 (Haeckel, 1887)
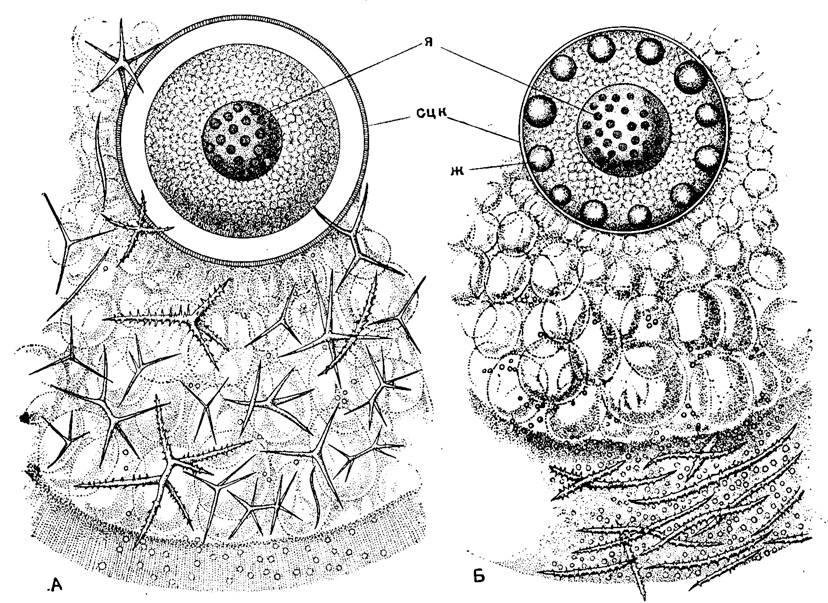
Рис. 26. Распределение скелетных игл в калимме у низших спумеллярий: А - Lamproxanthlum pandora Haeckel (Thalassosphderldae; живой экземпляр из северной части Тихого океана; иглы рассеяны по всей калимме; Б - Thalassoplancta brevispicula Haeckel (Thalassosphaeridae); живой экземпляр из южной части Атлантического океана; иглы сконцентрированы, в скелетном слое калиммы, × 80 (Haeckel, 1887): я - ядро; сцк - стенка центральной капсулы; ж - капли жира
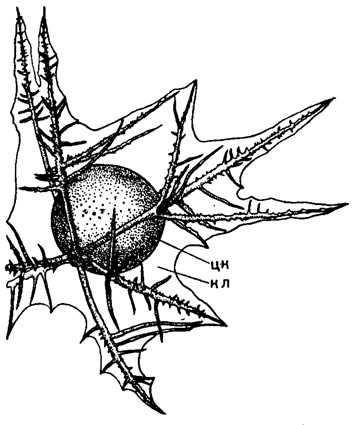
Рис. 27. Thalassothamnus pinetum Popofsky (Spumellaria, Thalassothamnidae); современный вид из вод Антарктики, × 15 (Popofsky, 1908): цк - центральная капсула; кл - калимма
В подотр. Sphaeroidea конструкция скелета наиболее типична для спумеллярий. Именно, в данном подотряде правильная шарообразная форма панциря, с многоосной симметрией, в принципе, сочетается с развитием концентрических скелетных ажурных сфер, вложенных одна в другую. В наиболее сложных случаях число таких сфер может достигать 10 и более (род Arachnosphaera среди Astrosphaeridae). Стенки сфер пронизаны порами, чаще правильной округлой формы, или, реже, сфера принимает вид сети с многоугольными ячеями, или становится губчатой, в виде массы переплетенных беспорядочно тонких перекладин. Правильная сферическая форма панциря у Cubosphaeridae может превратиться в шестигранную (рис. 28). Одна или две внутренних сферы могут закладываться внутри центральной капсулы (так называемые мозговые сферы).
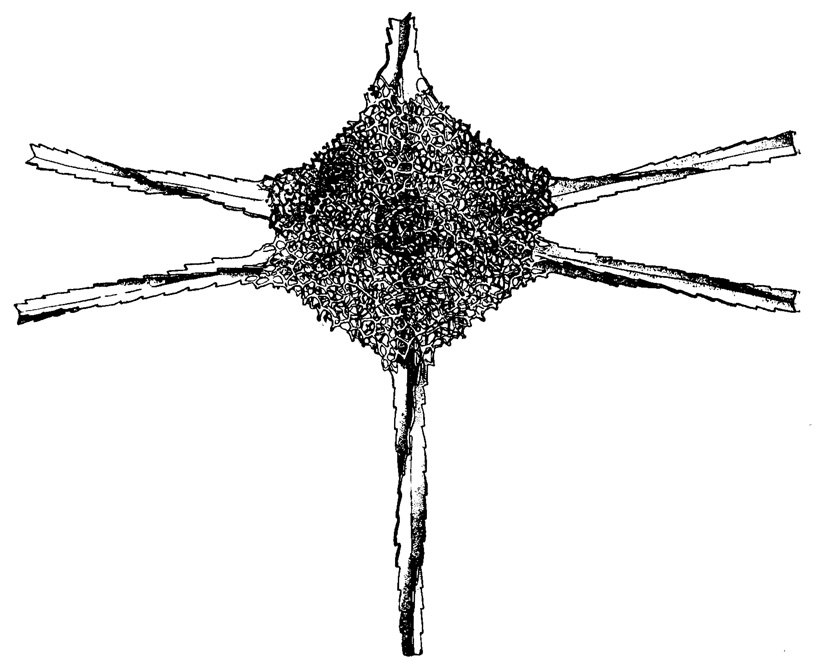
Рис. 28. Hexadoridium streptacanthum Haeckel (Spumellaria, Cubosphaeridae); современный вид из тропической части Тихого океана, × 270 (Haeckel, 1887)
В громадном большинстве случаев сферы панциря соединены друг с другом радиальными иглами, пронизывающими тело радиолярии и прикрепляющимися к стенке внутренней сферы. У ныне живущих Sphaeroidea нет примеров, когда радиальные иглы доходили бы до центра тела, сочленяясь подобно акантариям*. Но среди ископаемых описаны формы (Rüst, 1892), у которых радиальные иглы соединяются воедино внутри сферы панциря. Если нет радиальных игл (сем. Liosphaeridae), скелетные сферы соединены друг с другом посредством вторичных радиальных балок (перекладин), расположенных без особой правильности. Число радиальных игл бывает у Sphaeroidea 2, 4, 6 и больше (до нескольких десятков), чаще расположенных правильно. Относительная длина весьма варьирует. Иглы могут быть простыми, или с разветвлениями на концах, или с боковыми ветвями. В последнем случае тончайшие нитевидные окончания ветвей могут заканчиваться спатиллами (рис. 29) - явление довольно редкое у спумеллярий (в сем. Astrosphaeridae), в противоположность тому, что наблюдается у феодарий, где мы их встречаем часто. Поверхность игл может быть гладкая или с зазубринами; очень часто на игле развиваются до трех продольных гребней (у акантарий обычно четыре), проходящих прямо или по винтовой линии, и игла тогда закручивается спирально, сверлом (чего никогда не бывает у акантарий); у Astrosphaeridae (в наиболее сложно устроенных скелетах) мы видим иглы нескольких категорий. Первичные иглы пронизывают тело и соединяют, следовательно, сферы друг с другом (наиболее обычный случай). Вторичные же иглы отходят от поверхностной сферы и более длинные из них являются главными, а короткие - побочными.
* (Исключение составляют описываемые Поповским (Popofsky, 1912) роды Centrolonche и Centracontium среди Cubosphaeridae, у которых радиальные иглы сходятся в центре тела внутри единственной скелетной сферы; носам же автор замечает, что не исключена возможность в данном случае наличия внутренней сферы в редуцированном состоянии, к которой и прикреплены иглы, тем более что он сам изображает нечто шаровидное в месте соединения игл.)
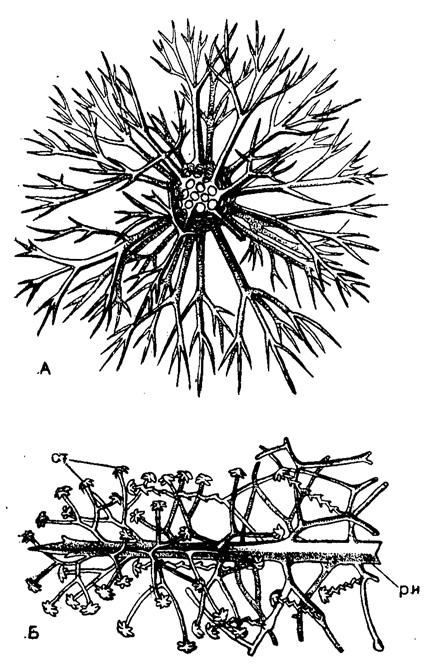
Рис. 29. А - Cladococcus scoparius Haecкel (Spumellaria, Astrosphaeridae); современный вид из тропической части Тихого океана; разветвленные на концах радиальные иглы, × 200. Б - Octodendron spaihillatum Haeckel (Spumellaria Astrosphaeridae); современный вид из тропической части Тихого океана; радиальные иглы с боковыми ответвлениями и спатиллами, × 550 (Haeckel, 1887): ри - ребристая игла; ст - спатиллы
В отр. Prunoidea план строения скелета в общем соответствует тому, что мы видели у Sphaeroidea, т. е. и здесь основу составляют скелетные сферы, вложенные одна в другую. Но, в отличие от Sphaeroidea, панцирь Prunoidea становится одноосным, вытянутым, радиально симметричным. При этом правильный шарообразный характер сохраняют внутренние скелетные элементы, а вытягивается вдоль оси главным образом наружный сетчатый слой. Происхождение этого слоя, по аналогии с Discoidea (см. дальше), весьма вероятно, надо считать вторичным явлением, тем более, что только в подотр. Prunoidea полярные иглы могут заменяться во всех семействах особенными полярными продырявленными трубками.
В сем. Ellipsidae только и имеется этот наружный, вытянутый, одинарный решетчатый панцирь. Он сохраняется и у Artiscidae, но на нем появляется поперечная, экваториальная перетяжка. В сем. Druppulidae панцирь становится многослойным (рис. 30). В сем. Sponguridae поверхностный слой панциря превращается в губчатую массу скелета, столь распространенную в других отрядах. Наконец, у Cyphimdae, Panartidae и Zygartidae появляются поперечные перетяжки на многоосном панцире, приводящие к формированию камер, расположенных метамерно вдоль оси. В случаях максимального расчленения (род Zygartus) сплошными сохраняются только несколько слоев около центральной капсулы, остальные же, в виде шапочек, колпачков, парашютов располагаются во много ярусов на полюсах панциря (рис. 31).
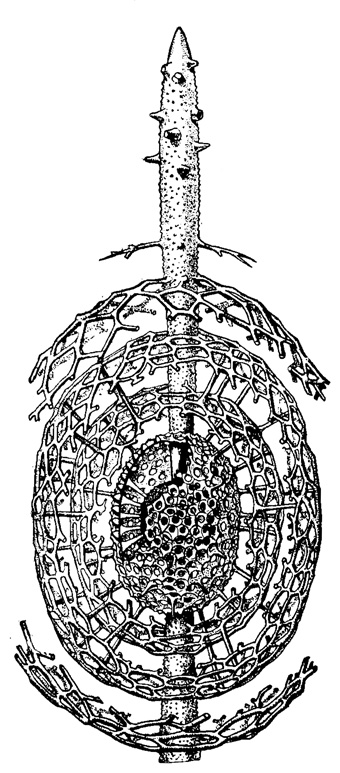
Рис. 30. Cromyatractus ceparius Haeckel (Spumellaria, Druppulidae); современный вид из тропической части Тихого океана; стенки концентрических эллипсоидов вскрыты × 125. (Haeckel, 1887)
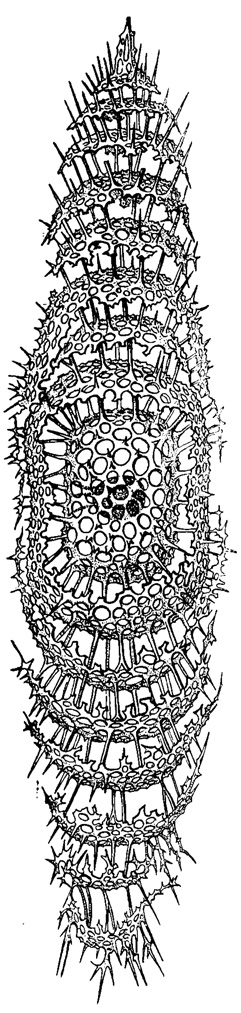
Рис. 31. Zygartus chrysalis Haeckel (Spumellaria, Zygartidae); современный вид из тропической части Тихого океана, × 275 (Haeckel, 1887)
Поверхность панциря у Panartidae может быть гладкой или шиповатой.
Полярные иглы чаще всего бывают одинаковыми по размерам, однако есть роды с недоразвитой одной иглой (Druppotractus, Xiphatractus) или одна игла исчезает совсем (Lithapium, Xiphostylus). В редких случаях или обе полярные иглы или одна из них может заменяться, пучком игл-шипов (Lithomespilus, Cyphirtus). Иглы бывают гладкими, трехгранными, но без. винтового скручивания.
Соединение внутренних сфер друг с другом и с наружным слоем осуществляется радиальными иглами, когда они есть, и радиальными; балками (перекладинами), расположенными довольно беспорядочно в экваториальной области панциря.
Особого устройства скелет наблюдается в подотр. Discoidea, сильно сплющенный по главной оси до степени плоской лепешки или диска, нередко со вздутием в центре. В сем. Cenodiscidae, Phacodiscidae и Coccodiscidae поверхность диска с обеих сторон покрыта особым продырявленным слоем вторичного скелета - факоидом (рис. 115), принимающим иногда губчатую структуру. По ребру факоид может быть оторочен скелетной пластинкой - экваториальным кругом, гладким, фестончатым, вырезанным или шиповатым и игольчатым, составленным чаще всего из прямоугольных ячей, расположенных однослойно. Но в сем. Coccodiscidae экваториальный слой состоит из игл, расположенных преимущественно многослойно; поры факоида округлые.
Внутри факоида скелет представлен обычными для спумеллярий одной или несколькими ажурными сферами, соединенными друг с другом и с наружным факоидом многочисленными балками, перекладинами. В сем. Porodiscidae факоид заменяется двумя поверхностными продырявленными или губчатыми пластинками, а экваториальный, круг распространяется до центра панциря с совершенно плоской, вроде монеты, поверхностью. В центре диска у Porodiscidae остается маленькая округлая камера скелета, ячейки же экваториального круга ложатся либо концентрически, либо завиваются в плоскую спираль (рис. 117-122).
Неравномерный рост панциря приводит у Porodiscidae к развитию звездообразно расположенных двух, трех, четырех, пяти или шести сетчатых рук, одинаковых или с преобладанием в размерах одной из них, нарушающей правильную радиальность. Между руками может протягиваться сетчатая скелетная перепонка - патагий (patagium, рис. 5), а их поверхность может покрываться губчатой массой скелета.
У Pylodiscidae одиночная центральная сфера окружена двумя слоями ажурного диска, с тремя большими, правильно по радиусам расположенными отверстиями, нередко подразделенными экваториальным кольцом.
Наконец, у Spongodiscidae в общем породисцидный панцирь покрыт с обеих сторон губчатой массой скелета (рис. 143).
Большой сложности достигает скелет в подотр. Larcoidea, теряющий радиальную симметричность. В схеме Здесь возникает эллипсоидная форма с тремя осями неодинакового размера. Самая длинная из осей - вертикальная, средняя по длине - боковая и самая короткая ось - передне-задняя. В исходном состоянии панцирь Larcoidea слагается из трех неодинаковых сплющенных (эллипсовидных) лентообразных скелетных колец; они пересекают друг друга в трех взаимно перпендикулярных плоскостях (рис. 32). Наибольшее кольцо является по положению фронтальным (боковым), среднее по размерам - экваториальным, а самое малое - сагиттальным. Эти ленты-кольца поддерживают простую, шаровидную или такую же трехзональную (ларкоидную) внутреннюю камеру. Кольца заменяют радиальные балки, свойственные прочим спумелляриям (при наличии- внутренних и внешних элементов скелета). Описанная ларкоидная камера далее входит в состав- всех видоизменений и усложнений скелета Larcoidea.
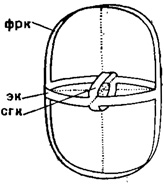
Рис. 32. Схема строения скелета Larcoidea (Spumellaria) (Schröder): фрк - фронтальное кольцо; эк - экваториальное кольцо; сек - сагиттальное кольцо
Так, например, она может быть окружена скелетными поясами у Pylonidae - получается панцирь с большими парными отверстиями-окнами. Или вокруг ларкоидной камеры правильным порядком располагаются шаровидно вздутые камеры с решетчатыми стенками (сем. Tholonidae, Zonaridae). Ларкоидная камера может окружаться неправильной формы стенками (сем. Phorticidae) или неправильно нагроможденными камерами (сем. Soreumidae). Очень, своеобразная наружная решетка в сем. Lithelidae может завернуться инволютно вокруг ларкоидной камеры в плоскую спираль. Или же спиралью располагаются скелетные камеры, образуя завитую башенку (турбоспираль) (рис. 33). Наконец, в сем. Larnacidae ленты-кольца исчезают, панцирь приобретает очень простую форму сплошного решетчатого слоя, даже иногда без внутренней камеры или с нею, шаровидной формы.
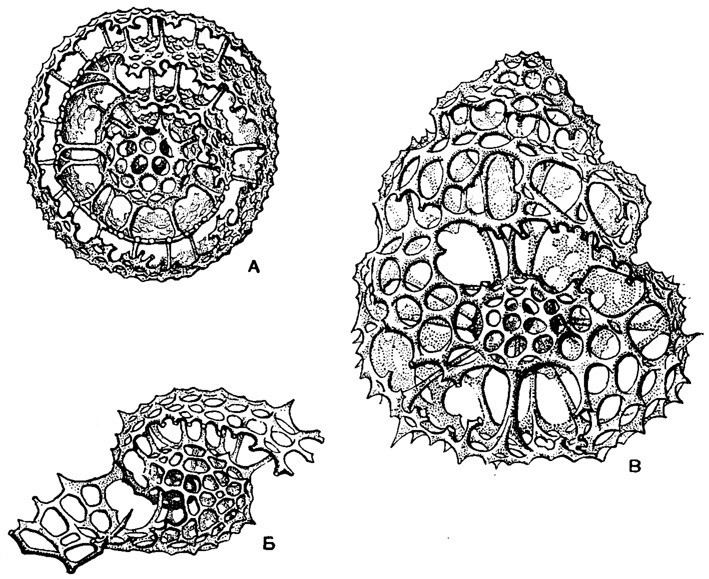
Рис. 33. Современные виды сем. Lithelidae (Spumellaria) из центральной части Тихого океана, × 300: А - Spirema melonia Haeckel (Spumellaria, Lithelidae); Б - Lithelius Solaris Haeckel; изображена только центральная часть панциря; В - Streblopyle helicina Haeckel (Haeckel, 1887)
Панцирь Larcoidea может быть с иглами, торчащими во все стороны, или без них, гладким. Сама стенка панциря, все его элементы в громадном большинстве решетчатые, но в редких случаях бывают и губчатые, подобно другим Spumellaria.
Скелет Nassellaria. Бесскелетное состояние у насселлярий (в подотр. Nassoidea) сочетается с резкой редукцией внекапсулярной плазмы (калиммы). Приходится признать, что одновременно произошло полное исчезновение скелета. Его роль приняла на себя плотная стенка центральной капсулы, занявшая периферическое положение и одетая тонким слоем внекапсулярной калиммы. Вторичный характер отсутствия скелета у Nassoidea главным образом подчеркивается фактом чрезвычайно большого сходства или тождества строения игл у некоторых низших насселлярий (из подотр. Plectellaria) с тем, что мы можем видеть у низших спумеллярий (из подотр. Sphaerocollidea). Последнее обстоятельство позволяет филогенетически вывести насселлярий из спумеллярий (см. систематику Nassellaria). Бесскелетные формы приходится поставить в стороне от этого пути и рассматривать их как вторично упростившиеся.
Скелет насселлярий в целом сравнительно морфологически весьма однороден, являясь производным единственной основного типа иглы (спикулы), в виде треножника с отходящим кверху лучом. Отсюда возникает и общая одноосность раковинки панциря насселлярий, наличие сужения на одном конце раковинки, и наоборот, развитие раструба, широко раскрытого у устья на другом - развивается, как уже было указано выше, форма парашюта, среди радиолярий представленная только в этом отряде (рис. 15). Правильные шарообразные раковинки не свойственны насселляриям и возникают крайне редко всем. Stephoidae в результате комбинирования вторично развивающихся колец скелета; при этом не бывает радиальных игл и никогда не встречается вложения элементов скелета один в другой, что столь распространено у спумеллярий.
Единый в общем план строения скелета насселлярий позволяет провести гомологию отдельных частей скелета и одинаковым элементам присвоить одни и те же обозначения, принятые и на наших рисунках.
Исходный тип скелета - спикула в виде треножника - типичен для представителей сем. Plectoidae подотр. Plectellaria (рис. 34). В наиболее примитивном случае эта игла слагается из четырех лучей, три из которых образуют треножник, а четвертый торчит кверху (типа триэны с углами между лучами в 120° - Plagoniscus). Обычно лучи зазубрены; они могут давать отростки в стороны; каждый луч может заканчиваться пучком разветвлений, и все это может анастомозировать друг с другом, создавая сеть, пронизывающую калимму тела радиолярии (например Polyplecta, Plectanium). Видоизменения этого типа иглы происходят путем вторичного увеличения числа лучей (вторичные лучи легко узнать по их меньшим размерам - Polyplagia), путем образования двойных игл, возникших спаиванием торчащих кверху лучей (Рlаgonium). Очень характерное видоизменение наблюдается часто в виде включения в состав иглы балки, разъединяющей два центра схождения лучей (Periplecta). И тут наблюдается вторичное разветвление лучей, и между ними могут развиваться дуговидные соединения (Plectantha, Phormacantha).
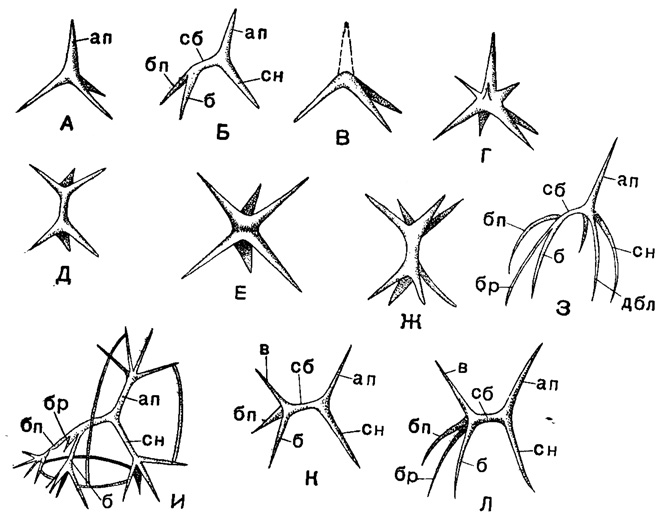
Рис. 34. Схематическое изображение игл скелета насселлярий (Plectellaria): А - тип рода Plagoniscus; Б - Periplecta; В - Trlplagia; Г - Potyplagia; Д - Plagonium; Е - Hexaplagia; Ж - Polyplagia; З - Campylacantha; И - Phormacantha; К - Plagiocarpa; Л - Verticillata; отростки: ап - апикальный; сн - спинной, б - боковой, бп - боковой правый, сб - срединная балка, в - вертикальный, дбл - добавочный боковой левый, бр - брюшной
В сем. Stephoidae из того же подотряда развиваются скелетные кольца в сочетании с лучами треножника или же при полной их редукции. Формирование колец происходит путем спаивания растущих навстречу отростков на лучах - получаются массивные дуги, переходящие прямо в основания лучей и вместе с последними образующие кольцо(Coriniscus, Zygocircus) (рис. 35). Наиболее примитивны виды, у которых развивается только одно сагиттальное кольцо. При дальнейшей специализации к этому кольцу может добавиться горизонтальное базальное кольцо (Semantis), в ряде случаев разделенное перегородками на вторичные мелкие кольца (Semantidium). Второе кольцо может занять и фронтальное положение, став под прямым углом к сагиттальному (Zygostephanus, рис. 36 и 37). Оба эти кольца могут быть опоясаны экваториальным третьим (Trissocircus). В результате может получиться правильный шар - редкий случай у насселлярий (Trissocyclus) (рис. 14, Б). При редукции некоторых дуг возникает довольно большое разнообразие кольчатых скелетов. Наконец, в подсем. Tympaninae два горизонтальных кольца соединяются различного рода и числа вертикальными перемычками; от этих колец могут сохраниться только их части, дуги, и получаются скелеты в виде многогранников типа Prismatium, Lithocubus и др. (рис. 37, Б). Гладкими все названные перекладины бывают редко; чаще на них развиваются шипы, боковые отростки подчас сложного, причудливого ветвления.
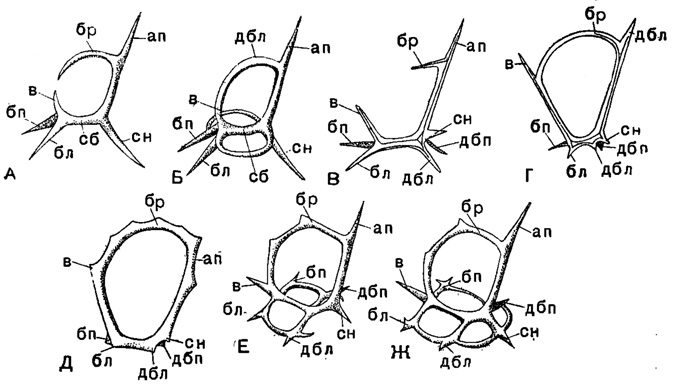
Рис. 35. Схематическое изображение игл скелета насселлярий (Plectellariа): А - тип рода Cortina; Б - Cotriniscus; В - Plagiocarpa; Г - Archicircus; Д - Zygocircus; Е - Semantis; Ж - Semantrum (Popofsky, 1913); отростки: ап - апикальный, сн - спинной, бл - боковой левый, бп - боковой правый; сб - срединная балка, в - вертикальный, дбл - добавочный боковой левый, дбп - добавочный боковой правый, бр - брюшной
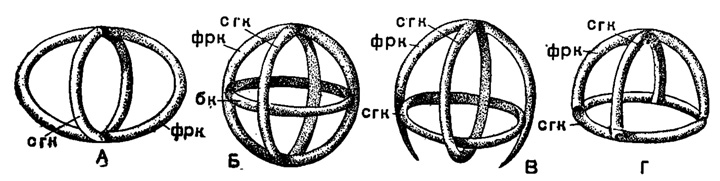
Рис. 36. Схематическое изображение скелета насселлярий сем. Stephoidae: А - тип рода Zygostephanusi Б - Тrissocircus; В - Podocoronis; Г - Coronidium (Popofsky, 1913) кольца: сгк - сагиттальное, фрк - фронтальное, бк - базальное
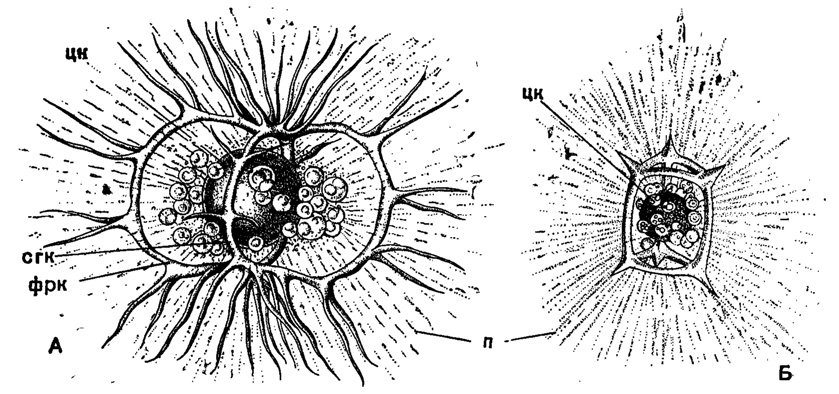
Рис. 37. А - Zygostephanus mulleri Haeckel (Nassellaria, Stephoidae); живой экземпляр из Средиземного моря, × 350; Б - Prismatium tripleurum Haeckel (Nassellaria, Stephoidae); живой экземпляр из Средиземного моря, × 350 (Haeckel, 1862): цк - центральная капсула, окруженная зооксантеллами; сгк - сагиттальное кольцо; фрк - фронтальное кольцо; п - псевдоподии
Настоящая решетчатая раковинка формируется в последнем подотр. Cyrtellaria и достигает в нем необычайного разнообразия, сохраняя, однако, в основе своего построения тот же скелетный треножник, о котором неоднократно говорилось выше. Решетчатую стенку раковинки можно представить возникшей из боковых ответвлений лучей. При этом в сем. Cyrtidae возможны два типа формирования скелета (рис. 38). В случае, если боковые ответвления лучей треножника формируют стенку раковинки со включением в ее состав самих лучей, раковинка становится граненой (трехгранной при исходном числе лучей три - Dictyophimus, и многогранной, когда число лучей увеличивается - Cinclopyramis, Plectopyramis). Если же лучи треножника остаются свободными, то раковинка приобретает цилиндрическую форму, а сами лучи могут либо на всем протяжении, либо только у основания соединяться со стенкой раковинки сетчатым ребром (Dictyocerys, Dictyocodon, Pterocanium, Artapilum). Число лучей очень часто равно исходному - трем, но может увеличиваться до девяти (довольно частый случай) и более. Редукция лучей треножника чаще сопровождается исчезновением нижних лучей, верхний же в виде той или иной степени развития шипа в большинстве случаев сохраняется (около него нередко развиваются дополнительные шипы).
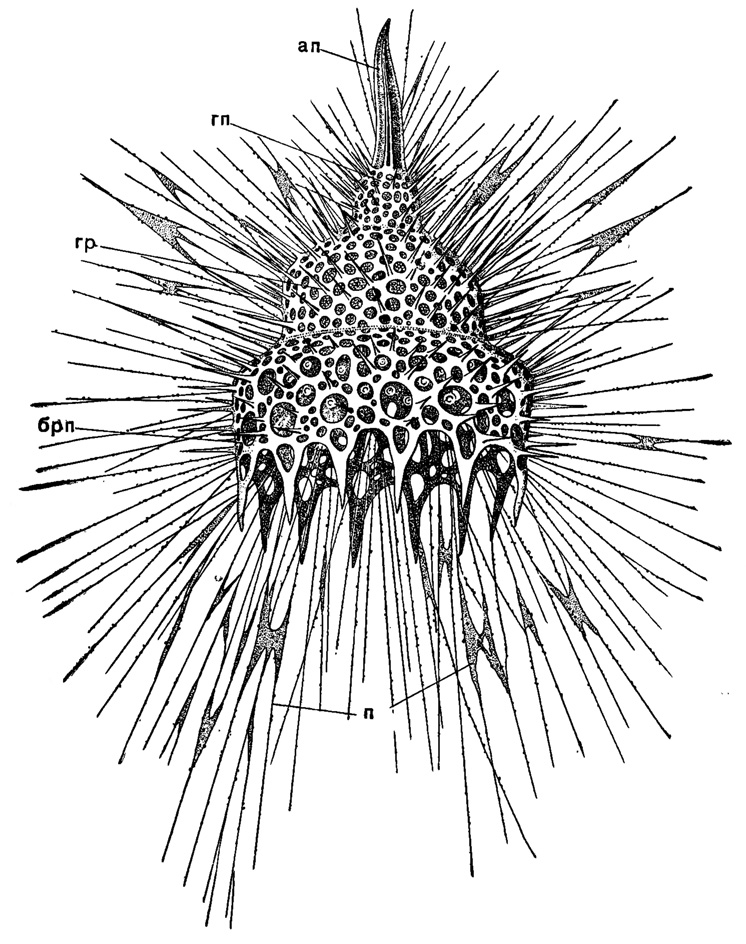
Рис. 38. Oalocyclas monumenium Haeckel (Nassellaria, Cyrtoidae); живой экземпляр из тропической части Тихого океана, × 300 (Haeckel, 1887): ап - апикальный шип; гп - головной отдел панциря; гр - грудной отдел; брп - брюшной отдел; п - псевдоподии
Край устья бывает либо гладким (Cyrtocalpis), либо зазубренным; зубцы могут достигать больших размеров (Calocyclas, Alacorys, Anthocyrtidium). Устье раковинки чаще широко зияет, но у многих форм замыкается решетчатой пластинкой-диафрагмой или резко суживается, превращаясь в решетчатую трубку, открытую (Eusyringium) или замкнутую (Theophaena).
Поперечные пережимы на раковинке Cyrtidae приводят к образованию многокамерности (рис. 164-170). В первую очередь,выше основания лучей обособляется цефалум (или "головной" отдел), часто шарообразной формы. Ниже его,в случае двухкамерной раковинки, будет торакс (или "грудной" отдел скелета). Между названными отделами внутри либо имеется прямое сообщение, либо на границе между ними залегает продырявленная диафрагма. Дополнительная перетяжка на грудном отделе приводит к обособлению так называемого абдомена ("брюшного отдела"). Последний, в свою очередь, может дать начало ряду камер постабломена. В громадном большинстве случаев камеры при таком варианте следуют одна за другой по прямой линии, изредка же раковинка изгибается в виде рога (Lithostrobus). Наконец, исключительно редко многокамерная раковинка у некоторых видов скручивается спиралью (Siphocampe spiralis).
Раковинка в сем. Botryoidae в общем напоминает таковую у Cyrtoidae, но отличается подразделением головного отдела продольными перетяжками на камеры, иногда беспорядочно расположенными. Кроме того, от головного отдела могут отходить решетчатые трубки, представляющие, по мнению Поповского, видоизмененные лучи все того же треножника.
В сем. Spyroidae скелет весьма разнообразный теряет радиальную симметрию и становится двустороннесимметричным. В основе этого скелета лежит тот же треножник, поверх которого развивается решетчатый цефалум. В его стенках закладывается скелетное кольцо, расположенное сагиттально. Там, где проходит кольцо, на раковинке получается вдавление, разделяющее головной отдел на две полусферических половины. Число лучей нередко низводится до двух, и они отходят тогда от "головного" отдела фронтально, подчеркивая этим двустороннесимметричный характер слета (Dorcadospyris). Концы лучей могут, в свою очередь, спаяться в сплошное кольцо (Gamospyris, Stephanospyris). Апикальный шип часто бывает развит очень сильно, и рядом с ним могут быть добавочные шипы (Triceraspyris). Цефалум может отделиться перетяжкой от торакса и даже от абдомена (Lamrospyris). Все отростки раковинки Spyroidae редко бывают гладкими, обычно они вооружены шипами и боковыми отростками.
Скелет Phaeodaria. Среди феодарий встречаются, правда, очень редко, бес. скелетные формы. Это состояние может быть первичным признаком примитивности (сем. Caementellidae в целом), или скелет исчезает вторично, как это мы имеем у некоторых Medusettidae (Miracella, Lobocella и др.). При этом мягкое тело радиолярии может быть защищено посторонними частицами, агломерированными на поверхности тела из обломков мелких радиолярий, диатомовых водорослей, силикофлягеллят и др). Конвергентно данная особенность проявляется и среди Caementellidae (род Caementella) и у Medusettidae (род Miracella).
Простейший тип скелета у феодарий слагается из отдельных полых, трубчатых игл, не связанных друг с другом, лежащих в теле и придающих ему многоосный характер (подотр. Phaeocystina). Иглы эти бывают двух категорий - тангентальные и радиальные. Тангентальные иглы обычно мелкие, слегка изогнутые, гладкие, чаще шиповатые и лежат слоем вблизи поверхности тела в калимме. Только тангентальные иглы встречаются редко - в сем. Cannorhaphidae с тремя родами. В других случаях, они сочетаются с радиальными иглами, пронизывающими толщу тела в отличие от Acantharia (не проникая, однако, своими проксимальными концами в центральную капсулу) и, как правило, лежащими довольно беспорядочно, не связанными друг с другом. Исключением является весьма небольшое семейство, всего с одним родом - Astracanthidae, где имеются только радиальные иглы, сходящиеся к центру тела и сочленяющиеся вместе (рис. 39). В редких случаях радиальные иглы гладкие; чаще они зазубрены или снабжены различного рода отростками - боковыми и концевыми. Формирование игл у Aulocleptes (сем. Aulacanthidae) связано со своеобразным процессом включения в состав иглы заглоченных игольчатых панцирей диатомей (Rhizosolenia) или даже игл других Aulacanthidae. Число радиальных игл может доходить до 200-300; у Aulacantha, чаще колеблется от 20 до 40.
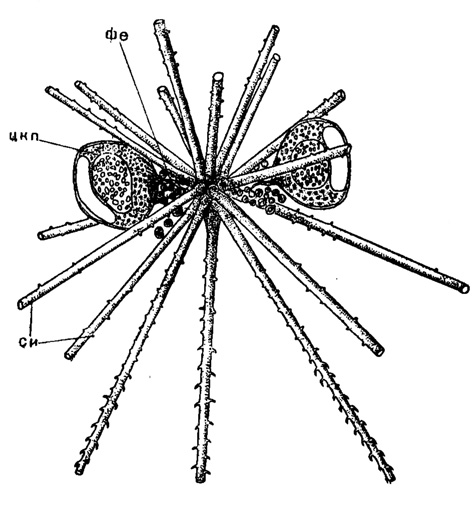
Рис. 39. Astracantha paradoxa Haecker (Phaeodaria, Astracanthidae); живой экземпляр из Индийского океана, × 50 (Haecker, 1908): фе - феодий; цкп - центральная капсула; си - скелетные иглы
Такой тип скелета свойственен только низшим феодариям. У прочих представителей этого отряда скелетные элементы соединяются в разнообразные комплексы. В подотр. Phaeocalpia тангентальные иглы спаиваются вместе, образуя сплошной панцирь, часто шаровидный, но одноосный, радиально-симметричный благодаря появлению большого отверстия - устья (пилома), ведущего внутрь. В сем. Castanellidae шарообразный панцирь имеет толстые стенки и пронизан многочисленными крупными порами. Полые радиальные иглы отходят от поверхности панциря наружу (внутрь тела они никогда не вдаются) и могут быть двух категорий - главные и побочные, обычно гладкие и неразветвленные. Устье часто бывает вооружено зубцами. У Circoporidae поры концентрируются около оснований игл; последние также полые и могут ветвиться на концах, панцирь же становится многогранным, с поверхностной скульптурой. У Polypyramidae панцирь ажурный, пронизан порами разной величины; ветвящиеся радиальные иглы основанием покоятся на пирамидальных подпорках. У Tuscaroridae и Porospathidae устье открывается на конце суженного, трубчатого перистома. Толстая стена панциря Tuscaroridae сплошная и пронизана мельчайшими порами, а у Porospathidae между сосочками поверхностной скульптуры имеются глубокие ямки (или тоже поры?). Полые неветвящиеся радиальные иглы у Tuscaroridae отходят как от краев устья, так и от поверхности панциря, а у Porospathidae только от стенки панциря. Тончайшие тангентальные иголочки всегда видны в толще стенки панциря у представителей подотр. Phaeocalpia.
В подотр. Phaeosphaeria тангентальные иглы преобразуются в поверхностную сеть из крупных ячеек. Перекладины также полые, и в местах соединения, в узловых точках они могут или сочленяться (сем. Aulosphaeridae) или спаиваться полностью (сем. Sagosphaeridae). В некоторых случаях сеть может быть двухслойной. Радиальные иглы отходят наружу от узловых точек сети и могут основанием начинаться от вершин пирамид, образованных перекладинами сети. Подобная сеть со слитыми в узловых точках перекладинами развивается и у колониальных Tuscaroridae (в роде Tuscaretta) - их радиальные иглы с шипами плотно укрепляются в ячеях этой сети. Наконец, в подотр. Phaeosphaeria, в сем. Cannospharidae, наряду с поверхностной сетью типа Aulosphaeridae, вокруг центральной капсулы развивается небольшой внутренний скелетный шар с устьем (пиломом), соединенный с поверхностной сетью многочисленными тонкими радиальными перекладинами.
В подотр. Phaeogromia сохраняется тип скелета, свойственный Tuscaroridae, т. е. имеется панцирь со сплошной стенкой, с мельчайшими порами, сложно устроенными, ведущими в особые микрополости в толще стенки (диатомовая структура) и с устьем на конце перистома. Панцирь уплощенный и становится двустороннесимметричным. В сем. Challengeridae радиальные иглы полые, короткие, отходят от ребра раковинки в определенных местах, аборально; край перистома у них может иметь зубцы и отростки, а сам перистом в виде глотки может вдаваться в просвет раковинки. В сем. Medusettidae радиальные иглы прикрепляются только по краю перистома; они длинные, и полость в них разделена близко поставленными перегородками. Стенка панциря ячеистая, альвеолярной структуры. У ряда Medusettidae разрастающаяся центральная капсула вытесняет панцирь; его остатки в виде первичного скелета сохраняются только в области перистома и пилома (Planetonetta, Atlanticella), а на смену исчезнувшему панцирю развивается вторичный скелет вокруг оттесненного в сторону феодия, и большой плавательный пузырь.
Наконец, особняком стоят двустворчатые скелеты в подотр. Phaeoconchia и Phaeodendria (каждый с одним семейством). В сем. Concharidae (Phaeoconchia) имеется занимающая поверхностное положение двустворчатая раковинка, сферическая или сплющенная, створки которой плотно приложены или гладкими краями (редко), или обычно имеется замок из ряда зубцов, заходящих друг за друга соседних створок и обусловливающих прочное их соединение. Стенки створок пронизаны крупными порами. Точно такую же двустворчатую пористую раковинку, но только очень маленькую, имеют вокруг центральной капсулы представители сем. Coelodendridae (Phaeodendria). К обеим сторонам прилегают особые шлемовидные покрышки, от которых в обе стороны от центральной капсулы отходит по системе радиальных игл, сложно разветвленных, часто дихотомически, и концевыми разветвлениями могущих образовать поверхностную скелетную сеть. Радиальные иглы могут быть двух сортов: короткие - дендриты и длинные - контурные иглы. Среди последних обычно выделяются длиной две, вокруг которых распределяется калимма, и все тело подразделяется тогда на две большие лопасти.
У многих представителей феодарий мы наблюдаем особого типа концевые разветвления радиальных игл - отмеченные уже у спумеллярий спатиллы. Это могут быть короткие веточки, булавовидно вздутые, неправильно расчлененные, или они могут быть представлены чечевицеобразными бляшками с зазубренным краем (сем. Aulacanthidae, Aulosphaeridae, Sagosphaeridae). Или же это тонкие нитевидные ответвления, собранные пучками и несущие на концах якоревидные зацепки из нескольких направленных назад и расположенных звездообразно зубчиков. Подобного сорта спатиллы в виде исключения отходят от булавовидно вздутых концов игл в роде Aulocoryne; они имеются у Cannosphaeridae, но особенно распространены у Medusettidae и Coelodendridae с их сильно развитыми радиальными иглами. С помощью спатилл на иглах особенно прочно укрепляется студенистая калимма, и скелетная функция игл получает наибольшее выражение.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'